











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La infección plasmodial placentaria submicroscópica (IPPS) por Plasmodium falciparum es frecuente en los países endé micos y contribuye al desarrollo de anemia materna y bajo peso al nacer 1. La IPPS por P. vivax (IPPS-vivax) es casi desconocida en el mundo.
La técnica estándar para el diagnóstico de malaria en las zonas endémicas es la gota gruesa, que, por definición, no detecta las infecciones submicroscópicas, estas son usualmente asintomáticas. Estas dos condiciones, submicroscópicas y asintomáticas, ha mantenido a la IPPS en el olvido absoluto hasta comienzos del siglo xxi. Pero ya hay voces con fuertes argumentos que dicen, por ejemplo, que las infecciones asintomáticas tienen consecuencias importantes en la salud y la sociedad, y deben renombrarse como infecciones de malaria crónicas 2.
La inmunopatogénesis de la malaria placentaria (MP) debe considerar que el embarazo es un estado fisiológico único en el que el sistema inmunológico materno debe proteger a la madre contra infecciones y otras noxas, y, a la vez, modular su respuesta inmune para evitar el rechazo del feto semialogénico 3. La presencia de Plasmodium y sus productos (como hemozoína) en el tejido placentario, alteran en esencia el ambiente inmune que regula la placenta 4.
En los casos de malaria gestacional y placentaria, el incremento de citoquinas TNF-α, IFN-γ e IL-10 se asocia con daño trofoblástico, bajo peso al nacer y prematurez 5. La expresión de citoquinas proinflamatorias placentarias es alta en mujeres con MP y la expresión de citoquinas antiinflamatorias es baja 5 - 9.
La MP altera la remodelación angiogénica 10 , 11. Los infiltrados de células mononucleares y sus productos inflamatorios se asocian con bajo peso al nacer y es más breve y menos inflamatoria en mujeres multigrávidas que en primigrávidas. Estas diferencias pueden explicar los diferentes resultados que estas mujeres y su descendencia padecen 12 - 14.
En el embarazo normal, son evidentes los procesos fisiológicos de apoptosis, inflamación, hipoxia, vasculogénesis y angiogénesis, entre otros, que están estrictamente controlados y en los que es decisivo el papel de señalizadores/mediadores de la comunicación celular (CC) 15 - 19. El sistema de CC es un complejo entramado entre células, las cuales envían señales a otras, estas últimas con funciones receptoras, produciéndose una respuesta fisiológica y un cambio en la actividad celular. En la CC, los señalizadores de procesos fisiológicos o fisiopatológicos son muchos y tienen diverso origen (hormonas, factores de crecimiento, citoquinas, prostaglandinas, leucotrienos, etc.) 20.
El objetivo del estudio fue determinar la asociación de la infección submicroscópica por P. vivax o P. falciparum con eventos histopatológicos, el comportamiento de células inmunitarias, expresión de genes asociados a citoquinas, hipoxia, angiogénesis e inflamación en placentas.
MENSAJES CLAVE
Motivación para realizar el estudio: Se tiene limitada evidencia sobre los efectos histopatológicos e inflamatorios de la infección placentaria submicroscópica por P. falciparum y P. vivax.
Principales hallazgos: Las placentas pertenecientes a mujeres residentes en la mayor zona endémica de Colombia, en el noroccidente del país, estaban afectadas por infección plasmodial submicroscópica (IPS), no detectada con gota gruesa, pero si con reacción en cadena de la polimerasa cuantitativa (qPCR). En la placenta, esta IPS causa daños tisulares y afecta mediadores de procesos como inflamación, hipoxia, angiogénesis, entre otros, comparada con placentas no infectadas. Tanto P. vivax como P. falciparum actúan como patógenos.
Implicancias: Hay que mejorar mucho las acciones de diagnóstico y tratamiento de la infección plasmodial gestacional en la consulta prenatal, que debe incluir obligatoriamente la práctica de gota gruesa en cada control.
