











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
La técnica de diafanización tiene como principal finalidad transparentar los tejidos blandos de un organismo, lo que permite observar los elementos óseos (Romero-Oliva y González-Rodríguez, 2019), se realiza a través de la digestión alcalina con hidróxidos y colorante rojo de alizarina para la tinción, el cual se fija a los fosfatos de calcio que se encuentran en el tejido óseo (Rejala et al., 2019), dicha técnica presenta grandes ventajas en la determinación de especies y es de utilidad para entender el desarrollo osteológico (Moreno-Guerra et al., 2019; Koch et al., 2024).
Los poecílidos son una familia de peces cosmo-politas de agua salobre y dulce en su mayoría, se encuentran en latitudes templadas y tropicales de África, India Occidental y América (Meffe & Snelson 1989; Ghedotti, 2000). La familia está representada por aproximadamente 270 especies válidas (Fricke et al., 2020), cuya distribución en el continente americano comprende desde Estados Unidos hasta el Noreste de Argentina (Miller et al., 2005), pero su mayor diversidad ocurre en Centroamérica, México y las islas del Caribe (Rosen y Bailey, 1963).
Todas las especies excepto una de esta familia son vivíparas, (Furness et al., 2019; Safian et al., 2023), ya que las hembras presentan fecundación interna por medio del gonopodio del macho, portando a los embriones en la gónada hasta su nacimiento (Torres-Martínez et al., 2023); las hembras son altamente fértiles, capaces de producir camadas sucesivas, esto debido a su capacidad de retener el esperma y presentar superfetación (Miller et al., 2005; de León-Gonzales, 2022), teniendo la capacidad de gestar simultáneamente dos o más grupos de embriones en diferentes etapas de desarrollo (García-Cabello, 2022).
En embriones de peces el desarrollo osteológico es un proceso detallado que se inicia con la formación de cartílago y su posterior osificación (Rodríguez-Ibarra et al., 2017); considerando los trabajos osteológicos con poecílidos son escasos y se han centrado generalmente en áreas específicas del esqueleto, por ejemplo, Mookerjee & Mazumdar (1940) con la descripción del desarrollo de la columna vertebral de Lebistes reticulatus.
Strauss (1990), examinó la variación de osificación en el cráneo de cinco especies de poecílidos; otro trabajo por Ruiz (1998), quien realizó la descripción osteológica del neurocráneo de Poecilia vivípara; por otra parte, Weisel (1967), comparó el desarrollo osteológico de Catostomus machhrocheilus y Poecilia reticulata, mencionando que las primeras piezas óseas en calcificarse en ambas especies son los huesos móviles, asociados con la respiración y alimentación. También la osteología, se ha utilizado para examinar relaciones sistemáticas Rauchenberger (1989), utilizó la osteología descriptiva de Gambusia hispaniolae y determinar interrelaciones entre los subgéneros del género Gambusia.
Debido a lo antes mencionado, el presente trabajo se centró en la aplicación de la técnica de diafanización en embriones de P. sphenops con la intención de obtener una identificación clara de las primeras piezas óseas que comienzan a formarse durante su ontogenia, de tal manera se plantearon los siguientes objetivos: Aplicar la técnica de diafanización y que nos ayude a determinar las primeras piezas óseas que se forman durante su desarrollo; así mismo, se separaron los embriones en relación al tamaño y se identificaron las piezas óseas presentes para determinar las diferentes etapas de desarrollo de cada embrión y de esta manera se espera encontrar que las primeras estructuras óseas en aparecer serían aquellas relacionadas con la alimentación, respiración y natación como sucede en otras especies de esta familia.
METODOLOGÍA
Material biológico
Los embriones fueron obtenidos de 12 hembras depositadas en la Colección Ictiológica del Centro de Investigaciones Biológicas “Dr. Edmundo Díaz Pardo” de la Universidad Autónoma del Estado de Morelos (CIB-UAEM)SEMARNAT-(MOR-CC-243-2011), y catalogados con los números 200 y 403; procedentes de dos localidades del estado de Morelos; “Las Huertas” y “Amacuzac” (Tabla 1).
