










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.22 no.3 Lima Dec. 2015
http://dx.doi.org/10.15381/rpb.v22i3.11432
doi: http://dx.doi.org/10.15381/rpb.v22i3.11432
TRABAJOS ORIGINALES
Bistriopelma, a new genus with two new species from Peru (Araneae: Theraphosidae: Theraphosinae)
Bistriopelma, un género nuevo con dos nuevas especies de Perú (Araneae: Theraphosidae: Theraphosinae)
Radan Kaderka
Address: Tyrsovo namesti 1732, 252 63 Roztoky u Prahy, Czech Republic
Email: radan.kaderka@seznam.cz
Abstract
Bistriopelma gen. nov. from the Andean region in Peru is here described, diagnosed and illustrated, together with two new species Bistriopelma lamasi sp. nov. as a type species and Bistriopelma matuskai sp. nov. This new genus differs from all other theraphosid genera, except the genus Phrixotrichus Simon 1889 (sensu Perafán & Pérez-Miles 2014) and the troglobite species Tmesiphantes hypogeus Bertani, Bichuette & Pedroso 2013, in the presence of two abdominal dorsolateral patches of the type III urticating setae. Bistriopelma gen nov. can be distiguished from Phrixotrichus by the different shape of spermathecae in females, consisting of two separate subparallel seminal receptacles in Bistriopelma gen. nov. but strongly divergent in Phrixotrichus and by the different morphology of male palpal bulb (embolus projecting retrolaterally from tegulum in Bistriopelma gen. nov. against posteriorly in Phrixotrichus); furthermore by the presence of basal spine on prolateral tibial apophysis, the absence of the type IV urticating setae and paired tarsal claws denticulate. Bistriopelma gen. nov. is separated from Tmesiphantes hypogeus by the position of the urticating seta patches (middle of abdomen in Bistriopelma gen. nov. and posterior half in Tmesiphantes hypogeus), the different shape of spermathecae (long separate seminal receptacles with subapical constriction in Bistriopelma gen. nov. and short, subtriangular and widely separated in Tmesiphantes hypogeus) and also in the extension of metatarsal scopulae. Additional comments and a distribution map of all species of this genus are provided.
Keywords: Araneae; Mygalomorphae; Peru; South America; taxonomy.
Resumen
Se describe, diagnostica e ilustra Bistriopelma gen. nov. de la región andina de Perú, con dos nuevas especies Bistriopelma lamasi sp. nov., especie tipo, y Bistriopelma matuskai sp. nov. Este género nuevo se diferencia del resto de arañas terafosidas, excepto el género Phrixotrichus Simon 1889 (sensu Perafán & Pérez-Miles 2014) y la especie cavernícola Tmesiphantes hypogeus Bertani, Bichuette & Pedroso 2013, en la presencia de dos manchas abdominales dorsolaterales de las setas urticantes de tipo III. Bistriopelma gen nov. se diferencia de Phrixotrichus en la morfología de la espermateca, estando compuesta en Bistriopelma gen. nov. de dos receptaculos seminales separados y subparalelos pero considerablemente divergentes en Phrixotrichus, en la morfología de los bulbos pedipalpales de machos (émbolo proyectándose retrolateralmente del tegulum en Bistriopelma gen. nov. contra posteriormente en Phrixotrichus); además en la presencia de la espina basal en la apófisis tibial prolateral, en la ausencia de las setas urticantes de tipo IV y en las uñas tarsales con dientes. Se diferencia de Tmesiphantes hypogeus en la posición de los parches de las setas urticantes (centro del abdomen en Bistriopelma gen. nov. y parte posterior en Tmesiphantes hypogeus), en la morfología de las espermatecas (receptáculos seminales prolongados y separados, subapicalmente estrechados en Bistriopelma gen. nov. y cortos, subtriangulares y considerablemente separados en Tmesiphantes hypogeus) y también en la extensión de las escopulas metatarsales. Se presentan comentarios adicionales y el mapa de distribucion de todas las especies de este género.
Palabras clave: Araneae; Mygalomorphae; Perú; América del Sur; taxonomía.
Introduction
The family Theraphosidae represents the largest and the most studied group of mygalomorph spiders. An exclusive defensive mechanism using modified abdominal barbed setae called "urticating setae" was recorded in the New World representatives belonging to the subfamilies Theraphosinae and partly Aviculariinae. The subfamily Theraphosinae is associated with the presence of type I, III, IV and VI urticating setae and the phylogeny of this group is still a subject of research.
In October 2013, while examining the Theraphosidae spider collection in the Museum of Natural History in Lima, Peru, a single male of an unknown species was discovered. This new taxon differed from known species, except the genus Phrixotrichus Simon 1889 (sensu Perafán & Pérez-Miles 2014) and the troglobite species Tmesiphantes hypogeus Bertani, Bichuette & Pedroso 2013, in the presence of two abdominal dorsolateral patches of the type III urticating setae. Later, in September 2014, a single female of this species was found during the field work near Puquio, Ayacucho, Peru. Due to the location of type III urticating setae in two dorsolateral patches, together with the shape of two separate subparallel seminal receptacles and the strongly elevated eye tubercle which in combination don´t fit any known theraphosid genus, the species was placed in a new theraphosine genus. This genus, including two new species, is here described, diagnosed and illustrated.
Material and methods
Abbreviations: Eye sizes and interdistances: AME = anterior median eyes, ALE = anterior lateral eyes, OQ = ocular quadrangle (including lateral eyes), PME = posterior median eyes, PLE = posterior lateral eyes. Spination: d = dorsal, p = prolateral, r = retrolateral, v = ventral. Male palpal bulb: A = apical keel, E = embolus, PI = prolateral inferior keel, PS = prolateral superior keel, VC = ventral crest. Cheliceral teeth pattern: v = small teeth, V = big teeth, - = space. PLS = posterior lateral spinnerets, PMS = posterior median spinnerets. Collections: BMNH = British Museum of Natural History, London, England. MUSM = Museo de Historia Natural, Lima, Peru. RKCP = Radan Kaderka private collection, Prague, Czech Republic.
Measurements were taken with an ocular micrometer according to the central axis of structures and are given in millimetres. The measurements of the leg and palpal segments were taken dorsally. The eye measurements were taken from the widest spans of the lens, AME in dorsal view, ALE, PLE and PME in dorsolateral view. The measurement of the total body length including cephalothorax and abdomen without spinnerets was made using a calliper.
The extent of tarsal and metatarsal scopulae on ventral side of both leg segments was expressed as a percentage of the total length of segment, from apical end.
The leg spination was described using a following method. Each leg segment was divided into four quadrants (ventral, prolateral, retrolateral and dorsal) and each quadrant described separately in basal, central and apical section. E.g. metatarsus I v 1-0-3 means that in the ventral plane (quadrant) there is one spine in basal section and three spines in apical section. If the bases of all three spine in apical section are located apically, then their position would be described by the term "apical" in brackets as in Bertani (2001). Unequal numbers of spines on the right and left side of the same leg segment were expressed by parentheses.
