Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los reptiles y los anfibios se encuentran entre los grupos de vertebrados más amenazados en todo el mundo (Stuart et al. 2004), pero también, en particular los reptiles son considerados uno de los taxones más relevantes ecológica y evolutivamente, porque han colonizado con éxito gran diversidad de hábitats, incluidos los océanos y algunos ecosistemas extremos y ambientalmente más inestables de la tierra (Pincheira-Donoso et al. 2013). La herpetofauna es también considerada un grupo sensible a los cambios ambientales (Pianka 1967, Miller et al. 2018) además, son efectivos taxones sustitutos de riqueza que permiten evaluar apropiadamente la compleja biodiversidad de un lugar (Lewandowski et al. 2010), es decir, cuanto mayor es la riqueza de anfibios y reptiles, mayor sería la riqueza de otros grupos taxonómicos difíciles de evaluar en un área determinada.
Los listados de especies y estudios taxonómicos son herramientas básicas para una adecuada gestión de la biodiversidad. Así mismo, la recopilación de toda la información concerniente a estudios taxonómicos y distribución de especies es fundamental para acciones de conservación como los listados de especies amenazadas y consecuentemente mejores decisiones en políticas de conservación (IUCN 2022). Sin embargo, varios departamentos del Perú no cuentan con estas elementales herramientas, y en algunos casos ni siquiera para los grupos más importantes como las plantas o vertebrados, incluidos los anfibios y reptiles.
Uno de estos departamentos es Ica, que a pesar de los nuevos registros y descripciones de nuevas especies herpetológicas en los últimos años (Venegas et al. 2017, Aguilar-Puntriano et al. 2019, Mendoza et al. 2021), no se ha sintetizado la información de distribución y trabajos taxonómicos que permitan identificar un conocimiento de la biodiversidad actual de reptiles y anfibios. Adicionalmente, cabe mencionar que el último listado de especies de reptiles y anfibios que proporciona información del departamento de Ica data de 1995 (Carrillo & Icochea 1995) y la última categorización de especies a nivel nacional (basada principalmente en distribución de especies y estudios taxonómicos) fue dada en el 2014 con el DS Nº 004-2014-MINAGRI.
En este trabajo se presenta una lista actualizada de reptiles y anfibios, se comentan los nuevos registros, se ilustra la distribución de las especies y se presentan claves taxonómicas para anfibios y reptiles del departamento de Ica. Por último, se discuten los límites de la distribución de la herpetofauna de la vertiente occidental de los Andes y el desierto costero (Desierto semicálido tropical según Britto 2017) en el sur de Perú.
Material y métodos
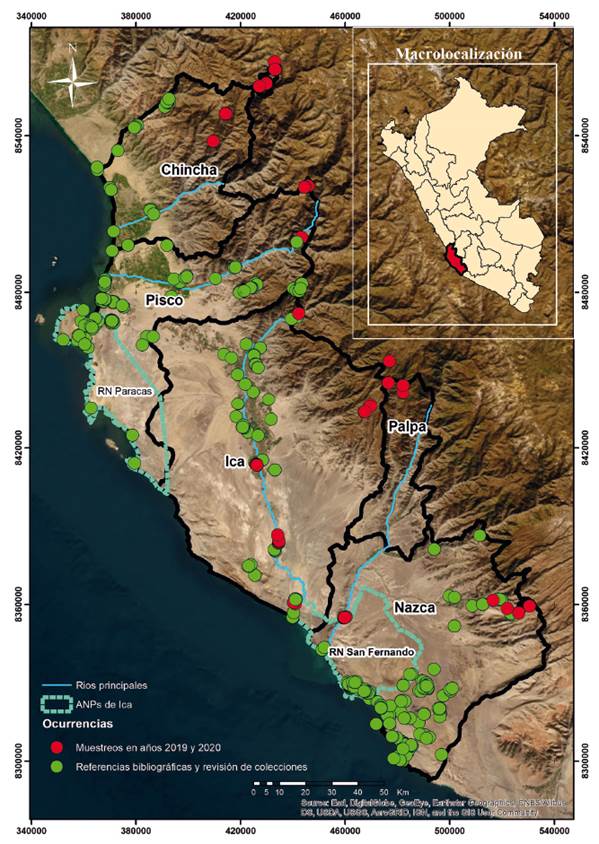
La ocurrencia de especies de anfibios y reptiles en el departamento de Ica fueron recopilados de artículos científicos y tesis (Pérez & Balta 2007, Pérez & Balta 2011, Venegas et al. 2017, Arapa 2018, Aguilar et al. 2019, Mendoza et al. 2021, Ormeño et al. 2021), los registros de la colección del Departamento de Herpetología del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (DH-MUSM) y del Centro de Ornitología y Biodiversidad (CORBIDI), registros del Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa (MUSA), seis informes técnicos de gestión ambiental depositados en el SERFOR y 95 expedientes del SENACE. Estas últimas fuentes de información se revisaron escrupulosamente para verificar el reporte, confirmando la identificación de las especies considerando reportes con evidencia fotográfica, muestras depositadas en museos o registros previos en la zona de evaluación de informe.