Diafanización
Se extrajeron las gónadas de las hembras grávidas y se colocaron dentro de cajas Petri, los embriones fueron sometidos a la técnica de diafanización propuesta por Hollister (1934) y Davis & Gore (1936) y modificada por Paredes-Lira (1998). La técnica consiste en recambios de hidróxido de potasio diluido con agua corriente al 4% para macerar y transparentar el tejido, para la tinción de los elementos óseos se utilizó rojo de alizarina, dando una tonalidad a las estructuras óseas de color morado, después de varios pasos se conservaron en glicerina industrial.
Selección y medición de embriones
Una vez diafanizados los embriones, fueron observados a través de un microscopio compuesto marca NIKON modelo 541795 a 5X y fotografiados con una cámara CANON EOS Rebel T3i, con adap-tador 23,2 mm, seleccionando aquellos embriones con presencia de material óseo (Figura 1).
Tabla 1 Datos de los sitios de recolección de las hembras de P. sphenops
| Municipio y Localidad | Fecha de Colecta | Coordenadas | Número de ejemplares (H) |
|---|---|---|---|
| Tlaquiltenango - Las Huertas | Sept/1994 | 18°28´ N y 99°08´ O | 6 |
| Amacuzac - Amacuzac | Nov/1993 | 18°35´ N y 99°22´ O | 4 |
| Sept/1994 | 2 |

Figura 1. A) Selección de embriones. B) Medición de embriones en posición fetal. Estos últimos a su vez fueron separados tomando como referencia la Longitud total (LT), poste-riormente se colocados en cajas Petri con hojas milimétricas por debajo, ya que los embriones presentaban una posición fetal, impidiendo el uso de una reglilla micrométrica para la medición (Figura 1) y determinar de esta manera la LT con respecto al número de piezas presentes durante su desarrollo embrionario.
Identificación de piezas óseas
Las piezas óseas encontradas fueron identificadas de acuerdo con los esquemas de Weisel 1967 (Tabla 2). Para la descripción de los ejemplares se anotó el número de elementos óseos presentes, ordenando los datos en base al número de piezas (del menor al mayor), además del acomodo que guardan en el cráneo, tronco y cola, estableciendo el grado de osificación en cada una de ellas y anotando las piezas pares y el lado simétrico donde se localizaron.
Análisis estadístico
Se realizó un análisis de correlación (Coeficiente de Pearson, 0,5) entre la LT y el número de piezas óseas presentes en los embriones; el cálculo de correlación se realizó en una hoja de cálculo de Excel utilizando la fórmula de correlación (=COEF.DE.CORREL(MATRIZ1, MATRIZ2)) ya incluida dentro del software, utilizando como matriz 1 la LT y como matriz 2 el número de piezas óseas presentes.
Tabla 2 Lista de elementos del esqueleto de Poecilia sphenops con abreviaturas adaptadas al español de Weisel 1967
| Abreviatura | Pieza ósea | Abreviatura | Pieza ósea | |
|---|---|---|---|---|
| PM | Premaxilar | F | Frontal | |
| MX | Maxilar | CB | Ceratobranquial | |
| D | Dentario | ATS | Autoesfenótico | |
| A | Articular | PTS | Pteroesfenoide | |
| C | Cuadrado | LAP | Lapilus | |
| IOP | Interopérculo | SAG | Sagitas | |
| POP | Preopérculo | AST | Asteriscus | |
| OP | Opérculo | RC | Radios Caudales | |
| SOP | Subopérculo | HP | Hipural | |
| SP | Simpléctico | UR | Urostilo | |
| HH | Hipohial | EP | Epural | |
| CH | Ceratohial | V | Vértebras | |
| BS | Branquiostegos | E | Escamas | |
| CL | Cleitro | EL, PF | Etmoides lateral prefrontal | |
| PRO | Proótico | BH | Basihial | |
| BO | Basioccipital | P | Palatino | |
| EO | Exoccipital | PV | Prevomer | |
| PS | Paraesfenoides | IF | Infrafaríngeos | |
| HM | Hiomandibular | SF | Suprafaríngeos |
RESULTADOS Y DISCUSIÓN
Análisis de las hembras superfetantes
De las 12 hembras presentes de las dos localidades, seis de cada localidad respectivamente, se obtuvieron un total 564 ovocitos de los cuales se descartaron aquellos inviables es decir que no presentan embriones aun formados, quedando 110 embriones presentes en tres hembras (Tabla 3), también se descartaron aquellos embriones que aún no presentaban piezas óseas, quedando un total de 66 embriones de 3,8 mm a 9,4 mm de LT.
