










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.12 no.3 Lima Oct./dic 2005
Eduardo A. Sanabria1, Lorena B. Quiroga y Juan C. Acosta
1 Departamento de Biología e Instituto y Museo de Ciencias Naturales, Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de San Juan, Argentina.
Presentado: 19/10/2005
Aceptado: 21/11/2005
Resumen
El presente trabajo estudia la dieta de una población de Leptodactylus ocellatus de los Humedales de Zonda, San Juan, al oeste de Argentina. Se analizaron 35 estómagos, las presas fueron clasificadas y los individuos se midieron en ancho y largo para calcular su volumen con la fórmula de la esfera elipsoide; además, se estimó la frecuencia de ocurrencia y abundancia. Se calcularon índices de diversidad, de similitud, amplitud de nicho e importancia relativa (IRI). Los presas mas importantes fueron: Hymenoptera, Anura, Coleoptera. Las presas con mayor porcentaje de presencia fueron los Hymenoptera Formicidae que representaron el 22,4%. No se encontró una relación significativa entre el ancho de la boca del depredador y el tamaño medio de la presa. La dieta observada en ambos sexos resultó similar.
Palabras clave: Dieta, Leptodactylus ocellatus, San Juan, Argentina.
Abstract
This work study the diet of Leptodactylus ocellatus from the swamp of Zonda, San Juan, in western Argentina. We analyze 41 stomaches, preys were classified and indiviudals were measured (wide and long), to calculate their volume with the sphere ellipsoid, furthermore, the frequency and the abundance was also considered. Indexes of diversity, of similarity, niche width and relative importance (IRI), were calculated. The more important preys were: Hymenoptera, Anura, Coleoptera. The preys with more percentage of presence were the Hymenoptera Formicidae (22,4%). Relationship among the predator's width mouth and the average size of the prey was not significant. Diet in both sexes were similar.
Keyworks: Diet, Leptodactylus ocellatus, San Juan, Argentina.
Los estudios de dieta son importantes por que permiten determinar el uso de recursos en el marco de la teoría de la segregación de nichos, nos proporcionan información sobre las historias de vida de las especies y nos brinda elementos para tomar decisiones de conservación y manejo.
La selección de las presas está influenciada tanto por el tipo de hábitat, como por la estacionalidad, y de acuerdo a esto, cada ítem alimentario pueden variar en cantidad y calidad (Duellman y Trueb, 1986).
El género Leptodactylus cuenta con, aproximadamente, cincuenta especies y con una gran diversificación, de especies en cuanto a su morfología, etología y ecología (Lajmanovich, 1996). El área de distribución de la especie se extiende desde el sur de Brasil y parte de Paraguay, Uruguay y en Argentina, desde Misiones hasta Buenos Aires, sur de Río Negro, Córdoba, San Luis, Mendoza, San Juan y oeste de Neuquen. El hábitat característico son pantanos y arroyos desde el nivel del mar hasta 1000 a 1200 m, principalmente en pastizales húmedos (Cei, 1980; Gallardo, 1987). Los juveniles de L. ocellatus han sido estudiados por Lajmanovich (1996) y los considera generalistas, con un marcado cambio de los grupos funcionales de la dieta con relación directa al aumento del tamaño corporal.
El objetivo del presente trabajo es analizar la dieta de L. ocellatus en los Bañados de Zonda, un humedal del Monte de San Juan, Argentina.
Área de estudio
El área de estudio está ubicada a 25 Km al Oeste de la ciudad de San Juan, en el Departamento Zonda (31°55´ S, 68°70´ W), Argentina (Fig.1). Son humedales de tipo temporal, alimentados por las aguas subterráneas que provienen del Dique de Ullum, que se encuentra contiguo a éstos. Pertenece a la provincia Fitogeográfica del Monte (Cabrera, 1994) ubicada a una altitud de 800 m, región de clima árido con una temperatura media anual de 17,3 ºC y una máxima media anual de 25,7 ºC y mínima media anual de 10,4 ºC, con las lluvias concentradas en verano con un promedio anual de 84 mm.
La vegetación del lugar es muy rica en especies propias de humedales, entre las más importantes se pueden mencionar a: Cortaderia sp., Tipha dominguensis, Malvella leprosa, Heliotropium sp., Phyla canescens, Melilotus indicus, Gnaphalium sp., Ciperus sp., Prosopis strombulifera, Prosopis sp., Atriplex sp., Larrea spp. (Cabrera, 1994).
Procedimiento
Desde febrero de 2002 hasta octubre de 2002, se realizaron 20 días de colecta. Éstas fueron hechas manualmente durante caminatas diurnas y nocturnas al azar en transectos obteniendo un total de 35 individuos en todo el periodo de estudio. Los individuos fueron fijados con formaldehído al 10% y luego conservados en alcohol al 70%, se determinó el sexo, se midió la longitud hocico cloaca (LHC) y el ancho de la boca a la altura de las comisuras, con un calibrador Vernier de 0,2 mm de precisión.