The denticulation of paired tarsal claws is described separately for prolateral and retrolateral claw (prolateral/retrolateral).
The terminology of male palpal bulb structures follows Bertani (2000). The terminology of subapical apophyses on tibia I follows Vol (2000).
The abdominal urticating setae were removed from the area of urticating setae by forceps, placed in alcohol and examined by a binocular microscope Leica S6D. The terminology of urticating setae follows Cooke et al. (1972). The barbs of urticating setae whose tips are oriented to the seta base are considered as reversed.
The biogeographical regionalisation follows Morrone (2014).
Other material examined: Bistriopelma sp., female from Peru, Ayacucho (RKCP 0701, exuvia); Euathlus truculentus Ausserer 1875, male holotype (BMNH 1590); Euathlus sp., male from Chile, between Santiago de Chile and Valparaiso (RKCP 0293); Phrixotrichus vulpinus Karsch 1880, male and female from Chile (RKCP 0066, 0067).
Taxonomy
Order: Araneae Clerck 1757
Infraorder: Mygalomorphae Pocock 1892
Family: Theraphosidae Thorell 1869
Subfamily: Theraphosinae Thorell 1869
Bistriopelma gen. nov.
Type species: Bistriopelma lamasi sp. nov.
Diagnosis: Bistriopelma gen. nov. differs from all other theraphosid genera, except the genus Phrixotrichus Simon 1889 (sensu Perafán & Pérez-Miles 2014) and the troglobite species Tmesiphantes hypogeus Bertani, Bichuette & Pedroso 2013, in the presence of two abdominal dorsolateral patches of the type III urticating setae, each with a stripe of long reddish-brown thicker setae, arranged diagonally to the longitudinal body axis. Bistriopelma gen nov. can be distiguished from Phrixotrichus by the shape of spermathecae in females, consisting of two separate subparallel seminal receptacles in Bistriopelma gen. nov. but strongly divergent in Phrixotrichus, by the different morphology of the male palpal bulb having the embolus projecting retrolaterally from tegulum (contra posteriorly in Phrixotrichus, Figures 32−33), furthermore by the presence of basal spine on prolateral tibial apophysis (absent in Phrixotrichus), the absence of the type IV urticating setae and by paired tarsal claws denticulate (smooth in Phrixotrichus); from Tmesiphantes hypogeus by the position of urticating seta patch in the middle of abdomen (contra in posterior half in Tmesiphantes hypogeus), the well-developed scopulae on metatarsi I and II covering 90−100% of this leg segment (contra 30% in tarsi I and II in T. hypogeus), the presence of scopulae on metatarsi IV covering 25−30% of this leg segment in Bistriopelma gen. nov. (absent in T. hypogeus) and the different shape of spermathecae (long seminal receptacles with subapical constriction in Bistriopelma gen. nov. contra short, subtriangular and widely separated in T. hypogeus).
Affinities: Bistriopelma gen. nov. shares with Tmesiphantes Simon 1892, Melloleitaoina Gerschman de Pikelin & Schiapelli 1960 and Magulla Simon 1892 the type III urticating setae, the reduced number of labial cuspules, the absence of stridulatory apparatus, similar morphology of male palpal bulb having two prolateral keels only, PS and PI, similar spermathecae consisting of two separate seminal receptacles with subapical constriction, except for Tmesiphantes hypogeus, and similar morphology of subapical apophyses on tibia I consisting of two unequal branches, prolateral tibial apophysis may be reduced or absent in some species.
Bistriopelma gen. nov. can be distinguished from Tmesiphantes by the urticating setae arranged in two dorsolateral patches (contra one patch in Tmesiphantes, except for T. hypogeus – see the Discussion), the absence of sexual dimorphism in urticating setae (only type III setae in both sexes of Bistriopelma gen. nov. contra setae of the type III or III+IV in males and type IV in females in Tmesiphantes nubilus, T. caymmii and T. aridai (Yamamoto et al. 2007; Gonzales-Filho et al. 2014) or type IV in males and type III in females in Tmesiphantes riopretano and T. perp (Guadanucci & Silva 2012)), furthermore by the well-developed scopulae on metatarsi I and II covering 90−100% of this leg segment (contra 0−50% in metatarsi I and 0−30% in metatarsi II in Tmesiphantes) and the presence of scopulae on metatarsi IV covering 25−30% of this leg segment in Bistriopelma gen. nov. but absent or only apically present in Tmesiphantes. Males of Bistriopelma gen. nov. can be distinguished from Tmesiphantes by the mutual position of PS and PI keel which are in close proximity in Bistriopelma gen. nov. (Figures 9−10) but widely separated in Tmesiphantes (Yamamoto et al. 2007: figures 3−5, 9−11, 14−16, 21−23; Guadanucci & Silva 2012: figures 3−5, 11−13; Gonzales-Filho et al. 2014: 1−3; Fabiano-da-Silva et al. 2015: figures 2−4).
Bistriopelma gen. nov. can be distinguished from Melloleitaoina by the urticating setae arranged in two dorsolateral patches (contra one patch in Melloleitaoina), the absence of sexual dimorphism in urticating setae (only type III setae in both sexes of Bistriopelma gen. nov. contra setae of intermediate morphology between the types III and IV in males and type IV setae in females in Melloleitaoina uru and M. yupanqui (Perafán & Pérez-Miles 2014)), furthermore by the well-developed scopulae on metatarsi I and II covering 90−100% of this leg segment (contra 5−50% in metatarsi I and 5−30% in metatarsi II in Melloleitaoina) and the presence of scopulae on metatarsi IV covering 25−30% of this leg segment in Bistriopelma gen. nov. but absent in Melloleitaoina. Males of Bistriopelma gen. nov. can be distinguished from Melloleitaoina by the mutual position of PS and PI keel which are in close proximity in Bistriopelma gen. nov. (Figures 9−10) but widely separated in Melloleitaoina (Perafán & Pérez-Miles 2014: figures 3−5, 9−10, 16−17, 26−27).
Bistriopelma gen. nov. can be distinguished from Magulla by the absence of the type IV urticating setae and the shape of the urticating seta patch (two dorsolateral patches of the type III setae in Bistriopelma gen. nov. contra two dorsolateral patches of the type III setae joined with a dorsal patch of type IV setae in Magulla), furthermore by the well-developed scopulae on metatarsi I and II covering 90−100% of this leg segment (contra 16−50% in metatarsi I and 16−40% metatarsi II in Magulla) and the presence of scopulae on metatarsi IV covering 25−30% of this leg segment in Bistriopelma gen. nov. but absent in Magulla. Males of Bistriopelma gen. nov. can be distinguished from Magulla by the mutual position of PS and PI keel which are in close proximity in Bistriopelma gen. nov. (Figures 9−10) but widely separated in Magulla (Indicatti et al. 2008: figures 2 A−C, F−H, K−M). Females of Bistriopelma gen. nov. can be distinguished from Magulla by the absence of subapical ventral loop on seminal receptacles (present in Magulla).