MATERIALES Y MÉTODOS
Diseño y población de estudio
Se realizó un estudio transversal donde se compararon tres grupos independientes. El tamaño de la muestra se definió por conveniencia, según las muestras placentarias existen tes en el banco de tejidos de nuestro grupo de investigación. Se seleccionaron 20 muestras con MP por vivax (MP-V) y 20 con MP por falciparum (MP-P) y como control se se leccionaron 19 muestras sin MP (MP-no). La prueba diag nóstica de infección plasmodial fue la reacción en cadena de la polimerasa cuantitativa en tiempo real (qPCR) en sangre placentaria.
Las mujeres y sus placentas fueron captadas en 2009 y 2016, en los hospitales de Puerto Libertador y Tierralta, municipios al sur del departamento de Córdoba, noroccidente de Colombia, en una región de alta endemicidad de malaria 21. El sur de Córdoba, junto con Urabá Antioqueño y Bajo Cauca Antioqueño, conforman la región ecoepidemiológica que más casos de malaria genera anualmente en Colombia 21.
Los criterios de inclusión fueron tener residencia permanente en esta región por lo menos durante el último año; no tener antecedentes de preeclampsia/eclampsia, enfermedad hipertensiva del embarazo, diabetes, VIH, y toxoplasmosis, rubeola, citomagalovirus, herpes simple (TORSCH); y tener parto con gestación de 36 a 41 semanas en alguno de los hospitales.
La edad gestacional se tomó de la historia clínica. Las mujeres del grupo control debían estar afebriles y las del grupo con infección plasmodial podían o no tener fiebre (solo 1% tuvo fiebre); asimismo deberían aceptar voluntariamente participar en el estudio. Los criterios de exclusión fueron retirar el consentimiento o la aparición de alguna complicación o enfermedad.
Diagnóstico de malaria
La sangre periférica materna se obtuvo en el momento del parto. La placenta se lavó con solución salina (0,9%) y de su cara materna se tomaron muestras de sangre y tejido para diagnóstico (gota gruesa, qPCR, histopatología) como se ha indicado en otros informes 22 - 25. Las muestras de gota grue sa fueron leídas por los investigadores y se definieron como negativas si 200 campos, con 100 aumentos, estuvieron li bres de parásitos.
El ADN se extrajo con el método de saponina-Chelex y la qPCR 22 , 23. El qPCR se ejecutó en la plataforma ABI 7500 FAST. Las muestras con un umbral de ciclo (Ct) <45 se analizaron en reacciones específicas de especies dúplex para P. falciparum y P. vivax 23. Se usó la amplificación de los genes 18S rRNA del ADN para la cuantificación. El número de copias de ADN se cuantificó a partir de la reacción específica del género frente a una curva estándar utilizando un plásmido que contiene un fragmento del gen 18S de P. falciparum.
Estudio histopatológico
El tejido placentario se procesó según procedimientos estan darizados 24 , 25. De cada placenta se tomaron dos fragmen tos y de cada uno se hizo una placa para estudio histológi co por microscopía de luz con procedimientos clásicos. Un fragmento procedió de un punto cercano a la inserción del cordón umbilical y el otro de la zona media (equidistante entre el cordón y el borde placentario). En total se leyeron 40 campos (20 por fragmento). La lectura fue cerrada frente a los resultados de gota gruesa y qPCR. Se usó aumento to tal de 400X para lectura histológica general. Se usó aumento total de 1000X para determinar la presencia de eritrocitos infectados (Ei) o hemozoína.
Cuantificación de la expresión de genes asociados a señalizadores/mediadores
Para cuantificar la expresión de genes asociados a señalizado res/mediadores y citoquinas en tejido placentario se usó un fragmento tisular preservado con RNA Later® (Qiagen) a 4 °C. Los mediadores se agruparon así: proinflamatorios (IL-2, TNF, IFNɣ, COX-1, COX-2, C5a), antiinflamatorios (IL-10, IL-4), angiogénicos (VEGF, VEGFR-1) y de hipoxia (HIF).