Tabla 3 Antecedentes de las hembras que presentaron embriones con piezas óseas
| Hembra (H) | Localidad/ fecha de colecta | Tamaño LT hembras (mm) | Peso (g) | Estadio de la gónada | Peso de la gónada (g) | # Embriones | # Embriones con piezas óseas | LT embriones (mm) |
| 01 | Amacuzac Nov/1993 | 65,28 | 6,5 | IV | 0,3 | 51 | 25 | 3,8 - 6 |
| 12 | Amacuzac Sep/1994 | 39,48 | 1,9 | V | 0,4 | 35 | 33 | 5 - 8,1 |
| 02 | Las Huertas Sep/1994 | 45,82 | 2,7 | IV | 0,4 | 24 | 8 | 5,5 - 9,4 |
La superfetación fue un fenómeno que se presentó en la hembra de menor LT, quedando tres grupos de embriones simultáneos (H12). Por su parte, la hembra de mayor LT presentó el número de embriones más alto por camada, sin embargo, sólo presentó dos grupos de embriones simultáneos y en general fueron los más pequeños e incompletos (H01) (Tabla 3). Zuñiga-Vega (2017) reportó que hembras de tamaño pequeño y mediano de Poeciliopsis baenschi presentan mayor grado de superfetación, mientras que las hembras grandes producen pocas o ninguna cría simultánea. Urbano-Amilpa (2006) menciona que la longitud total de la hembra por sí sola determina el tamaño de la camada y un aumento en longitud de la hembra se asocia con grandes camadas y aumento en el tamaño (LT) de las crías.
Las hembras presentaron diferencias y similitudes en el desarrollo de maduración de las gónadas, presentándose en dos estadios (IV - V). Las hembras de mayor LT (H01 - H02) presentaron sus gónadas en el estadio IV a diferencia de la hembra de menor LT (H12) la cual presentó su gónada en el estadio V. Si bien el peso de la gónada de la H02 y H16 es el mismo, su estadio de maduración es distinto, además que el peso entre ambas hembras varía significativamente (Tabla 3). Urbano-Amilpa (2006) mencionó la relación entre el peso de la hembra y el peso de la gónada en tres especies del género Xiphophorus observando que conforme aumenta el primero, el segundo también se incrementa. Young (2019) reportó que en hembras de Cnesterodon decemmaculatus el estadio IV de la gónada está relacionado a hembras juveniles mientras que el estadio V a hembras adultas.
Secuencia de osificación
Gracias a la técnica de diafanización, la osificación de las piezas óseas se pudo observar a partir del estadio 5 del desarrollo embrionario en distintas regiones del esqueleto en P. sphenops y Sancho-Martínez (2018), menciona al respecto que el desarrollo embrionario la familia Poeciliidae presenta 10 estadios esenciales para completar su formación anatómica-fisiológica, para este estudio P. shenops, solo presentó ocho estadios embrio-narios, pero fueron eliminados los tres primeros ya que no presentaron piezas en desarrollo. Urbano-Amilpa (2006), menciona que en el caso del género Xiphophorus los embriones en el estadio 5 se encuentran aún en etapa temprana de desarrollo y no se observa material óseo en la región cefálica y la cavidad ocular.
Los resultados obtenidos del número de embriones y piezas óseas observadas después de la diafanización se describen en Tablas 4 y 5. El total de los embriones se separaron en dos grupos; embriones iniciando su desarrollo óseo (2 - 9 Piezas óseas) (Tabla 4) y embriones terminando su desarrollo óseo (27 - 37 Piezas óseas) (Tabla 5).