Además, se realizaron evaluaciones de campo en zonas con vacíos de información en las provincias de Ica, Chincha, Nazca, Palpa y Pisco entre los años 2019 y 2020 (Anexo 1 y Anexo 3). Durante estas evaluaciones de campo se realizaron colectas y los ejemplares fueron depositados en el DH-MUSM. Se determinaron taxonómicamente las especies reportadas teniendo como referencia la colección de especímenes del DH-MUSM y el uso de bibliografía y claves especializadas (Peters & Orejas-Miranda 1970; Peters & Donoso Barros 1970; Dixon & Wright 1975, Langstroth 2011; Aguilar et al. 2007; Torres-Carbajal 2007a; Torres-Carbajal 2007b). También se recurrió a información de páginas especializadas como The Reptile Database (Uetz et al. 2021), Amphiaweb (AmphibiaWeb 2022) y la lista roja de especies amenazadas (IUCN Red List of Threatened Species, IUCN 2022).
Se generaron mapas en base a toda la información compilada con los puntos y coordenadas de presencia de especies de reptiles y anfibios. Se utilizó la aplicación ArcMapTM 10.1 disponible en el programa ArcGIS 10.1 para generar los mapas de distribución. Se utilizó el mapa con la información bibliográfica para seleccionar las áreas con vacíos de información como paso previo al trabajo de campo.
Las claves de identificación de especies de anfibios y reptiles fueron realizadas utilizando las características diagnósticas mencionadas en otras publicaciones (Peters & Orejas-Miranda 1970; Peters & Donoso Barros 1970; Dixon & Huey 1970; Dixon & Wright 1975; Aguilar et al. 2007; Torres-Carbajal 2007b) y nuestras propias observaciones utilizando la colección del DH-MUSM.
Se recopiló información de distribución geográfica de literatura herpetológica (ver citas en la Discusión) y de la colección del DH-MUSM para examinar los patrones de distribución de los anfibios y reptiles, en la zona del desierto costero y vertiente occidental del departamento de Ica.
Por razones de espacio solo se incluye información de distribución desde Ancash (limite norte) hasta el norte de Chile (limite sur); aunque algunas especies pueden distribuirse más allá de ambos limites, incluso en otros países. Los resultados de esta sección se muestran en la Discusión.
Resultados
Se registraron veintidos especies: tres anfibios pertenecientes a las familias Bufonidae, Telmatobiidae y Leptodactylidae respectivamente (Tabla 1); y diecinueve reptiles de las familias Tropiduridae, Liolaemidae, Phyllodactylidae, Teiidae, Colubridae, Viperidae y Leptotyphlopidae (Tabla 1). La distribución de las especies registradas muestra en las Figuras del 3 al 5.
Tabla 1 Anfibios y reptiles registrados en el departamento de Ica. Categoría de protección por el Estado Peruano (D.S. 004 2014 MINAGRI) o IUCN: VU = Vulnerable, NT = Casi Amenazado; Presencia en un área natural protegida (ANP): CP = Concesión Privada, RN = Reserva Nacional, RP = Reservada Paisajística, SIP = Sistema de Islas y Puntas, SN = Santuario Nacional, ZR = Zona Reservada. * = Nuevos registros.
| Clase | Orden | Clado | Familia | Especie | Distribución restringida | Protegido/IUCN | ANP | |
|---|---|---|---|---|---|---|---|---|
| AMPHIBIA | ||||||||
| Anura | ||||||||
| Bufonidae | Rhinella limensis (Werner, 1901) | RN San Fernando | ||||||
| Telmatobiidae | Telmatobius rimac* Schmidt, 1954 | Lima, Ica | VU (IUCN) | RP Nor Yauyos-Cochas | ||||
| Leptodacylidae | Pleurodema marmoratum* (Duméril and Bibron, 1840) | VU (IUCN) | RN Salinas y Aguada Blanca y Pampa Galeras | |||||
| REPTILIA | ||||||||
| Squamata | ||||||||
| Iguania | ||||||||
| Tropiduridae | Microlophus peruvianus (Lesson, 1830) | RN Paracas y San Fernando, SIP | ||||||
| Tropiduridae | Microlophus tigris (Tschudi, 1845) | NT (DS 004 2014 MINAGRI) | RN Lachay, ZR Ancón | |||||
| Tropiduridae | Microlophus thoracicus (Tschudi, 1845) | Ica | RN Paracas y San Fernando | |||||