Los diferentes señalizadores, excepto C5a, se midieron por cuantificación relativa del ARNm mediante qPCR. Se hizo cuantificación relativa para determinar los niveles de expresión de los mediadores del estudio en relación con los niveles de expresión del gen constitutivo y luego de obtener la relación entre el gen de interés y el gen constitutivo. Se aplicó el procedimiento de Pfaffl para determinar el delta-delta CT como aparece a continuación 26:

Para cuantificar la fracción C5a del complemento en las muestras de suero placentario, se usó el estuche comercial Human C5a Elisa Kit, siguiendo las recomendaciones del fabricante.
Fuentes secundarias de información y grupos de estudio
Después de la inclusión, se aplicó un cuestionario clíni co-epidemiológico. La historia clínica fue usada como fuen te de datos. Se evaluaron parturientas y sus placentas, repar tidas en tres grupos: 19 en el grupo sin MP (MP-no) o grupo control, 20 en el grupo con MP-falciparum (MP-F) y 20 en el grupo con MP-vivax (MP-V).
Análisis estadístico
Se usaron los programas SPSS 18.0 y GraphPad Prism 5. Las decisiones sobre significación se tomaron con una probabilidad menor de 0,05. Se usó la prueba de Kolmo gorov-Smirnov para evaluar la distribución normal de las variables cuantitativas. Para evaluar la homocedasticidad de las varianzas se usó la prueba de Levene. Se usó la prue ba de Mann-Whitney para determinar si había diferencia entre dos grupos independientes. Se aplicó una prueba no paramétrica de Kruskal-Wallis para determinar si había di ferencia entre tres grupos independientes, posteriormente, se usó la prueba de Dunn para identificar los pares de grupos que diferían.
RESULTADOS
El 95% (38/40) de los casos de MP corresponden a infección submicroscópica. De las gestantes, 25% (15/59) presentaron antecedentes de malaria durante la gestación actual.
El rango de la edad fue de 14 a 41 años y el promedio de los tres grupos fue similar (MP-no: 22; MP-F: 25; MP-V: 23). La edad gestacional promedio fue de 38,6 semanas (rango 36 a 41). Las gestaciones previas tuvieron promedio de 2,8. El 36% fueron primigestantes, el 20% secundigestantes y el 44% multigestantes (3 a 9). De los 59 partos, 5 fueron por cesárea.
Al momento del parto, la hemoglobina promedio fue de 11,1 g/dL. Las mujeres con MP tuvieron niveles menores de hemoglobina que las no infectadas (10,86 g/dL y 11,69 g/dL, respectivamente). La hemoglobina fue menor en el grupo de MP-V (10,75 g/dL) que en MP-F (10,97 g/dL), pero sin diferencia significativa (p = 0,506). El promedio de peso neonatal en el grupo MP-no fue 2974 g; en MP-F fue 2852 g y en MP-V de 2737 g. La diferencia de las medias de peso neonatal fue 237 g entre hijos del grupo MP-V y MP-no, y de 115 g entre hijos del grupo MP-V y MP-F.
El grupo control (MP-no) tuvo ausencia de necrosis y mayor frecuencia de aterosis, abrupcio y trombos que en los otros dos. El grupo MP-V presentó resultados similares a MP-F en aterosis, necrosis, infarto, depósitos de fibrina y trombos, pero menor cantidad de hemozoína y eritrocitos infectados. Los depósitos de fibrina estuvieron en todas las placentas. En el espacio intervelloso, 95-100% de las placentas presentaron hemorragia y 6 de cada 10 placentas mostraron trombos y calcificaciones. Existió diferencia significativa (p < 0,05) entre los tres grupos en abrupcio, nodos sincitiales y hemozoína. El grupo de MP-F generó dicha diferencia. En la comparación por especie, no hubo diferencia significativa (p > 0,05) (Tabla 1).