Los embriones más pequeños en el que se pudo observar el inicio de la osificación midieron 3,8mm de LT; por otra parte, Strauss (1990) en su trabajo señala que algunos poecílidos comienzan su osificación durante el desarrollo embrionario a los 4 mm de LT y continua durante el estadio juvenil a diferencia de P. sphenops y Weisel (1967) observó que en Poecilia reticulata a partir de los 4,2 mm de LT comienza la osificación.
En otras especies de familias distintas dentro del rango de los 3,8 mm de LT se ha observado la osificación de diversas piezas óseas, como Álvarez-Pliego (2004) con Diapterus peruvianus, determi-naron la formación de los primeros radios de la aleta caudal. Rodríguez-Ibarra et al., (2014) deter-mina con Lutjanus guttatus el inició de la formación del esqueleto axial con los arcos neurales y hemales en embriones de 3,8mm de LT.
En el caso de P. sphenops, los embriones con presencia de 2 a 3 piezas óseas tuvieron una LT de 5 a 7,4 mm (Tabla 4). El Emb.1 fue el que presentó menor número de piezas óseas osificadas; los opérculos (OP) y el cleitro (CL). Los embriones con presencia de 3 piezas óseas mostraron estructuras óseas distintas entre sí; el Emb.2, si presento un opérculo, pero no el cleitro, en su lugar presento los branquiostegos (BS) y el paraesfenoides (PS) ambos comenzando su desarrollo de osificación, el tercer Emb.3 no tuvo nada en común con los pasados y presentó, primeramente, las vértebras que solo se obser-vaban como un cordón seguido de los otolitos sagitas (SAG) y lapilus (LAP) que presentan poco desarrollo. Weisel (1967) señala a los cleitros piezas pares importantes y necesaria para la respiración, los músculos que están unidos a esta pieza ayudan a mover los arcos branquiales.
Los embriones con presencia de 4 a 5 piezas óseas tuvieron una LT de 3,8 a 8 mm (Tabla 4). A partir de los embriones que presentan 4 piezas óseas se forma el dentario (D), pieza presente, pero poco osificada para este grupo de embriones; así mismo, comienza la osificación de los radios caudales (RC) con la formación de 2 radios hasta los 6 radios en embriones con 5 piezas óseas. Los embriones con 5 piezas óseas en adelante ya presentan los opérculos y el cleitro sin excepción.
Tabla 4 Resultados del desarrollo osteológico por camada de Poecilia sphenops embriones con pocas piezas óseas
| # Embrión (Emb.) | Camada | Localidad | # Hembra | # Piezas óseas | Longitud Total (mm) |
| 1 | (4) - 09 - 1994 | Amacuzac | 12 | 2 | 7,4 |
| 2 | (4) - 11- 1993 | Amacuzac | 01 | 3 | 5 |
| 3 | (4) - 09 - 1994 | Amacuzac | 12 | 3 | 6,9 |
| 4 | (4) - 11- 1993 | Amacuzac | 01 | 4 | 4 |
| 5 | (4) - 11- 1993 | Amacuzac | 01 | 4 | 4,6 |
| 6 | (4) - 11- 1993 | Amacuzac | 01 | 4 | 5,3 |
| 7 | (4) - 09 - 1994 | Amacuzac | 12 | 4 | 6,8 |
| 8 | (4) - 09 - 1994 | Amacuzac | 12 | 4 | 7 |
| 9 | (4) - 09 - 1994 | Amacuzac | 12 | 4 | 7 |
| 10 | (4) - 09 - 1994 | Amacuzac | 12 | 4 | 8 |
| 11 | (4) - 11- 1993 | Amacuzac | 01 | 5 | 3,8 |
| 12 | (4) - 11- 1993 | Amacuzac | 01 | 5 | 3,8 |