Bistriopelma gen. nov. shares with Phrixotrichus Simon 1889 the urticating setae arranged in two dorsolateral patches, the presence of type III urticating setae, the reduced number of labial cuspules, with Euathlus Ausserer 1875 and Phrixotrichus the the absence of stridulatory apparatus, the presence of two subequal prolateral keels in male palpal morphology, PS and PI, and the similar morphology of subapical apophyses on tibia I consisting of two unequal branches. It differs from Phrixotrichus and Euathlus in the absence of type IV urticating setae, the presence of denticulation on paired tarsal claws, the spermathecal shape and the male palpal bulb morphology (embolus is projecting retrolaterally from tegulum in Bistriopelma gen. nov. contra posteriorly in Phrixotrichus and Euathlus, see Figures 32−33, 34−36; Perafán & Pérez-Miles 2014: figures 4A, 4B, 5A, 5B, 6C, 6D, 7A, 7B, 8B, 8C, 9A, 9B, 10A, 10B). The separate seminal receptacles in Bistriopelma sp. nov. are subparallel but strongly divergent in Phrixotrichus and Euathlus (Perafán & Pérez-Miles 2014).
Bistriopelma gen. nov. shares with Agnostopelma Pérez-Miles & Weinmann 2010 the urticating setae of the type III, the reduced number of labial cuspules, the denticulation of paired tarsal claws, the absence of stridulatory apparatus and the presence of two prolateral keels, PS and PI, in male palpal bulb. Bistriopelma gen. nov. can be distinguished from Agnostopelma Pérez-Miles & Weinmann 2010 by the absence of the type IV urticating setae, the presence of scopulae on tarsi IV (absent in Agnostopelma), the presence of two subapical apophyses on tibia I (none in Agnostopelma), the different morphology of male palpal bulb (embolus long and tapering, projecting retrolaterally from tegulum in Bistriopelma gen. nov. contra short and stout embolus projecting posteriorly in Agnostopelma), the well-developed scopulae on metatarsi I and II covering 90−100% of this leg segment (contra 0−50% in metatarsi I and II in Agnostopelma), the presence of scopulae on metatarsi IV covering 25−30% of this leg segment in Bistriopelma gen. nov. but absent in Agnostopelma and the different spermathecal shape in females (seminal receptacles are separated in Bistriopelma gen. nov. but basally fused in Agnostopelma).
Etymology: The generic name refers to the abdominal morphology with the type III urticating setae arranged in two (lat. bi-) dorsolateral patches, each with a stripe (lat. stria) of long reddish-brown thicker setae, arranged diagonally to the longitudinal body axis.
General description
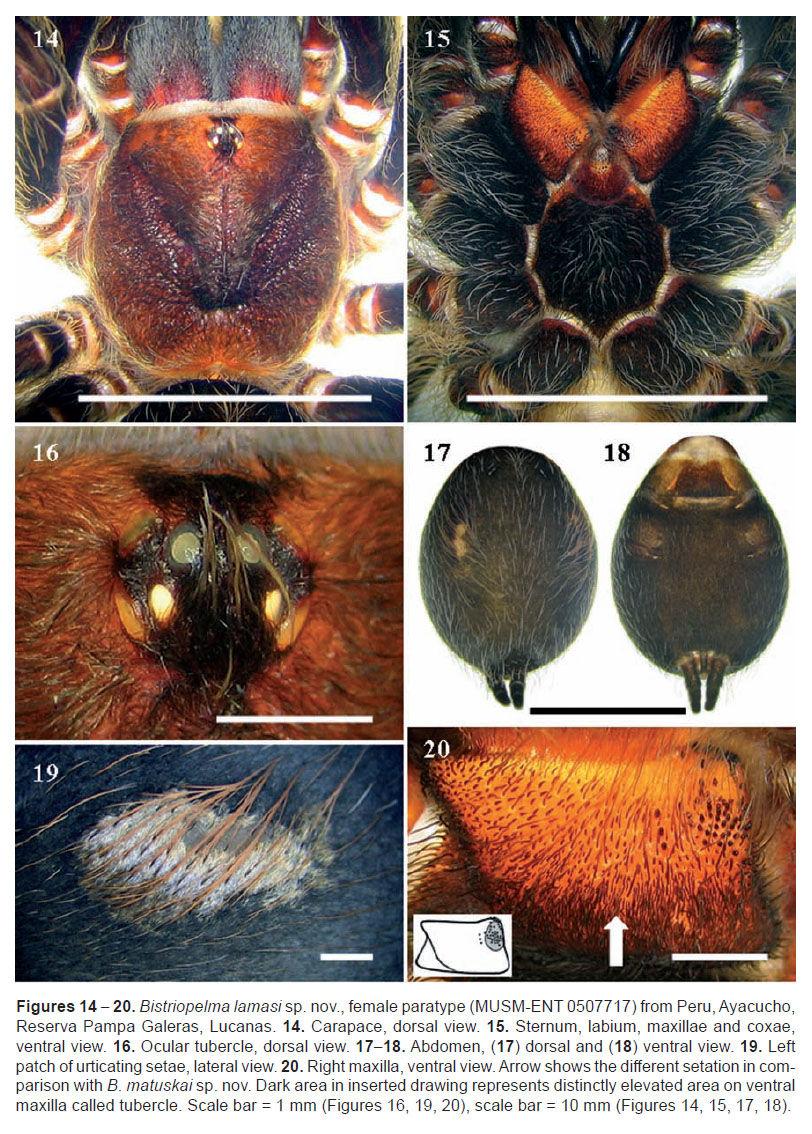
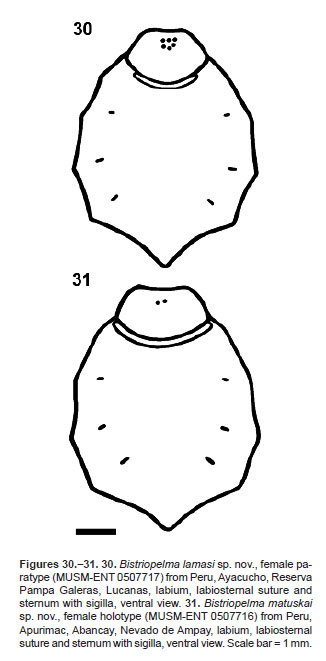
The genus Bistriopelma comprises small up to medium-sized spiders, with total length 19.3 up to 24.8 mm, exluding chelicerae and spinnerets. Carapace oval, uniformly coloured. Caput domed. Ocular tubercle oval (Figures 4, 16, 25), strongly elevated, distinctly wider than longer, with eight eyes, anterior eye row slightly procurved, posterior row recurved in dorsal view. Clypeus indistinct to narrow. Fovea transverse, straight, deep or absent in Bistriopelma matuskai sp. nov. and Bistriopelma sp. from Peru, Ayacucho. Chelicerae without rastellum and stridulatory bristles, with teeth on promargin (7−10), first basal teeth are complemented with granulation. Labium domed, wider than longer, with reduced number of cuspules in anterior third (2−5), maxillae with 88−142 cuspules in basal third on ventral side. Almost all cuspules are located on indistinctly or distinctly elevated area called tubercle in the following text. Anterior apical two thirds of ventral maxillae covered with very short spiniform setae (Figures 20, 29). Maxillary lobe pronounced into conical process. Labiosternal groove distinct, shallow and flat, with two joined elongated sigilla. Sternum oval, with three pairs of small, oval sigilla located near coxae III, coxae II and coxae I, posteriorly separated from the margin approximately by their own 3.5−7.0 diameters. Legs uniformly hirsute. Leg pattern (from longest to shortest): I>IV>II>III (male holotype of Bistriopelma lamasi sp. nov.; female holotype Bistriopelma matuskai sp. nov.) or IV>I>II>III (female paratype of Bistriopelma lamasi sp. nov.). Leg segments: generally uniform to slightly incrassate on femur III in males.