Se analizaron los estómagos determinándose las presas a nivel de orden y familia. A cada presa se le midió su largo (L) y ancho (A) con el fin de calcular su volumen con la fórmula de una esfera elipsoide (Dunham, 1983).
Para conocer la contribución de cada categoría de alimento a la dieta se calculó el Índice de Importancia Relativa (IRI) propuesto por Pinkas et al., (1971). Para calcular la jerarquía de las presas en la dieta se aplicó al valor de IRI el índice de jerarquización (DJ), que toma el valor más alto del IRI y calcula el porcentaje de todos los demás valores a partir de él. Si el porcentaje del ítem presa se encuentra incluido entre el 100% y 75% se lo considera fundamental; si se ubica entre el 75% y 50% se lo considera secundario, si esta entre 50% y 25% es accesorio, y si está por debajo del 25% se lo considera accidental (Martori, 1991).
La amplitud del nicho trófico fue calculada a partir de la utilización del índice de Levins (1968).
Nb=(Spij2)-1
Donde, Nb es amplitud del nicho trófico y pij es la probabilidad de hallar el ítem i en la muestra j.
Además se calculó la diversidad trófica (H) según el criterio de Hurtubia (1973), también se calcularon las novedades tróficas acumuladas utilizándose para establecer la muestra mínima de ejemplares en este estudio.
La comparación de las dietas de machos y hembras, se analizó con el índice de correlación de Spearman; donde H0 es: no existe relación entre los contenidos estomacales de ambos sexos. El grado de semejanza entre los contenidos estomacales de machos y hembras fue analizado con el coeficiente de similitud de Jaccard. La relación entre el tamaño de la presa, el LHC y el ancho de la boca se analizó con el índice de correlación de Pearson.
Resultados y discusión
Se analizaron un total de 35 estómagos, de los cuales 7 estuvieron vacíos correspondiendo a 4 machos y 3 hembras.
De los que contuvieron alimento 15 pertenecieron a hembras, cuyo LHC medio fue de 40,56 mm (SD= 12,4; Rango= 55 mm), y el tamaño medio de la boca fue de 14,6 mm (SD= 4,2). El resto de los individuos fueron machos (n= 13), el LHC medio fue de 47,3 (SD= 18,3; Rango= 65,7 mm) y el tamaño medio de la boca fue de 17,3 (SD= 7,8).
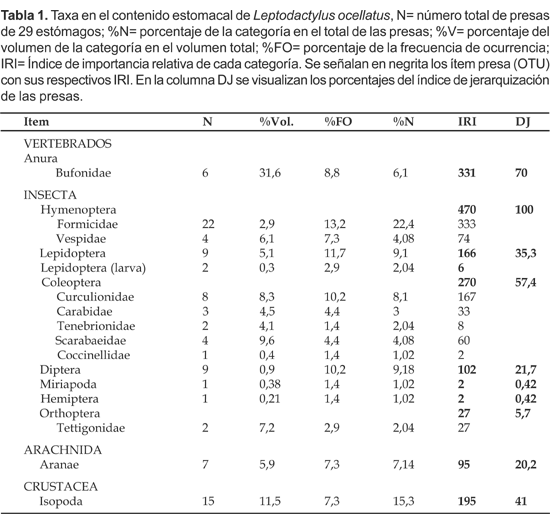
El espectro trófico se basó en la identificación de 117 presas, con un número medio de presas de 4,17 (SD= 4,7) por estómago, con un total de 16 taxa identificados. Siguiendo el criterio de unidad taxonómica operacional (OTU) (Sneath y Sokal, 1973) se consideraron solamente 10 ítem presa: Anura, Hymenoptera, Lepidoptera, Coleoptera, Diptera, Miriapoda, Hemiptera, Orthopera, Aranae e Isopoda. Según el índice de jerarquización agrupamos en: Ítem fundamentales para la dieta: Hymenoptera (100%); los ítem secundarios de la dieta: Anura (70%), Coleoptera (57,4%); Ítem accesorios de la dieta: Isopoda (41%), Lepidoptera (Adultos y Larvas) (35,3%) y como ítem accidentales de la dieta: Aranae (20%), Orthoptera (5,7%), Miriapoda (0,5%), Hemiptera (0,5%).
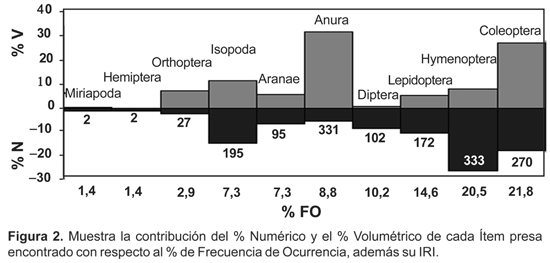
La contribución de cada categoría se ve reflejada por el índice de importancia relativa (IRI), los mayores valores correspondieron a Hymenoptera (407), Anura (331) y Coleoptera (270) (Tabla 1). Las presas con mayor porcentaje de presencia fueron Formicidae que representaron el 22,4%, seguido por Isopoda con el 15,3 % del total de los organismos (Fig. 2).