| Tropiduridae | Microlophus theresiae (Steindachner, 1901) | RN Paracas y San Fernando | ||||||
| Tropiduridae | Stenocercus icaMendoza, Ramírez, Barrera & Aguilar-Puntriano, 2021 | Ica | ||||||
| Tropiduridae | Stenocercus ornatissimus* (Girard, 1858) | Lima, Ica | NT (IUCN) | ZR Bosque de Zarate | ||||
| Liolaemidae | Ctenoblepharys adspersa Tschudi, 1845 | Lima, Ica | VU (IUCN); VU (DS 004 2014 MINAGRI) | RN Paracas, San Fernando, ZR Ancón, RN Lachay | ||||
| Liolaemidae | Liolaemus evaristoi* Gutiérrez, Chaparro, Vásquez, Quiroz, Aguilar-Kirgin & Abdala, 2018 | Ica, Huancavelica | ||||||
| Liolaemidae | Liolaemus nazca Aguilar, Ramírez, Castillo, Mendoza, Vargas & Sites Jr., 2019 | Ica | RN San Fernando | |||||
| Lacertoidea | ||||||||
| Teiidae | Dicrodon heterolepis (Tschudi, 1845) | NT (DS 004 2014 MINAGRI) | ZR de Illescas, RN San Fernando | |||||
| Gekkota | ||||||||
| Phyllodactylidae | Phyllodactylus gerrhopygus (Wiegmann, 1834) | RN Paracas y San Fernando, SIP | ||||||
| Phyllodactylidae | Phyllodactylus angustidigitusDixon and Huey, 1970 | Ica | NT (IUCN) | RN Paracas | ||||
| Phyllodactylidae | Phyllodactylus lepidopygus (Tschudi, 1845) | VU (IUCN); VU (DS 004 2014 MINAGRI) | RN Lachay | |||||
| Phyllodactylidae | Phyllodactylus cf. sentosusDixon and Huey, 1970 | Ica | RN San Fernando | |||||
| Serpentes | ||||||||
| Colubridae | Pseudalsophis elegans (Tschudi, 1845) | RN Paracas y San Fernando | ||||||
| Colubridae | Incaspis tachymenoides* (Schmidt & Walker, 1943) | SN Lagunas de Mejía y CP Lomas de Atiquipa | ||||||
| Colubridae | Oxyrhopus fitzingeri (Tschudi, 1845) | |||||||
| Viperidae | Bothrops pictus (Tschudi, 1845) | VU (DS 004 2014 MINAGRI) | RN San Fernando | |||||
| Leptotyphlopidae | Epictia tesselata* (Tschudi, 1845) | Lima, Ica | ||||||
Nuevos registros (Figuras 1-2)
Clase Amphibia
Orden Anura
Familia Telmatobiidae Fitzinger, 1843
Telmatobius rimac Schmidt, 1954
El primer registro fue un grupo de renacuajos y una hembra adulta (MUSM 40457) en un riachuelo entre pajonales y matorrales andinos a 3 km rio arriba del distrito San Pedro de Huacarpana (provincia de Chincha) con coordenadas WGS84 UTM zona 18 L Este 429664 Norte 8560397 a 4001 m. También se encontraron varios renacuajos y cinco ejemplares adultos (MUSM 40485 a MUSM 40489) en un pequeño arroyo con pozas en Huacarpana, con coordenadas 18 L Este 414151 Norte 8548561 a 3848 m. Además, se encontraron renacuajos (MUSM 40528) y cinco adultos (MUSM 40529, 40530, 40531, 40532, 40533) en una orilla con rocas del rio Tibillo (distrito de Tibillo, provincia de Palpa).
Familia Leptodactylidae Werner, 1896
Pleurodema marmoratum (Duméril & Bibron, 1840)
Se registraron cuatro individuos (MUSM 40458, 40459, 40460, 40461) bajo piedras al borde de un pequeño riachuelo, con coordenadas 18 L Este 429764 Norte 8560362 a 4031 m y otros dos individuos (MUSM 40462 y 40463), ambos a cuatro kilómetros del centro poblado San Pedro de Huacarpana (provincia de Chincha). También se reportaron dos individuos (MUSM 40498 y 40499) bajo piedras en la zona más alta de la misma localidad, con coordenadas 18 L Este 433030 Norte 8568441 a 4430 m.
Clase Reptilia
Orden Squamata
Familia Colubridae Oppel, 1811
Incaspis tachymenoides (Schmidt & Walker, 1943)
El único registro fue de un individuo adulto (MUSM 40490), desplazándose entre rocas, rodeadas de matorrales en el distrito de San Juan de Yanac en la provincia de Chincha con coordenadas 18 L Este 409435 Norte 8537940 a 3213 m.
Familia Leptotyphlopidae Stejneger, 1892
Epictia tesselata (Tschudi, 1845)
Se encontró el registro de un individuo adulto (MUSM 37859) en la revisión de la colección del DH-MHNSM, reportada en las zonas altas de la provincia de Pisco, en la localidad de Huancacasa.