Dense scopulae on ventral side of all tarsi, metatarsi I−II 90−100% scopulate, posterior metatarsi partly scopulate, scopulae more extended on anterior than on posterior legs. Tarsal scopulae I, II usually undivided, on tarsi III, IV usually divided by longitudinal band of setae. Retrolateral side of femur IV and prolateral side of femur I without pad of plumose setae. Maxillary and trochanteral stridulatory setae or bristles absent. Spination as in species descriptions. Dorsal face of all tarsi and cymbium with two irregular longitudinal rows of very short claviform trichobothria. Paired tarsal claws with teeth (0−6), third claw absent in all tarsi. Claw tufts dense, bilobate, present on all tarsi.
Abdomen uniformly coloured, with two pale dorsolateral stripes of urticating setae (Figures 7, 19, 28). Urticating setae of type III. Four spinnerets present. PLS composed of three digitiform segments. PMS digitiform, mono-segmentated.
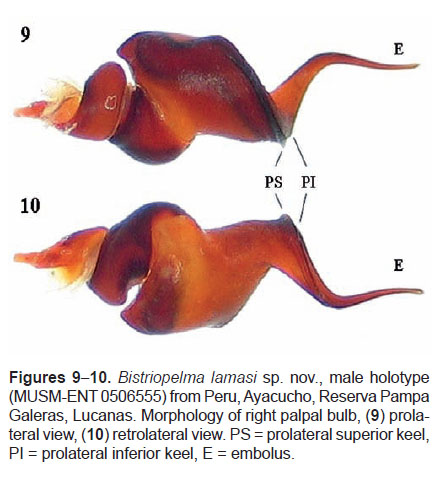
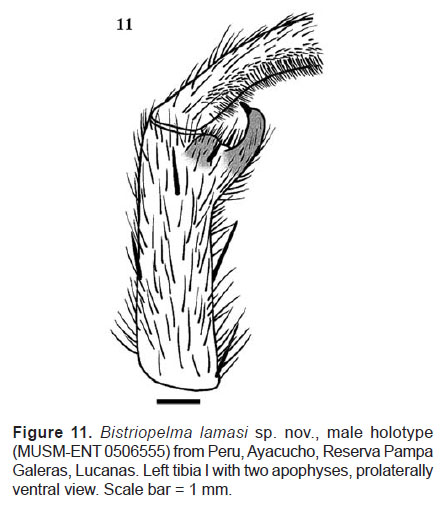
Male palpal organ with a narrow embolus projecting retrolaterally from tegulum, sigmoidly curved (Figures 9−10), with two close subparallel prolateral keels, PS and PI. Tegulum with short tegular apophysis, projecting anteriorly. Cymbium dorsally with spiniform setae, prolateral cymbial lobe distinctly larger than retrolateral lobe. Two unequal tibial apophyses are present on tibia I (Figure 11): a longer ventral tibial apophysis, with short apical spine and a shorter prolateral tibial apophysis with single, retrolateral spine at base. Metatarsus I not sigmoidly curved, without basal or median protuberance on retrolateral face. It contacts the ventral tibial apophysis when flexed.
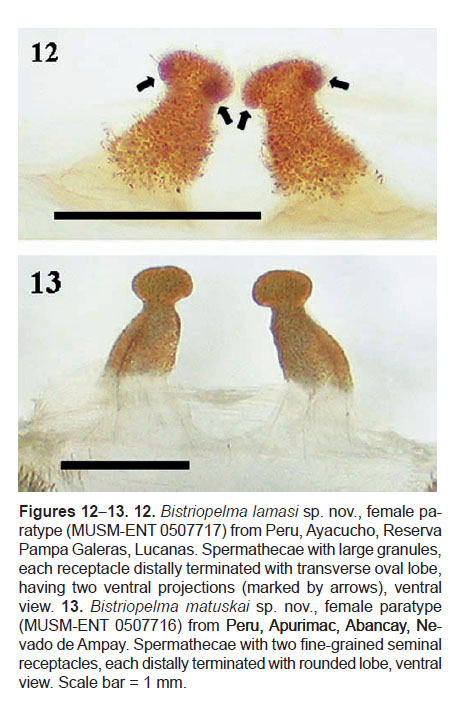
Females with spermathecae composed of two separated seminal receptacles, each with subapical constriction and without sclerotized basal plates (Figures 12, 13). Both oval apical lobes in Bistriopelma lamasi sp. nov. with two ventral sclerotized projections (Figure 12). Both seminal receptacles can be sclerotized with large granules or fine-grained.
Bistriopelma lamasi sp. nov.
Figures 1−12, 14−20, 30, Table 1−2, 4

Types: Male holotype (MUSM-ENT 0506555) from Peru, Ayacucho, Reserva Pampa Galeras, Lucanas, altitude 4.000 m a.s.l., April 21, 1974, G. Lamas M. col.; female paratype (MUSM-ENT 0507717) from Peru, Ayacucho, Reserva Nacional Pampa Galeras, Lucanas, 14°37.533'S 74°17.355'W, 3.749 m a.s.l., 25.IX.2014, R. Kaderka col.
Diagnosis: Females of Bistriopelma lamasi sp. nov. differ from B. matuskai sp. nov. in the presence of a deep fovea (absent in B. matuskai sp. nov.), seminal receptacles with large granules (fine-grained in B. matuskai), different leg pattern in females (IV>I>II>III in B. lamasi sp. nov. and I>IV>II>III in B. matuskai sp. nov.), the well-developed basal tubercle covered with cuspules (Figure 20), indistinct in B. matuskai sp. nov., and different setation on ventral maxilla (see Figures 20, 29; marked by arrows).