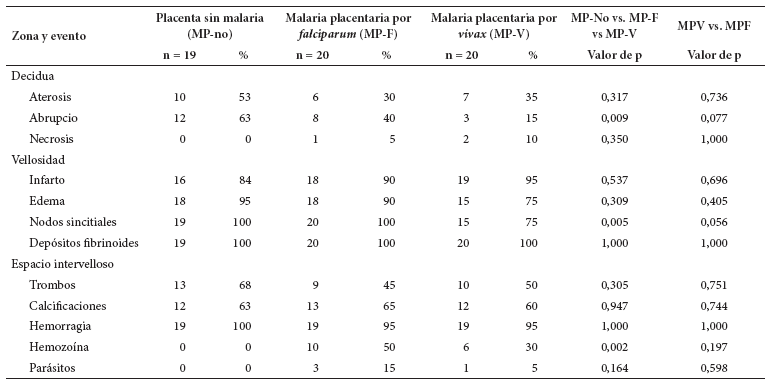
En el grupo MP-no, la aterosis y el abrupcio presentan una cantidad escasa, con promedios de 1,3 y 2,2 eventos. En ese mismo orden; infarto y edema vellositarios tuvieron 9,1 y 8,7 eventos en promedio. En el mismo grupo MP-no, la cantidad de depósitos de fibrina y de nodos sincitiales es de 72,5 y 109, como promedio. Las placentas sin infección tienen 366 vellosidades, 1463 capilares y 3,97 capilares por vellosidad. El promedio de hemorragias fue 18, mientras que los de trombos y calcificaciones fueron de dos (estuvieron en 60-70% de ellas) (Tabla 2).
Tabla 2 Magnitud de los eventos histológicos en placenta, según presencia de infección plasmodial
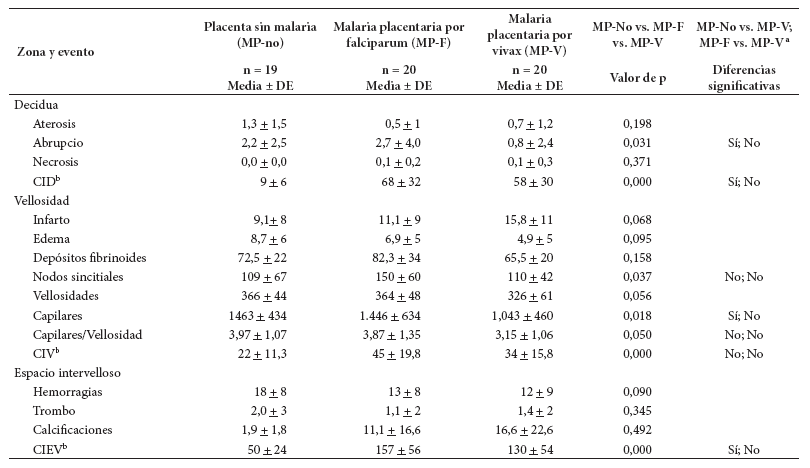
a Indica si la prueba de Dunn mostró diferencia entre cada par de comparaciones (MP-no vs. MP-V; MP-V vs. MP-F) con p < 0,05.
b CID: células inmunes en decidua. CIV: células inmunes en vellosidad. CIEV: células inmunes en espacio intervelloso. DE: Desviación estándar
En los grupos con MP-F y MP-V la aparición de la infección placentaria reduce la cantidad de aterosis, de edema vellositario, de capilares por vellosidad, de hemorragias y trombos, pero aumenta la de infarto vellositario, calcificaciones y células inmunes. Solo unos pocos eventos presentan diferencia significativa (Tabla 2).