Familia Tropiduridae Bell, 1843
Stenocercus ornatissimus (Girard, 1858)
Se obtuvo el registro de 15 individuos entre machos, hembras y juveniles (MUSM 40464, 40465, 40466, 40477, 40478, 40479, 40480, 40481, 40482, 40511, 40512, 40513, 40514, 40515, 40516), encontrados principalmente bajo rocas, pero también desplazándose sobre piedras entre matorrales altoandinos y roquerios. Los registros fueron realizados en los distritos de San Pedro de Huacarpana en la provincia de Chincha (18 L Este 414553 Norte 8548484 a 3875 m.) y Huancano en la provincia de Pisco (18 L Este 445748 Norte 8521189 a 3549 m).
Familia Liolaemidae Frost & Etheridge, 1989
Liolaemus evaristoiGutiérrez, Chaparro, Vásquez, Quiroz, Aguilar-Kirgin & Abdala, 2018
Se reportaron individuos machos, hembras y juveniles (MUSM 40445, 40446, 40447, 40448, 40449, 40450, 40451, 40452, 40453, 40454, 40455, 40456, 40491, 40492, 40493, 40494, 40495, 40496, 40497, 40500, 40501, 40502, 40503, 40504, 40505, 40506, 40507, 40508, 40509, 40510), encontrados principalmente bajo rocas entre matorrales y pajonales cercanos a un arroyo. Los reportes fueron en la zona más alta del distrito San Pedro de Huacarpana (provincia de Chincha), con coordenadas 18 L Este 429674 Norte 8560264 a 3998 m.

Figura 1 Nuevos registros de anfibios y serpientes para el departamento de Ica. 1 Pleurodema marmoratum, 2 Telmatobius rimac, 3 Epictia tesselata y 4 Incaspis tachymenoides.

Figura 2 Nuevos registros de saurios para en el departamento de Ica. 1 Microlophus sp., 2 Stenocercus ornatissimus, 3 Phyllodactylus cf. sentosus y 4 Liolaemus evaristoi.
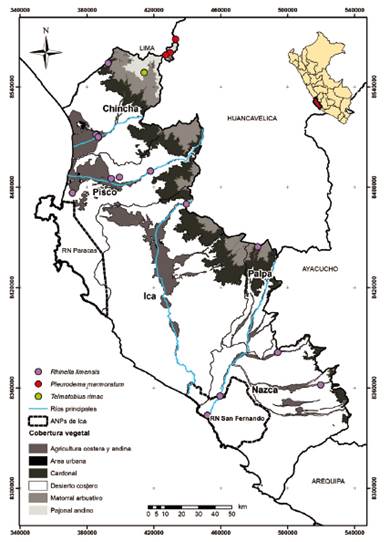
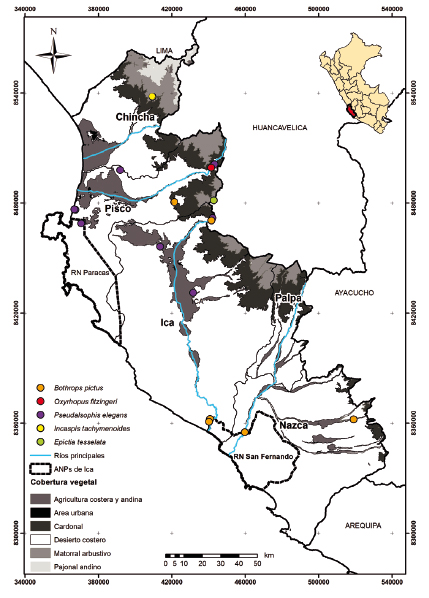
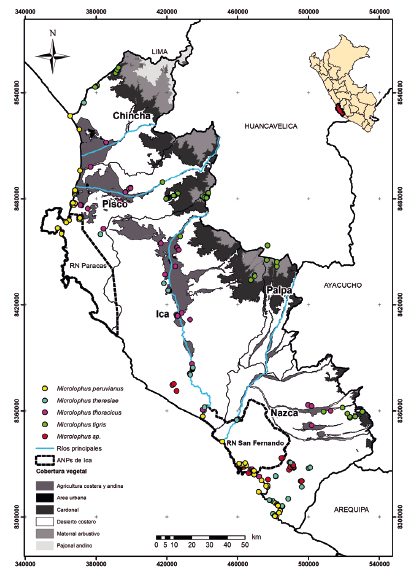
Figura 5 Distribución de saurios del género Microlophus por cobertura vegetal en el departamento de Ica.
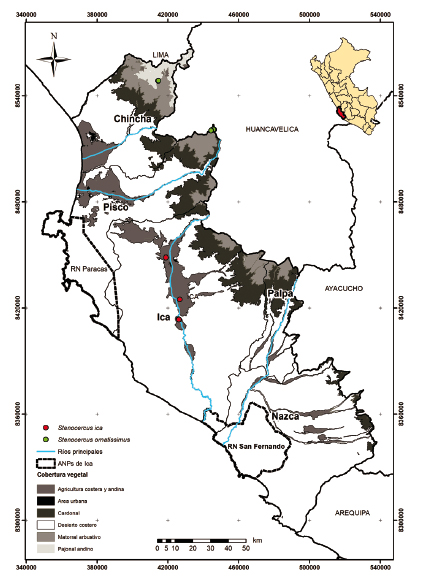
Figura 6 Distribución de saurios del género Stenocercus por cobertura vegetal en el departamento de Ica.