Etymology: The specific epithet is dedicated to Dr. Gerardo Lamas Müller, Peruvian entomologist and systematist specialized in butterflies (Lepidoptera), who collected this species approximately fourty years ago.
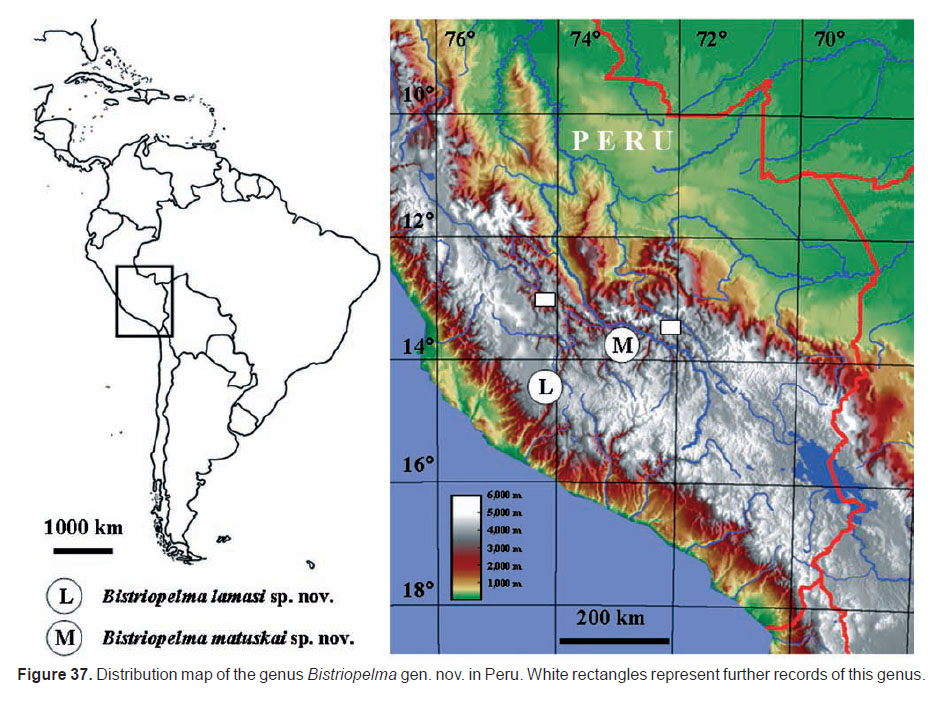
Distribution (Figures 2, 37): Known only from the type locality.
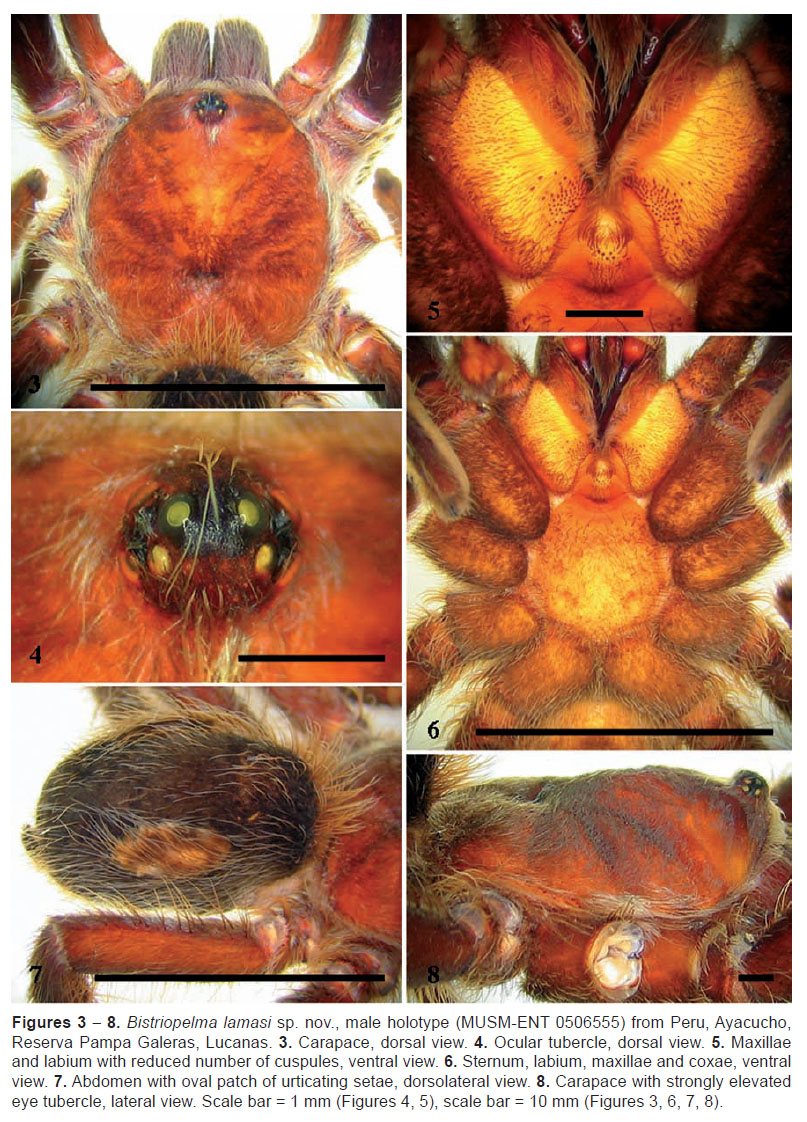
MALE (MUSM-ENT 0506555): Total length: 19.26, carapace (Figure 3), length 9.36, width 8.75, chelicerae with 8 teeth on promargin. Cheliceral teeth pattern from the basal end: right side: V-V-VVVV-V-V. Left side: V-V-VVVV-V-V. Anterior eye row slightly procurved, posterior eye row slightly recurved. Eye sizes and interdistances: AME 0.286 (circular), ALE 0.317 (subcircular), PME 0.234 (oval), PLE 0.286 (subcircular), AME-AME 0.234, AME-ALE 0.083, PME-PME 0.572, PME-PLE 0.078, ALE-PLE 0.156, AME-PME 0.057, OQ length 0.689, width 1.261. Ocular tubercle strongly elevated, almost rounded (Figures 4, 8), length 1.092, width 1.261, clypeus narrow, indistinct. Fovea transverse, straight, deep, width 1.17, 6.62 from the anterior edge of carapace. Labium length 1.35, width 1.76, anterior third with 5 cuspules, maxillae with 111−112 cuspules, located in basal tubercles, except few of them (Figure 5). Sternum (Figure 6), length 4.47, width 4.27, three pairs of sternal sigilla located near coxae III (oval 0.221, 0.78 from the edge of sternum), coxae II (oval 0.156, 0.65 from the edge of sternum) and near coxae I. Leg pattern: I>IV>II>III. Incrassate leg segments: slightly incrassate femur III.