| 13 | (4) - 11- 1993 | Amacuzac | 01 | 5 | 4 |
| 14 | (4) - 09 - 1994 | Amacuzac | 12 | 5 | 5 |
| 15 | (4) - 11- 1993 | Amacuzac | 01 | 5 | 5,1 |
| 16 | (4) - 09 - 1994 | Amacuzac | 12 | 5 | 6,1 |
| 17 | (4) - 09 - 1994 | Amacuzac | 12 | 5 | 6,5 |
| 18 | (4) - 09 - 1994 | Amacuzac | 12 | 5 | 7,7 |
| 19 | (4) - 11- 1993 | Amacuzac | 01 | 6 | 5 |
| 20 | (4) - 11- 1993 | Amacuzac | 01 | 6 | 5,4 |
| 21 | (4) - 09 - 1994 | Amacuzac | 12 | 6 | 6,8 |
| 22 | (4) - 09 - 1994 | Amacuzac | 12 | 6 | 7,1 |
| 23 | (4) - 11- 1993 | Amacuzac | 01 | 7 | 4,9 |
| 24 | (4) - 11- 1993 | Amacuzac | 01 | 7 | 5 |
| 25 | (4) - 11- 1993 | Amacuzac | 01 | 7 | 5,3 |
| 26 | (4) - 09 - 1994 | Amacuzac | 12 | 7 | 7,2 |
| 27 | (4) - 09 - 1994 | Amacuzac | 12 | 7 | 7,3 |
| 28 | (4) - 09 - 1994 | Amacuzac | 12 | 7 | 7,8 |
| 29 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 4,5 |
| 30 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 4 |
| 31 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 4,2 |
| 32 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 4,5 |
| 33 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 4,5 |
| 34 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 4,9 |
| 35 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 5 |
| 36 | (4) - 11- 1993 | Amacuzac | 01 | 8 | 5,2 |
| 37 | (4) - 09 - 1994 | Amacuzac | 12 | 8 | 6,4 |
| 38 | (4) - 09 - 1994 | Amacuzac | 12 | 8 | 6,8 |
| 39 | (4) - 09 - 1994 | Amacuzac | 12 | 8 | 7,4 |
| 40 | (4) - 09 - 1994 | Amacuzac | 12 | 8 | 7,6 |
| 41 | (4) - 09 - 1994 | Amacuzac | 12 | 8 | 7,7 |
| 42 | (4) - 09 - 1994 | Amacuzac | 12 | 8 | 8,1 |
| 43 | (4) - 11- 1993 | Amacuzac | 01 | 9 | 4 |
| 44 | (4) - 11- 1993 | Amacuzac | 01 | 9 | 4 |
| 45 | (4) - 11- 1993 | Amacuzac | 01 | 9 | 4,8 |
| 46 | (4) - 11- 1993 | Amacuzac | 01 | 9 | 6 |
| 47 | (4) - 09 - 1994 | Amacuzac | 12 | 9 | 7 |
| 48 | (4) - 09 - 1994 | Amacuzac | 12 | 9 | 7,1 |
| 49 | (4) - 09 - 1994 | Amacuzac | 12 | 9 | 7,2 |
| 50 | (4) - 09 - 1994 | Amacuzac | 12 | 9 | 6 |
Los otolitos, por su parte, presentan una forma de cazuela y dentro de estos se forman varias piezas dispersas que no se pueden identificar, ya que solo se ven puntos oscuros, y es así en la mayoría de los embriones con presencia de otolitos (Figura 2). A diferencia de otros estudios, donde no se describe por talla, el desarrollo de las vértebras (V) es intermitente para P. shenops, debido a que algunos embriones siguen presentándose como un cordón y otros ya las presentan formadas variando entre 3 - 14 vértebras.
Los embriones con presencia de 6 a 7 piezas óseas tuvieron una LT de 4,5 a 7,8mm en P. shenops (Tabla 4), los Emb.19 al Emb.22 con 6 piezas óseas en general presentaron el paraesfenoides y los branquiostegos aún en desarrollo y no están presentes en todos los embriones, las vértebras dejan de verse como un cordón, sin embargo, aún se encuentran poco desarrolladas. Es en los Emb.23 al Emb.28 con 7 piezas óseas que se observa la presencia de los branquiostegos sin excepción.