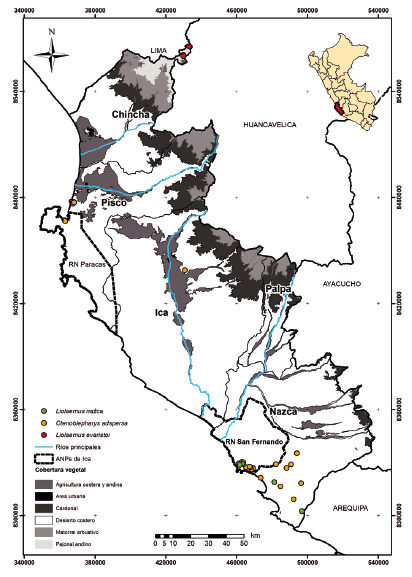
Figura 7 Distribución de saurios de la familia Liolaemidae por cobertura vegetal en el departamento de Ica.
Clave para los anfibios del departamento de Ica
1a. Glándulas parotoideas presentes, con formaciones glandulares y tubérculos dorsolaterales. Rhinella limensis
1b. Glándulas parotoideas ausentes. 2
2a. Piel dorsal sin tubérculos, con glándulas redondeadas o alargadas, zona ventral de muslos sin coloración amarilla en vida. Pleurodema marmoratum
2b. Piel dorsal sin tubérculos, sin glándulas visibles, zona ventral de muslos con coloración amarilla en vida. Telmatobius rimac
Clave para los saurios del departamento de Ica
1a. Con escamas grandes en la cabeza, no granulares, pupilas circulares. 2
1b. Con escamas pequeñas granulares en la cabeza y dorso, pupilas verticales. 11
2a. Con escamas grandes laminares en la cabeza, superciliares yuxtapuestas no en diagonal, escamas del vientre rectangulares más grandes que las dorsales, con lengua bifurcada, cabeza roja en vida en machos adultos. Dicrodon heterolepis
2b. Con escamas de diferente tamaño en la cabeza, superciliares imbricadas en diagonal, escamas del vientre de igual o mayor tamaño que las dorsales, lengua no bifurcada. 3
3a. Escama interparietal de tamaño similar que las escamas circundantes, machos con o sin poros preanales. 4
3b. Escama interparietal grande, aproximadamente 1/3 del ancho de la cabeza, machos sin poros preanales. 8
4a. Machos sin poros precloacales. 5
4b. Machos con poros precloacales. 6
5a. Ausencia de pliegue oblicuo en el cuello. Stenocercus ica
5b. Presencia de pliegue oblicuo en el cuello. Stenocercus ornatissimus
6a. Ciliares (“peines”) bien desarrolladas en vista dorsal y lateral, hocico truncado. Ctenoblepharys adspersa
6b. Ciliares no desarrolladas en vista dorsal y lateral, hocico no truncado, punteagudo. 7
7a. Con manchas negras en la gula, vientre en vida naranja con manchas negras y flancos con verde esmeralda en machos adultos. Liolaemus nazca
7b. Sin manchas negras o poco notorias en la gula, vientre en vida amarillo en machos y escamas celestes en el dorso y cola. Liolaemus evaristoi
8a. Dos o más hileras de escamas entre la escama nasal y primera supralabial, mentón blanco o con bandas o líneas grises. Microlophus thoracicus
8b. Una hilera de escamas entre la escama nasal y primera supralabial. 9
9a. Escamas de la fila media del dorso no agrandada, machos con halo naranja alrededor del ojo en vida y en hembras naranja opaco. Microlophus theresiae
9b. Escamas de la fila media del dorso agrandada, sin halo naranja alrededor del ojo en vida. 10
10a. Palmas de las manos completamente quilladas, con manchas negras en la gula de machos que pueden fusionarse y escamas dorsales del antebrazo levemente quilladas. Microlophus tigris
10b. Palmas de las manos completamente lisas o parcialmente quilladas, con chevrones negros en la gula de machos y usualmente escamas dorsales del antebrazo lisas o no quilladas. Microlophus peruvianus
11a. Con presencia de una sola placa anal muy alargada, 10 a 20 veces el tamaño de otras escamas ventrales. 12
11b. Sin placa anal, escamas cerca de la cloaca de igual en tamaño a otras escamas ventrales. 13
12a. Dígitos cortos, con lamelas terminales ensanchadas con uña no visible. Phyllodactylus gerrhopygus
12b. Dígitos largos, con lamelas terminales delgadas con una uña visible. Phyllodactylus angustidigitus
13a. Ausencia de tubérculos grandes dorsales en la tibia. Phyllodactylus lepidopygus
13b. Presencia de tubérculos grandes dorsales en la tibia. Phyllodactylus cf. sentosus
Clave para serpientes del departamento de Ica.