Scopulae: All tarsi, 100% scopulate, metatarsi I-II 100%, metatarsi III 50% scopulate, metatarsi IV 25% scopulate. Tarsi I with scopula entire, tarsi II with scopula entire, with very thin longitudinal row of long setae, tarsi III-IV divided by a band of setae. Dorsal face of all tarsi and cymbium with two irregular longitudinal rows of very short claviform trichobothria. Denticulation pattern of paired tarsal claws on right leg (prolateral / retrolateral row): I 6/6, II 6/5, III 2/2, I 2/2. Plumose setae on retrolateral face of femur IV absent. Maxillary and trochanteral stridulatory bristles absent.
Spination: femora I p 0-0-1, II d 0-2-1, III d 0-0-2, IV 0 and femora of palps 0; patellae I 0, II p 0-1-1, III−IV 0 and patellae of palps 0; tibiae I v 2-1-1, p 0-1-1, II v 0-3-3 (apical), p 0-1-1, r 0-1-0, III v 1-2-0, p 2-2-1, r 1-1-0, IV p 0-2-1, r 1-1-1 and tibiae of palps p 0-1-1; metatarsi I v 0-0-1 (apical), II v 0-0-3 (apical), p 1-1-1, r 0-1-0, III v 3-2-3 (apical), p 1-1-1, r 1-1-1, IV v 2-2-3 (apical), p 0-1-1, r 1-1-1, tarsi I−IV and tarsi of palps 0.
Palpal bulb (Figures 9−10): with a narrow embolus projecting retrolaterally from tegulum, sigmoidly curved, and two close subparallel keels, PS and PI, PI without tooth or teeth. Tegulum with short tegular apophysis. Cymbium dorsally with spiniform setae, prolateral cymbial lobe distinctly larger than retrolateral lobe, not covered with spiniform setae. Two unequal tibial apophyses present on tibia I (Figure 11): a ventral tibial apophysis with a single apical spine, a prolateral tibial apophysis with a single inner spine, approximately of the same length as the apophysis. Metatarsus I contacts the ventral tibial apophysis when flexed.
Abdomen (Figure 7) with type III urticating setae located in two dorsolateral patches, 4.06 distant from each other. Urticating setae patch: length 3.81, width 1.37. Abdomen length 9.77, width 6.31. PLS triarticulated, length 2.97, basal segment 1.46, middle segment 0.55, apical segment 0.96, all digitiform. PMS monoarticulated, length 0.91.
Coloration: lost due to a long preservation in alcohol.
FEMALE (MUSM-ENT 0507717) (Figure 1): Total length: 24.80, carapace length 9.57, width 8.86, caput elevated (Figure 14), chelicerae with 10 teeth on promargin, with granulation near last two basal teeth. Cheliceral teeth pattern from the basal end: right side: V-VvvvvVVvV. Left side: V-VvvvvvVVV. Anterior eye row slightly procurved, posterior eye row recurved. Eye sizes and interdistances: AME 0.338 (circular), ALE 0.377 (oval), PME 0.260 (oval), PLE 0.398 (oval), AME-AME 0.156, AME-ALE 0.104, PME-PME 0.572, PME-PLE 0.065, ALE-PLE 0.182, AME-PME 0.112, OQ length 0.910, width 1.287. Ocular tubercle almost circular, strongly elevated (Figure 16), length 1.274, width 1.287, clypeus narrow. Fovea transverse, straight, width 1.38, 7.13 from the anterior edge of carapace. Labium length 1.52, width 2.05, anterior third with 5 cuspules, maxillae with 88−142 cuspules, located in distinct basal tubercles, except few of them (Figure 20). Sternum (Figures 15, 30), length 5.40, width 4.52, three pairs of small and narrow sternal sigilla located near coxae III (oval 0.140, 0.962 from the edge of sternum), coxae II (oval 0.130, 0.910 from the edge of sternum) and near coxae I (oval 0.104, 0.728 from the edge of sternum). All sigilla distant approximately 7 diameters from the edge of carapace. Sternum covered with long pale setae. Leg pattern: IV>I>II>III. Incrassate leg segments absent. Maxillary and trochanteral stridulatory bristles absent.
Scopulae: All tarsi 100% scopulate, metatarsi I-II 100%, metatarsi III 40% scopulate, metatarsi IV 30% scopulate. Tarsi I-II with scopula entire, tarsi III-IV divided by a wide band of setae. Dorsal face of all tarsi with two irregular longitudinal rows of very short claviform trichobothria. All paired tarsal claws with 2−4 teeth on each segment: I: 4/4, II: 3/4, III: 2/3, IV: 2/2. Plumose setae on retrolateral face of femur IV absent. Prolateral face of coxa I densely covered with short spiniform setae in basal half.
Spination: femora I−IV and femora of palps 0; patellae I−IV and patellae of palps 0; tibiae I 0, II v 0-0-1 (apical), III v 0-0-2 (apical), p 0-1-0, IV 0 and tibiae of palps v 0-0-2 (apical); metatarsi I v 0-0-1 (apical), II v 1-0-3 (apical), III v 0-0-2 (apical), p 1-1-1, r 0-0-1, IV v 0-1-3 (apical), p 0-1-1, r 0-1-0, tarsi I−IV and tarsi of palps 0.
Spermatheca (Figure 12): two separated seminal receptacles with large granules, each distally terminated with transverse oval lobe, having two ventral projections.
Abdomen (Figures 17, 18) with type III urticating setae located in two dorsolateral patches (Figure 19), 6.30 distant from each other. Urticating setae patch: length 4.83, width 2.44. PLS triarticulated, length 4.00, basal segment 1.74, middle segment 1.07, apical segment 1.19, all digitiform. PMS monoarticulated, length 1.20.
Coloration: dorsal view: dark brown, legs and abdomen covered with long pale setae, except for two abdominal dorsolateral patches of urticating setae covered with long reddish-brown thicker setae, arranged diagonally to the longitudinal body axis. Patellae I, II and palpal patella with two almost equal longitudinal stripes without covering pubescens, patellae III, IV with two unequal diagonal stripes. Tibiae with two longitudinal stripes. Ventral view: dark brown, except reddish brown maxillae and labium. Legs covered with long pale setae.
Bistriopelma matuskai sp. nov.
Figures 13, 21−29, 31, Table 3, 4


Types: Female holotype (MUSM-ENT 0507716) from Peru, Apurimac, Abancay, Nevado de Ampay, 13°34.182'S 72°53.791'W, 4.003 m a.s.l., September 23, 2014, R. Kaderka col.; female paratype (MUSM-ENT 0507712) from Peru, Apurimac, Abancay, Nevado de Ampay, 13°34.147'S 72°54.396'W, 4.398 m a.s.l., September 23, 2014, R. Kaderka col.