Las células inmunes estuvieron en todas las placentas, en los tres compartimentos, e independientemente de la infección. Las cantidades fueron diferentes, en el grupo MP-no se observó 9 células inmunes en decidua, 22 en vellosidad y 50 en espacio intervelloso; en el grupo MP-V se observó 58, 34 y 130, en el mismo orden; en el grupo MP-F se encontró la mayor cantidad de células inmunes: 68, 45 y 157, respectivamente (Tabla 2).
En la Tabla 3 se muestra la expresión de mediadores inflamatorios, angiogénicos e hipóxicos por grupo. La inflamación (COX-1, COX-2, IL-10, IFNɣ, TNF, C5a) fue significativamente mayor en MP. IL-2 e IL-4 variaron poco entre los tres grupos. C5a, aunque no mostró diferencia estadísticamente significativa entre los grupos, presentó valores mayores en MP-V comparado con otros dos grupos. En la angiogénesis, VEGF y VEGFR-1 no presentaron diferencia significativa entre grupos, pero su expresión aumentó en grupos en MP. En la hipoxia, HIF-1α tuvo diferencia significativa, con mayores valores en MP. En general, los mediadores que no presentaron diferencia significativa entre tres grupos fueron IL-2, VEGF, VEGFR-1 y C5a. Para VEGFR-1 y C5a, la alta variabilidad intragrupo es, probablemente, un factor que afecta la significación estadística al comparar los tres grupos.
Tabla 3 Expresión de mediadores de inflamación, angiogénesis e hipoxia según grupo de estudio
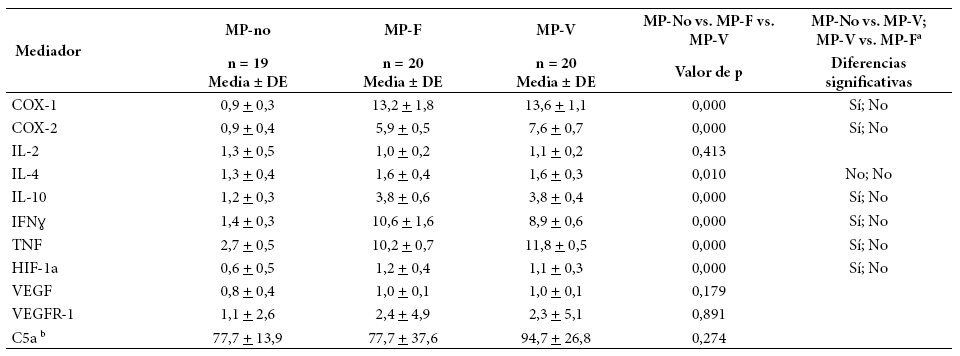
a Indica si la prueba de Dunn mostró diferencia entre cada par de comparaciones (MP-no vs MP-V; MPV vs. MP-F), con p < 0,05.
b Para C5a se midió su concentración en sangre y no la expresión de su gen asociado.
En las Figuras 1, 2 y 3 se muestran las correlaciones lineales bivariadas entre eventos histológicos y mediadores de procesos. En el grupo sin infección, es claro el predominio de correlaciones significativas (CS) negativas y débiles (p < 0,10). Los depósitos de fibrinas, los nodos sincitiales y los trombos reúnen la mayor cantidad de CS. Las seis CS que tienen los depósitos fibrinoides se hacen con siete mediadores de procesos, tanto promotores de inflamación (COX-2 [pero no COX-1], C5a, IFNɣ [pero no TNF], VEGF) como de hipoxia (HIF) y angiogénesis (VEGF, pero no su receptor). Las cinco CS de los nodos sincitiales se hacen con tres de los mediadores que también las poseen con los depósitos fibrinoides (COX-2, HIF y VEGF) y con las citoquinas 2 (proinflamatoria) y 10 (antiinflamatoria). Las cuatro CS de los trombos suceden con los mediadores de inflamación TNF, IL-2, VEGF y el mediador de hipoxia HIF.