1a. Cabeza y cola similares, ojos vestigiales o muy reducidos, escamas ventrales del mismo tamaño que las dorsales. Epictia tesselata
1b. Cabeza, cola y ojos diferenciados, escamas ventrales agrandadas, de mayor tamaño que las dorsales. 2
2a. Presencia de una foseta loreal entre el ojo y la narina, escamas quilladas en el dorso, escamas pequeñas detrás del ojo. Bothrops pictus
2b. Ausencia de una foseta loreal entre el ojo y la narina, escamas lisas en el dorso, escamas generalmente grandes detrás del ojo. 3
3a. Diseño dorsal pardo oscuro con manchas irregulares blancas, amarillas o naranjas en vida, escama anal no dividida, ojos completamente visibles dorsalmente. Oxyrhopus fitzingeri
3b. Diseño dorsal no como el anterior, escama anal dividida, ojos parcialmente visibles dorsalmente. 4
4a. Diseño dorsal con manchas en zig-zag o fusionadas a lo largo del cuerpo, formando bandas oscuras y claras desde la mitad posterior del cuerpo, o formando bandas oscuras y claras a lo largo del cuerpo. Pseudalsophis elegans
4b. Diseño dorsal con manchas pares a lo largo del cuerpo con manchas laterales que pueden fusionarse anteriormente formando una banda, o con diseño dorsal uniforme con una banda lateral más oscura. Incaspis tachymenoides
Discusión
Nuevos registros para el departamento de Ica. Antes del presente trabajo en el departamento de Ica se conocía solo una especie de anfibio, nueve de lagartijas y cuatro de serpientes, totalizando 14 especies (Rodriguez et al. 1993, Carillo e Icochea, 1995). En el presente estudio se reportan 22 especies (tres anfibios, 14 lagartijas y cinco serpientes). Los nuevos registros no publicados para el departamento de Ica son los anfibios Telmatobius rimac y Pleurodema marmoratum, las lagartijas Liolaemus evaristoi, Stenocercus ornatissimus, y dentro de las serpientes el colúbrido Incaspis tachymenoides y la serpiente ciega Epictia tesselata. El incremento obtenido en registros de especies de reptiles y anfibios se debió principalmente a las evaluaciones de campo que fueron actividades complementarias a la búsqueda bibliográfica (Anexo 1 y Anexo 3).
Los géneros predominantes en número de especies y registros fueron Microlophus (Tropiduridae) y Phyllodactylus (Phyllodactylidae). La mayoría de las especies de ambos géneros se encuentran en hábitats costeros, en playas, lomas y valles. Sin embargo, Microlophus tigris y Phyllodactylus lepidopygus pueden llegan a más de los 1500 m de altitud (Carrillo & Icochea 1995, Lehr et al. 2002, Pérez & Balta 2007, este estudio). En ambos géneros se encuentran poblaciones con incertidumbre taxonómica como Microlophus sp. (Figs. 2 y 5) y Phyllodactylus cf. sentosus (Figs. 2 y 5). Microlophus sp. está reportada principalmente en las lomas de la Reserva Nacional San Fernando (Arapa 2017, RSF, CA, AM, JPZ y CR observaciones personales). Esta población presenta un patrón de coloración que por una parte recuerda a M. peruvianus, reportada principalmente en playas rocosas y acantilados (Quispitúpac & Pérez 2008) y por otra a M. tigris, reportada generalmente en hábitats de Lomas (Carrillo & Icochea 1995, Ruiz et al. 2017). Los registros de Microlophus sp. son sólo fotográficos y no permiten examinar otras características fenotípicas que harían posible su asignación a una especie conocida de Microlophus. También se registró con el nombre de P. sentosus a una población ubicada en las desembocaduras del río Ica (Venegas et al. 2017) y río Grande en Nasca (Arapa 2018). Antes del 2017, esta especie solamente era conocida para un área muy restringida del departamento de Lima (Pérez & Balta 2016), existiendo una gran distancia (350 km aproximadamente) entre las localidades de Lima y los registros en el departamento de Ica (Arapa 2018, Venegas et al. 2017). Sin embargo, análisis moleculares y nueva evidencia fenotípica no publicada (JPZ observación personal) sugieren que las poblaciones al sur de Ica pueden representar un nuevo linaje, pero hasta que no se completen estos estudios sobre delimitación de especies siguiendo un enfoque integrativo (como por ejemplo Aguilar et al. 2013) estamos considerando a esas poblaciones como Phyllodactylus cf. sentosus.