Diagnosis: Females of Bistriopelma matuskai sp. nov. differ from B. lamasi sp. nov. in the absence of fovea, fine-grained seminal receptacles in females (large granules in B. lamasi), the different leg pattern in females (I>IV>II>III in B. matuskai sp. nov. and IV>I>II>III in B. lamasi sp. nov.), the undeveloped basal tubercle covering with cuspules and different setation of ventral maxilla (Figures 20, 29).
Etymology: The specific epithet is dedicated to Ondřej Matuška, my brother-in-law, who accompanied me during my field research.
Distribution (Figures 22, 37): Known only from the type locality.
FEMALE HOLOTYPE (MUSM-ENT 0507716) (Figure 21): Total length: 24.39, carapace (Figure 23), length 10.27, width 9.09, chelicerae with 8-10 teeth on promargin. Cheliceral teeth pattern from the basal end: right side: V-VVVVVV-V. Left side: V-V-VVVVVVV-V. Anterior eye row slightly procurved, posterior eye row recurved. Eye sizes and interdistances: AME 0.343 (circular), ALE 0.351 (oval), PME 0.299 (drop-like), PLE 0.364 (oval), AME-AME 0.273, AME-ALE 0.247, PME-PME 0.780, PME-PLE 0.083, ALE-PLE 0.286, AME-PME 0.195, OQ length 0.845, width 1.664. Ocular tubercle strongly elevated, length 1.404, width 1.664 (Figure 25), clypeus narrow, 0.286. Fovea absent, the posterior end of caput 7.4 from the anterior edge of carapace. Labium length 1.40, width 2.39, anterior third with 2 cuspules, maxilla with 94−115 cuspules, located in indistinct basal tubercles (Figure 29). Sternum (Figures 24, 31), length 5.57, width 4.67, three pairs of sternal sigilla located near coxae III (oval 0.255, 1.040 from the edge of sternum), coxae II (oval 0.177, 0.754 from the edge of sternum) and near coxae I (oval 0.156, 0.546 from the edge of sternum). All sigilla distant approximately 3.5−4.3 diameters from the edge of carapace. Sternum without long pale setae. Leg pattern: I>IV>II>III. Incrassate leg segments absent. Maxillary and trochanteral stridulatory bristles absent.



Scopulae: All tarsi, 100% scopulate, metatarsi I 100%, metatarsi II 90%, metatarsi III 50% scopulated, metatarsi IV 25% scopulate. Tarsi I-II with scopula entire, tarsi III-IV divided by a wide band of setae. Dorsal face of all tarsi with two irregular longitudinal rows of very short claviform trichobothria. All paired tarsal claws with 0−4 teeth on each segment: I: 4/3, II: 2/2, III: 0/0, IV: 2/3. Plumose setae on retrolateral face of femur IV absent, retrolateral face partly bare. Prolateral face of coxa I densely covered with short spiniform setae.
Spination: femora I−IV and femora of palps 0; patellae I−IV and patellae of palps 0; tibiae I 0, II v 0-0-1 (apical), III v 0-0-1 (apical), IV 0 and tibiae of palps 0; metatarsi I 0, II v 0-0-3 (apical), III v 0-(0−1)-2 (apical), p 0-1-1, r 0-1-1, IV v 0-1-2 (apical), r 0-1-1, tarsi I−IV and tarsi of palps 0.
Spermathecae (Figure 13): two separated seminal receptacles, fine-grained, each distally terminated with rounded lobe.
Abdomen (Figures 26, 27) with type III urticating setae located in two dorsolateral patches (Figure 28), 4.60 distant from each other. Urticating setae patch: length 4.42, width 2.03. PLS triarticulated, length 4.37, basal segment 1.95, middle segment 1.12, apical segment 1.30, all digitiform. PMS monoarticulated, length 1.17.
Coloration: dorsal view: dark brown, legs and abdomen covered with long pale setae, except two abdominal dorsolateral patches of urticating setae covered with long reddish-brown thicker setae, arranged diagonally to the longitudinal body axis. Patellae I, II and palpal patella with two almost equal longitudinal stripes without covering pubescens, patellae III, IV with two unequal diagonal stripes. Tibiae with two such longitudinal stripes, more distinct in posterior legs. Ventral view: dark brown, except reddish brown maxillae and labium.
MALE: unknown.

Biological and ecological notes: The genus Bistriopelma gen. nov. comprises the fossorial, cold-adapted spiders inhabiting altitudes of approximately 3,500−4,500 m a.s.l. in the Peruvian Andes, in Puna biogeographical province (Figures 2, 22, 37). The typical habitat for this genus are mountain grass slopes covered with stones under which spiders have their burrows. The average annual temperature in distribution area of both new species is 9.5−11.2°C and the temperature in these altitudes may fall to near zero. The average annual precipitation in distribution area is 600−800 mm, with a maximum in January-March (100−150 mm) and with a rainfall minimum in May-August (5−15 mm).
Discussion
Apart from the genera Bistriopelma gen. nov. and Phrixotrichus, Tmesiphantes hypogeus is another species with the type III urticating setae arranged in two dorsolateral patches. However, both patches are located in posterior half of abdomen (contra middle of abdomen in Bistriopelma gen. nov. and Phrixotrichus). As the male of Tmesiphantes hypogeus is unknown, this species was putatively included in Tmesiphantes because of the almost circular sternum, the low number of labial cuspules, the presence of the type III urticating setae, widely separated seminal receptacles that taper to their apex and the geographical affinity (Bertani et al. 2013).
A similar arrangement of urticating setae in two dorsolateral patches is also present in Magulla but both patches are joined with a bigger dorsal patch of the type IV setae (Indicatti et al. 2008). Magulla also shares with Bistriopelma gen. nov. the elevated caput, a weak spination, especially in females (26−29 spines in described types of Magulla and 17−24 in both types of Bistriopelma gen. nov.) and denticulation of paired tarsal claws.
While Bistriopelma gen. nov. inhabits Puna biogeographical province in the Peruvian Andes and Melloleitaoina, a closely related genus to Tmesiphantes (Pérez-Miles et al. 1996; Yamamoto et al. 2007), the adjacent Yungas and Monte biogeographical provinces in the northern Argentina (Perafán & Pérez-Miles 2014), the genera Tmesiphantes and Magulla are known only from the Xingu-Tapajós, Atlantic and Parana Forest biogeographical provinces in the most eastern Brazilian states (Yamamoto et al. 2007; Indicatti et al. 2008; Guadanucci & Silva 2012; Bertani et al. 2013; Gonzales-Filho et al. 2014; Fabiano-da-Silva et al. 2015). Fabiano-da-Silva et al. (2015) examined all species of the genera Tmesiphantes and Melloleitaoina and the authors are persuaded that all previous diagnoses fail to separate them from each other due to interspecific morphological variations and it is very likely that these two genera are synonym. (Fabiano-da-Silva et al. 2015). A molecular analysis based on an appropriate set of loci would help to resolve the mutual position of both taxa which are known to date only from two geographically isolated areas.