Se encontró una alta relación lineal y positiva entre los diferentes ítem alimentarios de machos y hembras (Spearman, R= 0,83; p<0,001), lo que indica que las presas son consumidas en proporciones similares por ambos sexos, también se calculó el índice de similitud para los ítem consumidos por machos y hembras arrojando un valor de 0,63 lo que indica que las dietas son similares en un 63%.
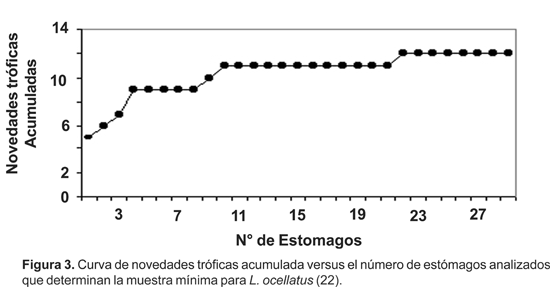
La diversidad trófica (H) fue de 0,16 (SD= 0,11), la figura 3 muestra la curva de novedades tróficas acumuladas que se estabiliza aproximadamente en el estómago número 22 indicando la muestra mínima para este estudio. La amplitud de nicho tuvo un valor de 7,6 siendo menor que el observado por Lajmanovich (1996) para la misma especie (Nb= 11,6); Maneyro et al. (2004) sugieren que L. ocellatus en las diferentes épocas del año y con los diferentes grupos etarios, realiza cambios en las estrategias alimentarias, permutando éstas en un continuo entre generalista y especialista.
No se encontró una relación significativa entre el ancho de la boca del predador y los parámetros que estiman el tamaño medio de la presa como el ancho medio de la presa (Spearman, R= 0,21, p>0,26), el volumen medio de la presa (Spearman, R= 0,28, p>0,14) y el largo medio de la presa (Spearman, R= 0,36, p>0,05), en este último caso observamos una tendencia similar a la encontrada en otros estudios de dieta de anfibios, donde se correlaciona el tamaño de la presa con el tamaño del predador (Basso, 1990; Lajmanovich, 1996).
Literatura citada
Basso N. G., 1990. Estrategias Adaptativas en una Comunidad Subtropical de Anuros. Cuadernos de Herpetología, Serie Monografías N° 1, 70pp.
Cabrera, A. L., 1994. Enciclopedia Argentina de Agricultura y Jardinería, Tomo II, Regiones Fitogeográficas Argentinas, Editorial ACME S.A.C.I.
Cei, J. M., 1980. Amphibians of Argentina. Italian Journal of Zoology, N. S. Monografia 2, 609 pp.
Duellman W. E. & L. Trueb. 1986. Biology of Amphibians. MacGraw-Hill Book Company Pp.:609.
Dunham, A. E., 1983. Realized niche overlap, resourse abundance and intensity of interspecific campetition. In: Huey R.D.; Pianka E.R. & Schoener T. W. (eds.) Lizard Ecology. London, Harvard University. 261-280.
Gallardo, J. M., 1987. Anfibios Argentinos, Guía para su Identificación. Biblioteca Mosaico, Pp.:98.
Hurtubia, J., 1973. Trophic diversity measurement in sympatric predatory species. Ecology, 54(4):885-890.
Lajmanovich, R., 1996. Dinámica Trófica de Juveniles de Leptodactylus ocellatus (Anura: Leptodactylidae), en una isla del Paraná, Santa Fe, Argentina. Cuadernos de Herpetología, Tucumán, 10(1-2): 11-23.
Levins, R., 1968. Evolution in Changing environment. New Jersey, Prisenton, Univ. Press: 1-120.
Maneyro R., D. Naya, I. Rosa, A. Canavero & A. Camargo, 2004. Diet of the South American frog Leptodactylus ocellatus (Anura: Leptodactylidae) in Uruguay. Iheringia, Ser. Zool., Porto Alegre, 94(1):57-61.
Martori, A., 1991. Alimentación de los adultos de Eoproctus asper (Dugés 1853) en la montaña media del Pirineo catalán. (España). Rev. Esp. Herp. 5:23-36.
Pianka, E.P., 1982. Ecología evolutiva. Universidad de Texas, Austin. Editorial Omega, S.A. Pp.: 536
Pinkas L.; M. Oliphant & Z. Iverson. 1971.Food Habitat of albacore bluefin, tuna and bonito in California waters. California Dep. Of Fish an Game Bulletin. 152:1-350.
Sneath P. H. & R. R. Sokal. 1973. Numerical taxonomy. W. H. Freeman and Co. San Francisco.
Correspondencia:
1Departamento de Biología e Instituto y Museo de Ciencias Naturales, Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de San Juan, Avenida España 400 (Norte), Sta. Maria de Oro 367(s), CP: 5400, San Juan, Argentina
E-mail Eduardo Sanabria: sanabria_eduardo@hotmail.com