Otros géneros de saurios muestran menor número de especies en la región de Ica. Por ejemplo, en Stenocercus (Tropiduridae) mediante la recopilación de información en las colecciones científicas y las evaluaciones de campo complementarias se obtuvo el registro y descripción de una nueva especie, Stenocercus ica (Mendoza et al. 2021). Otro nuevo registro es Stenocercus ornatissimus, la cual fue reportada en las evaluaciones de campo en la sierra de la provincia de Pisco a 3500 m aproximadamente. En Liolaemus se reportan dos especies para el departamento de Ica. Una de ellas es L. nazca, reportada para las lomas de Marcona (Aguilar et al. 2019) y en las lomas de la Reserva Nacional de San Fernando (Arapa 2018). Asimismo, L. evaristoi fue descrita para la provincia de Huaytara en Huancavelica (Gutiérrez et al. 2018). Adicionalmente, en las evaluaciones de campo se obtuvo un nuevo registro en el centro poblado de San Pedro de Huacarpana en la provincia de Chincha a 4200 m aproximadamente.
Con respecto a las serpientes, ya se tenían reportes de las especies Bothrops pictus, B. roedingeri (Viperidae), Pseudalsophis elegans y Oxyrhopus fitzingeri (Colubridae), pero la revisión de bibliografía permitió obtener datos geográficos más precisos de estas especies (Carrillo & Icochea 1995, Lehr et al. 2002, Arapa 2018). Además, se obtuvo el registro de Epictia tesselata (Leptotyphlopidae), comúnmente llamada “serpiente ciega”, en las zonas altas de la provincia de Pisco, en la localidad de Huancacasa (Vargas 2015) y de Incaspis tachymenoides en la evaluación de campo complementaria en el distrito de San Juan de Yanac (Chincha). Un caso especial es el estado taxonómico de B. roedingeri, para el cual diferentes autores la consideran un sinónimo de B. pictus (Campbell & Lamar 1992) o un linaje independiente (Fenwick et al. 2009, Carrasco et al. 2012). En este estudio consideramos a B. roedingeri como un sinónimo de B. pictus.
Con respecto a los anfibios (Anura) se contaba con el único registro del “sapo” Rhinella limensis (Bufonidae) (Rodriguez et al. 1993). En la revisión bibliográfica se pudieron obtener coordenadas de ubicación de esta especie dentro del departamento de Ica. Sin embargo, gran parte de la información estaba principalmente distribuida en hábitats costeros y desérticos, lo cual limitaba el posible registro de especies de anfibios en zonas más altas. En las evaluaciones de campo complementarias se priorizó evaluar particularmente las zonas altas del departamento de Ica, donde se registraron a Telmatobius rimac (Telmatobiidae) y Pleurodema marmoratum (Leptodactylidae) en la provincia de Chincha. Telmatobius rimac también se registró en la provincia de Palpa, al sur del departamento de Ica, siendo esta su localidad más al sur.
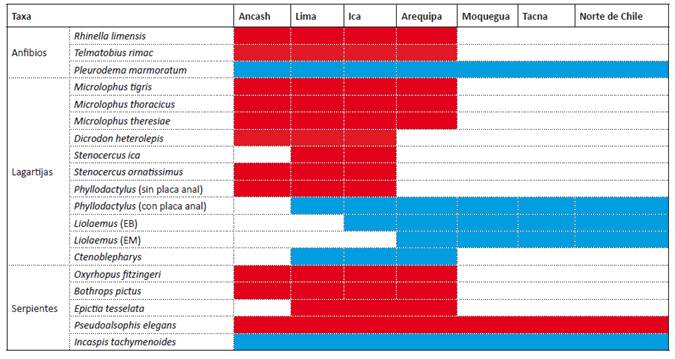
Patrones de distribución de la herpetofauna en el desierto costero del Pacífico y la vertiente occidental al sur de Perú. Varios linajes de anfibios y reptiles presentan límites de distribución en el desierto costero y en la vertiente occidental de los Andes al sur de Perú, y son reemplazados por otra comunidad de herpetofauna (Tabla 2). En el caso de los anfibios, Rhinella limensis termina su distribución sur en el departamento de Arequipa en las altitudes medias y bajas, y es reemplazado por R. spinulosa en las partes de altitud media y alta hasta Chile (Correa et al. 2010, Urra 2013, Pereyra et al. 2021). Cabe mencionar que R. limensis se encuentra en el grupo R. spinulosa, pero se encuentra más relacionado con R. vellardi formando un grupo monofilético hermano de un clado constituido por R. spinulosa y otras especies (Pereyra et al. 2021). De la misma manera, T. rimac presenta su límite de distribución sur en el departamento de Ica y es reemplazado en la vertiente occidental al sur de Perú y Chile por especies del grupo marmoratus como T. arequipensis y T. peruvianus (Sáez et al. 2014, Victoriano et al. 2015, Fibla et al. 2017, Barrionuevo 2017).