On the other hand, representatives of the genera Euathlus and Phrixotrichus are known only from Chile and Argentina (Nequén, Catamarca), from both sides of the Andes (Perafán & Pérez-Miles 2014; Schiapelli & Gerschman de Pikelin 1963). The Phrixotrichus species were found in low altitudes (50−150 m a.s.l.) in the central Chile but some Euathlus species in altitudes reaching up to 3,200 m a.s.l. in the southern end of the Atacaman biogeographical province in the northern Chile (Perafán & Pérez-Miles 2014).
Finally, Bistriopelma gen. nov. and Agnostopelma occur in similar habitats of the Andean mountain range, preferring altitudes above 3,000 m a.s.l., with an average annual temperature of 9−11°C. However, in contrast to Bistriopelma gen. nov. Agnostopelma inhabits the Paramo biogeographical province in the Colombian Andes.
The unique biogeographic area occupied by Bistriopelma gen. nov. and its distinctive morphological characteristics bring support to the naming of a new taxon for this group of theraphosid spiders.
Acknowledgement
I would like to express my gratitude to Diana Silva Dávila from Museo de Historia Natural, Lima, Peru for her assistance during my visit to the MUSM spider collection and for comments on an earlier version of this manuscript, to Rogério Bertani and two anonymous reviewers for valuable comments on the manuscript and to my wife for her support and love.
Literature cited
Bertani R. 2000. Male palpal bulbs and homologous features in Theraphosinae (Araneae: Theraphosidae). The Journal of Arachnology, 28: 29−42. http://dx.doi.org/10.1636/0161-8202(2000)028[0029:MPBAHF]2.0.CO;2
Bertani R. 2001. Revision, cladistic analysis, and zoogeography of Vitalius, Nhandu, and Proshapalopus; with notes on other theraphosine genera (Araneae, Theraphosidae). Arquivos de Zoologa, São Paulo, 36 (3): 265−356. http://dx.doi.org/10.11606/issn.2176-7793.v36i3p265-356
Bertani R., M.E. Bichuette & D.R. Pedroso. 2013. Tmesiphantes hypogeus sp. nov. (Araneae: Theraphosidae), the first troglobitic tarantula from Brazil. Anais da Academia Brasileira de Ciências, 85: 235−243. http://dx.doi.org/10.1590/S0001-37652013005000007
Clerc C. 1757. Aranei Svecici, Descriptionibus et Figuris Æneis Illustrati, ad Genera Subalterna Redacti, Speciebus ultra LX Determinati. Regiae Societatis Scientiarum Upsaliensis, Stockholmiae. 154pp.
Cooke J.A.L., V.D. Roth & F.H. Miller. 1972. The urticating hairs of theraphosid spiders. American Museum Novitates, 2498: 1−43.
Fabiano-da-Silva W., J.P.L. Guadanucci & M.B. Da Silva. 2015. Tmesiphantes mirim sp. nov. (Araneae: Theraphosidae) from the Atlantic Forest of Bahía, Brazil, biogeographical notes and identification keys for species of the genus. Zoologia, 32 (2): 151−156. http://dx.doi.org/10.1590/S1984-46702015000200006
Gonzales-Filho H.M.O., A.D. Brescovit & S.M. Lucas. 2014. A new species of Tmesiphantes (Araneae, Theraphosidae). Inheringia, série Zoologia, Porto Alegre, 104 (2): 223−227. http://dx.doi.org/10.1590/1678-476620141042223227
Guadanucci J.P.L. & W.F. Silva. 2012. Two new species of Tmesiphantes Simon (Araneae: Mygalomorphae: Theraphosidae). Studies on Neotropical Fauna and Enviroment, 47 (2): 139−145. http://dx.doi.org/10.1080/01650521.2012.688559
Indicatti R.P., S.M. Lucas, J.P.L. Guadanucci, F.U. Yamamoto. 2008. Revalidation and revision of the genus Magulla Simon 1892 (Araneae, Mygalomorphae, Theraphosidae). Zootaxa, 1814: 21−36.
Morrone J.J. 2014. Biogeographical regionalisation of the Neotropical region. Zootaxa, 3782: 1−110. http://dx.doi.org/10.11646/zootaxa.3782.1.1
Perafán C. & F. Pérez-Miles. 2014. The Andean tarantulas Euathlus Ausserer, 1875, Paraphysa Simon, 1892 and Phrixotrichus Simon, 1889 (Araneae, Theraphosidae): phylogenetic analysis, genera redefinition and new species descriptions. Journal of Natural History: 1−30. http://dx.doi.org/10.1080/00222933.2014.902142
Perafán C. & F. Pérez-Miles. 2014. Three new species of Melloleitaoina Gerschman and Schiapelli, 1960 (Araneae, Mygalomorphae,Theraphosidae) from northern Argentina. ZooKeys, 404: 117−129. http://dx.doi.org/10.3897/zookeys.404.6243
Pérez-Miles F., S.M. Lucas, P.I. Da Silva, R. Bertani. 1996. Systematic revision and cladistic analysis of Theraphosinae (Araneae: Theraphosidae). Mygalomorph, 1 (3): 33−68.
Pérez-Miles F. & D. Weinmann. 2010. Agnostopelma: a new genus of tarantula without a scopula on leg IV (Araneae, Theraphosidae, Theraphosinae). Journal of Arachnology, 38 (1): 104−112. http://dx.doi.org/10.1636/A07-60.1
Pocock R.I. 1892. Liphistius and its bearing upon the classification of spiders. Annals and Magazine of Natural History, 10 (6): 303−314.
Schiapelli R.D., B.S. Gerschman de Pikelin. 1963. Los generos chilenos Phrixotrichus Simon, 1889 y Paraphysa Simon, 1892 (Theraphosidae, Araneae) en la Argentina: nuevas citas de algunas arañas comunes a ambos países. Revista de la Sociedad entomológica Argentina, 26: 103−108.
Thorell T. 1869. On European spiders. Nova Acta Regiae Societatis Scientiarum Upsaliensis, (3) 7: 109–242.
Vol F. 2000. Description de Bonnetina cyaneifemur, gen. n. & sp. n. (Araneae, Theraphosidae, Theraphosinae) du Mexique. Arachnides, 44: 2−9.
Yamamoto F.U., S.M. Lucas, J.P.L. Guadanucci & R.P. Indicatti. 2007. Revision of the genus Tmesiphantes Simon (Araneae, Mygalomorphae, Theraphosidae). Revista Brasileira de Zoologia, Vol. 24, n. 4: 971−980.
Presentado: 06/03/2015
Aceptado: 08/10/2015