En este segundo grupo de embriones las vértebras comienzan a formar anillos en el cordón vertebral.
La osificación de la columna vertebral puede iniciar a partir de los 3,8mm de LT con la formación del cordón vertebral y la posterior diferenciación de las tres primeras vértebras, sin embargo, también se encontraron embriones de mayor a 3,8mm de LT sin desarrollo en la columna.
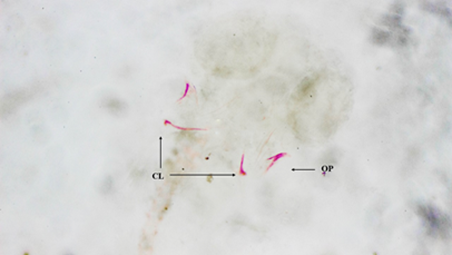
Figura 2 Fotografía vista ventral. Embrión de Poecilia sphenops. Otolitos en forma de cazuela con mucha pigmentación.
Esto difiere a lo observado por Weisel (1967) en P. reticulata donde la osificación de las vértebras inicia a partir de los 5 mm de LT. Otros autores han descrito ampliamente el desarrollo de la columna vertebral, en distintas familias, considerando a Hernández, et al. (2016) con Prochilodus lineatus mencionan que a partir de los 7,8 mm de LT comienza una ligera osificación de los primeros centros vertebrales. Por su parte, Rodriguez-Ibarra, et al., (2014) con Lutjanus guttatus, describen el desarrollo de la columna a partir de los 3,8mm de LT con la presencia de estructuras cartilaginosas. Nuevamente, Rodriguez-Ibarra, et al. (2017) con Amphiprion ocellaris mencionan que a los 4 mm de LT ya se presentan la mayoría de sus elementos vertebrales, pero aún en etapa de cartílago.
Los embriones con presencia de 8 a 9 piezas óseas tuvieron una LT de 4 a 8,1 mm (Tabla 4) para P. shenops, a partir de los embriones con 8 piezas óseas ya se presentan los branquiostegos y el paraesfenoides. El número de vértebras aumenta (21 vértebras) a partir de los embriones que presentan 9 piezas óseas (Emb.42 - Emb.50); sin embargo, se encuentran poco desarrolladas y sin cerrar. De igual manera, el número de radios caudales (RC) aumenta (9 RC), las siguientes piezas óseas en aparecer y formarse son los frontales (F) y el basioccipital (BO).
Generalmente los primeros huesos en osificarse son los opérculos, el cleitro, los branquiostegos, las vértebras, radios caudales y el paraesfenoides, las siguientes piezas pueden variar entre los otolitos sagitas, otolitos lapilus y el dentario. Weisel (1967) y Strauss (1990), mencionan que los primeros huesos en osificarse en los poecílidos son los maxi-lares, los dentarios, los opérculos y el cleitro seguidos de estructuras asociadas con movimien-tos respiratorios complejos, incluidos los prema-xilares, articulares, cuadrantes y simplécticos.
Los embriones con presencia de 27 a 30 piezas óseas tuvieron una LT de 6,6 a 7mm (Tabla 5). Los Emb.51 a Emb.54 no presentaron otolitos o solamente presentaron uno siendo el lapilus o sagitas, pero no ambos al mismo tiempo. Las vértebras dejan de presentarse como un cordón, y ya se presentan 27 - 28 vértebras donde la 1er y 8va vértebra están más desarrolladas que sus espinas, se presentan de 24 a 30 radios caudales. Los Emb.51 al Emb.54 presentan bien desarrollados; el maxilar (MX), preopérculo (POP), subopérculo (SOB) y simpléctico (SP), seguidos del ceratohial (CH), cuadrado (C), pterosfenoide (PTS), el etmoi-des lateral prefrontal (EL, PF), el ceratobranquial (CB), autosfenotico (ATS), proótico (PRO), intero-pérculo (IOP), hipohial (HH) e hiomandibular (HM) por lo cual es difícil determinar una secuencia de osificación para estas piezas. Basándonos en el grado de osificación las siguientes piezas en aparecer son el articular (A), basihial (BH) y exoccipital (EO) seguidos por el hipural (HP) que presenta poco desarrollado. El prevomer (PV) y el palatino (P) son poco visibles o están comenzando a desarrollarse. Se observó que a partir de los embriones que presentan 29 piezas óseas (Emb.52 - 53) en adelante, en ocasiones el premaxilar (PM), infrafaríngeos (IF) y los suprafaríngeos (SF) ya presenta dientes (Figura 3).