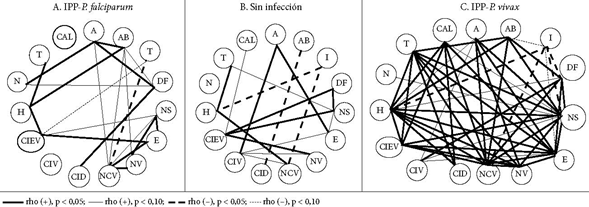
Figura 1 Correlaciones lineales bivariadas significativas (CS) entre eventos histológicos placentarios
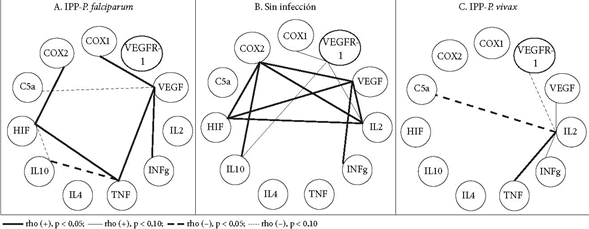
Figura 2 Correlaciones lineales bivariadas significativas (CS) entre mediadores de procesos placentarios de angiogénesis, inflamación e hipoxia
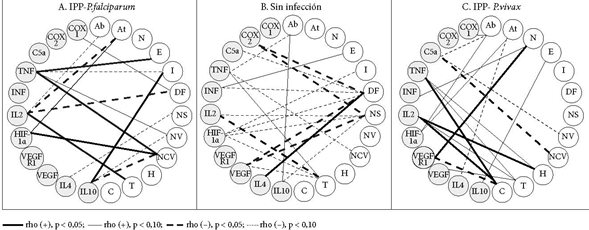
De otro lado, hay que resaltar las CS que establecen varios mediadores con algunos eventos histológicos. Es el caso de HIF, además de asociarse con los nodos sincitiales, los depósitos fribrinoides y los trombos, también lo está con infarto. TNF se relaciona con abrupcio, trombos y capilares vellositarios.
En el grupo MP-F, las CS se reducen y la polaridad cambia; ahora, los polos no son los nodos sincitiales, los depósitos fribrinoides y los trombos, sino el único polo que se destaca son los capilares vellositarios (tres CS: con TNF, HIF e IL-10). El TNF tuvo CS con edema, infarto, el número de vellosidades y el número de capilares por vellosidad; la IL-2 tuvo CS con abrupcio, aterosis, trombos y depósitos fibrinoides.
Para el grupo MP-V, las CS abundan como en la ausencia de infección, pero tiene otros centros: abrupcio, trombos, calcificaciones. Las CS del abrupcio se dan con COX-1, C5a e HIF-1α; las de trombos y calcificaciones con el TNF e IL-2. Además los trombos se asocian a VEGF, en cambio, las calcificaciones se relacionan con VEGFR-1.
DISCUSIÓN
En el presente trabajo se hallaron diferencias importantes en los efectos placentarios de las dos especies plasmodiales evaluadas. La cantidad de abrupcio, nodos sincitiales, vello sidades, capilares y capilares por vellosidad es significativa mente menor cuando hay P. vivax, respecto a P. falciparum. Por otra parte, P. vivax comparado con las placentas sin in fección, generó más infarto y menos abrupcio, edema, ve llosidades, capilares y capilares por vellosidad. La MP-IPPS debida a P. falciparum tiene pruebas sobre su naturaleza pa tógena en gestantes y sus hijos 1. Inclusive en estudios en la misma zona geográfica en donde se hizo este trabajo 27 - 29, la infección submicroscópica placentaria por P. vivax ocasiona efectos nocivos.