Por otro lado, las lagartijas de los géneros Dicrodon (Teiidae) y Stenocercus (Tropiduridae) terminan su distribución sur en el departamento de Ica, no encontrándose especies de estos géneros más al sur o en el norte de Chile (Mendoza et al. 2021, Ruiz de Gamboa 2020). Aguilar et al. (2013) señaló que las especies de Liolaemus (Liolaemidae) que habitan las partes más altas de los Andes tienen su límite de distribución norte en los departamentos de Ancash y Huánuco en el centro de Perú, mientras que los que habitan altitudes medias y bajas de la vertiente occidental y el desierto costero presentan su límite de distribución norte en los departamentos de Arequipa e Ica respectivamente. Sin embargo, Ctenoblepharys adpersa, que pertenece a la misma familia, (Liolaemidae) se distribuye solo desde el norte de Lima hasta el norte de Arequipa en el desierto costero (Ormeño et al. 2021). De igual manera, especies del género Phyllodactylus (P. angustidigitus y P. gerrophygus) que presentan una placa anal terminan su distribución norte al sur del departamento de Lima y son completamente reemplazados, más al norte, por otras especies de Phyllodactylus que carecen de este atributo fenotípico (Aguilar et al. 2007). Las especies de Microlophus del grupo peruvianus que habitan la costa y vertiente occidental como M. thereisae, M. thoracicus y M. tigris tienen su límite de distribución sur en el departamento de Ica y Arequipa, siendo reemplazados por otras especies del grupo peruvianus desde el sur de Perú hasta Chile (Zeballos et al. 2002). Aunque Benavides et al. (2007) identificó individuos como M. tigris al sur de Perú, estos serían parte del complejo M. peruvianus (CA observación personal).
En el caso de las serpientes, no se encuentran representantes de las familias Viperidae y Leptotyphlopidae mas al sur de Perú o en Chile, siendo el extremo de su distribución sur los departamentos de Ica y Arequipa (Carrillo & Icochea 1995, Ruiz de Gamboa 2020). De igual manera, Oxyrophus fitzingeri (Colubridae) tendría su distribución sur en el departamento de Arequipa y no existen representantes de este género en Chile (Arapa 2018, Mamani-Cabana 2020, Ruiz de Gamboa 2020).
Una posible barrera para las especies de anfibios y reptiles cuya distribución sur termina en el departamento de Ica y norte de Arequipa serían las zonas áridas (precipitación menor a 50 mm/año) e hiperáridas (precipitación menor a 5 mm/año) del desierto peruano y de Atacama (Luebert 2021). Ambos desiertos estarían presentes desde el oligoceno tardío (33 millones de años AP) para las zonas áridas, pero se habría intensificado durante el Mioceno hace ocho millones de años o más para las zonas hiperáridas (Guerrero et al. 2013, Luebert 2021). La herpetofauna cuya distribución sur termina en el departamento de Ica y Arequipa sería reemplazada en algunos casos por otras que toleran estos ambientes hiperáridos y cuya distribución norte termina en el centro-sur de la vertiente occidental y desierto costero de Perú. Sin embargo, la extensión geográfica de esta zona hiperárida varió durante las fluctuaciones climáticas del cuaternario con episodios de mayor humedad (Betancourt 2000, Böhnert 2020) y pueden haber contribuido a la distribución actual de los anfibios y reptiles. Un ejemplo de lo mencionado lo ofrecen las especies de Liolaemus del grupo montanus que habitan las zonas áridas e hiperáridas de Perú y Chile. Las especies de Liolaemus de este grupo son principalmente andinas, pero las que habitan las zonas desérticas del desierto costero y vertiente occidental convergen en mayor o menor medida fenotípicamente con Ctenoblepharys adspersa también habitante del desierto costero (Aguilar et al. 2018). Sin embargo, C. adspersa sería un linaje antiguo (no menor a los 55 millones de años, Aguilar et al. 2018) y con una distribución restringida que no se traslapa con la distribución de los Liolaemus convergentes. Estos Liolaemus serían linajes más jóvenes (no mayor a los 8 millones de años, Aguilar et al. 2018) y representarían una invasión posterior e independiente del desierto costero y la vertiente occidental más árida (Guerrero et al. 2013, Aguilar et al. 2018). Este patrón de distribución parece concordante con una influencia temprana de las zonas áridas e hiperáridas desde el oligoceno para C. adspersa y una tardía para las especies convergentes de Liolaemus cuando regresaron las condiciones hiperáridas en el cuaternario.
Tabla 2 Distribución de anfibios y reptiles presentes en el departamento de Ica que terminan su distribución en la vertiente occidental y desierto costero en Perú. “X” los taxones con distribución principalmente norte (solo se incluye hasta el departamento de Ancash) y terminan su distribución en los departamentos de Ica y Arequipa. “O” los taxa con distribución principalmente sur (solo los que ocurren hasta Chile) y terminan su distribución en los departamentos de Ica y Lima. EB = elevaciones bajas, EA = elevaciones altas.