Por otra parte, trabajos previos podrían darnos una idea de la secuencia de aparición y osificación de algunas de las piezas dentro de este grupo de huesos, por ejemplo, Weisel (1967) observó que en embriones de 5 mm LT de Poecilia reticulata los maxilares, premaxilares y subopérculos son los primeros en desarrollarse y osificarse.
Tabla 5 Resultados del desarrollo osteológico por camada de Poecilia sphenops embriones con la mayoría de sus piezas óseas
| # Embrión (Emb.) | Camada | Localidad | # Hembra | # Piezas óseas | Longitud Total (mm) |
| 51 | (9) - 09 - 1994 | Las Huertas | 02 | 27 | 7 |
| 52 | (4) - 09 - 1994 | Amacuzac | 16 | 29 | 6,4 |
| 53 | (4) - 09 - 1994 | Amacuzac | 16 | 29 | 6 |
| 54 | (9) - 09 - 1994 | Las Huertas | 02 | 30 | 6,6 |
| 55 | (9) - 09 - 1994 | Las Huertas | 02 | 31 | 7,8 |
| 56 | (9) - 09 - 1994 | Las Huertas | 02 | 31 | 9,4 |
| 57 | (9) - 09 - 1994 | Las Huertas | 02 | 32 | 7 |
| 58 | (9) - 09 - 1994 | Las Huertas | 02 | 32 | 8,1 |
| 59 | (4) - 09 - 1994 | Amacuzac | 16 | 32 | 6,5 |
| 60 | (4) - 09 - 1994 | Amacuzac | 16 | 32 | 6,1 |
| 61 | (9) - 09 - 1994 | Las Huertas | 02 | 33 | 6,5 |
| 62 | (4) - 09 - 1994 | Amacuzac | 16 | 33 | 6,4 |
| 63 | (4) - 09 - 1994 | Amacuzac | 16 | 33 | 6,2 |
| 64 | (9) - 09 - 1994 | Las Huertas | 02 | 35 | 6,8 |
| 65 | (4) - 09 - 1994 | Amacuzac | 16 | 35 | 6,4 |
| 66 | (4) - 09 - 1994 | Amacuzac | 16 | 37 | 6,4 |
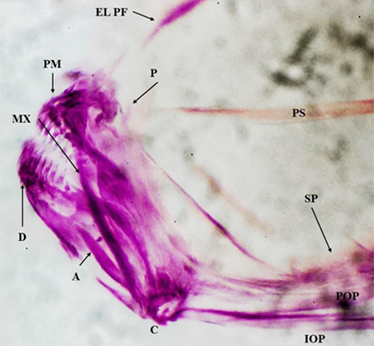
Figura 3 Fotografía vista lateral. Embrión de P. sphenops región mandibular. Presencia de dientes en el D; Dentario y PM; Premaxilar. Se puede observar que la estructura de la mandíbula termina con la presencia del A; Articular, C; Cuadrado, MX; Maxilar, P; Palatino, SP; Simpléctico. También ya se presenta el IOP; Interopérculo y POP; Preopérculo.
Los embriones con presencia de 31 a 33 piezas óseas tuvieron una LT de 6,5 a 9,4 mm (Tabla 5). Los Emb.55 a Emb.63 ninguno en este grupo de embriones presentó otolitos. Las escamas, por su parte, tienen distinto estado de desarrollo en este grupo de embriones. También se observó que a partir de los embriones que presentan 33 piezas óseas en adelante, todos ya presentan el urostilo (UR); sin embargo, en algunos embriones aún no se encuentra dividido.