Las comparaciones del grupo de P. vivax contra P. falciparum o contra el grupo sin infección apoyan fuertemente la capacidad patogénica placentaria de P. vivax. Estos hallazgos sientan bases para explicar sus efectos por un mecanismo que no necesariamente involucra la citoadhesión de eritrocitos infectados al tejido placentario, como se da en infecciones por P. falciparum.
Existe escasa información sobre las correlaciones entre eventos histológicos y mediadores de procesos. Cuando no hay infección, las CS entre eventos histológicos y mediadores de procesos son predominantemente menores al 10%. Esto se puede interpretar como un estado homeostático, en el que no predomina una respuesta específica, sino que todas ellas se encuentran activas, pero en estados basales y equilibrados. En caso contrario la gestación estaría en riesgo.
Cuando existe infección plasmodial, el panorama de las CS cambia en forma radical, tanto cuando se trata de infección plasmodial placentaria por P. falciparum (IPP-F), pero sobre todo en infección plasmodial placentaria por P. vivax (IPP-V). Las CS que había en el estado de no infección desaparecen y surgen otras nuevas en las infecciones plasmodiales. Los centros o polos de CS que eran los nodos sincitiales y los depósitos fibrinoides desaparecen y son reemplazados por polos que ahora pertenecen a los mediadores de procesos, especialmente de inflamación. La IL-2, el TNF e la IL-10 son polos en IPP-F, mientras que la IL-2, el TNF y el C5a lo son en IPP-V. En ausencia de infección, las vellosidades y los capilares están prácticamente sin CS, cuando aparece infección ellos surgen como polos. Todo esto expresaría el estado inflamatorio exacerbado de la placenta como consecuencia de la infección.
En el presente estudio no se midieron hormonas en ninguno de los trimestres de la gestación. La progesterona, estrógenos, andrógenos y glucocorticoides están involucrados en la gestación desde la implantación hasta el parto; su biosíntesis y su metabolismo son el resultado de vías complejas que involucran al feto, la placenta y la madre 30. Muchas hormonas sexuales tienen clara interacción con el sistema inmune y muchos mediadores de procesos fisiológicos también interactúan con tal sistema. Existe, entonces, una compleja red fisiológica entre hormonas, mediadores de procesos fisiológicos y sistema inmune.
Entre las fortalezas del estudio está su aproximación integral a la MP, abordando la relación entre eventos histopatológicos y mediadores de inflamación, angiogénesis e hipoxia. A nuestro conocimientos estas relaciones no han sido estudiadas. Se tuvo un grupo control bien definido, el cual fue negativo para Plasmodium según técnica altamente sensible y específica (qPCR), sin presencia de TORCHS, VIH, eclampsia-preeclampsia, y diabetes. Sin embargo, no se pudo descartar la presencia de parásitos intestinales y desnutrición.
Entre las limitaciones señalamos el pequeño tamaño muestral y la selección por conveniencia, generado por las condiciones de tiempo y dinero disponibles. Los mediadores evaluados representan básicamente el proceso inflamatorio de la placenta a término, otros mediadores de procesos no fueron medidos suficientemente. El análisis de las correlaciones entre hallazgos histológicos y mediadores de procesos es superficial porque la información disponible es escasa o inexistente.
En conclusión, cuando ocurre IPPS hay cambios tisulares placentarios y cambios en la expresión de mediadores del proceso inflamatorio, tanto si el agente causal es P. falciparum como si es P. vivax. Si es que el parásito es patógeno para la placenta, cabe esperar que cuando las parasitemias se incrementen, más fuertes serán sus efectos. La infección placentaria por P. vivax contribuye al aumento de hallazgos histológicos placentarios asociados con deterioro y daño del tejido. La alteración de la estructura placentaria estuvo principalmente asociada a la disminución de vellosidades y del número de capilares por vellosidad, así como con el aumento de lesiones degenerativas isquémicas asociados con calcificaciones e infarto. Estos resultados podrían correlacionarse con disminución del peso al nacer y con disminución en los niveles de hemoglobina.