En cuanto a los otolitos no está muy claro cuáles son los primeros en aparecer si los sagitas o lapilus, si bien los otolitos sagitas tienen más presencia que los otolitos lapilus en los embriones, también son estos los que en más ocasiones solo se presenta uno del par, por lo que podría considerarse que en general se forman y osifican al mismo tiempo. Sin embargo, en base a la literatura y de acuerdo con Rojo (2017) en la mayoría de las especies, el otolito sagitas es el primero en aparecer.
Los embriones con presencia de 35 a 37 piezas óseas tuvieron una LT de 6,4 a 6,8 mm (Tabla 5) (Figura 4). A partir de los Emb.64 y Emb.65 nuevamente observan los otolitos sagitas y lapilus también ya están presentes los otolitos asteriscus (AST) estando poco visibles o bien desarrollados. Se presenta un máximo de 30 vértebras bien. En estos embriones (Emb.64 - Emb.66) es poca la presencia de escamas. Sin embargo, ninguno de los embriones presentó el epural (EP).
Finalmente, ningún embrión presentó epural por lo que podrían considerarse dos posibilidades, la primera es que dicha pieza es la última en aparecer y osificarse antes de que el pez nazca, siendo de esta manera que las crías de Poecilia sphenops nacen con su esqueleto completamente formado, o como segunda posibilidad las crías aun después de su nacimiento continúan con la formación de dicha pieza, como lo descrito por Weisel (1967) en embriones de Poecilia reticulata donde las crías aun después de su nacimiento continúan con la formación de unas pocas piezas óseas como los proóticos y el basihial.
Correlación LT - Número de piezas óseas
Como resultado del cálculo de correlación se obtuvo un valor de 0,311045 lo que resulta en una correlación moderada entre LT y el número de piezas óseas. Esto ya se venía observando durante la obtención de datos ya que se encontró que embriones de tallas mayores a la media (6,09mm) en muchas ocasiones presentaban menor número de piezas óseas que embriones de tallas menores a la media y viceversa. Weisel (1967) en Poecilia reticulata, menciona que conforme el organismo crece el número de piezas óseas aumenta.
CONCLUSIONES
El tamaño de la hembra está relacionado con el número de embriones por camada, es decir entre más grande es una hembra mayor es el número de embriones que presenta. El grado de superfetación no depende del estado de maduración de la gónada ni del tamaño de la hembra.
La superfetación es un fenómeno presente en hembras de tamaño pequeño, sin embargo, dicho fenómeno permite a las hembras pequeñas presentar igual o mayor número de embriones por camada, que las hembras de mayor tamaño; siendo así que los embriones simultáneos se dividen en tres grupos; embriones sin presencia de piezas óseas, embriones con presencia de pocas piezas (2-9 piezas) y embriones con presencia de mayoría de piezas (27-37 piezas) presentes.
El cálculo de correlación de Pearson que corres-ponde entre el tamaño del embrión y el número de piezas óseas resultó de en un valor de 0.31 considerándose como una correlación moderada, sin embargo, debido a la ausencia de embriones con un número de piezas óseas que van de 9 a 27 este resultado podría cambiar ya que no aparecieron embriones con intermedios entre 9 a 27 piezas para determinar un orden de aparición y osificación del premaxilar, maxilar, preopérculo, subopérculo y simpléctico que corresponden a las estructuras de la alimentación.
Si queda claro que ni el tamaño de la hembra, ni el tamaño de la gónada determina la osificación inicial, pero sí cuando los embriones se encuentran en el estadio 5 del desarrollo embrionario.
En general P. sphenops presenta similitudes en la secuencia de osificación de sus primeras piezas óseas con otros poecílidos, sin embargo, se caracteriza por no seguir una secuencia estricta en el desarrollo de todos sus embriones en una sola gónada.
El tamaño mayor de una muestra de hembras grávidas permitiría un informe más preciso de la secuencia, tiempos de osificación y grado de superfetación.

