










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.24 no.4 Lima Dec. 2017
http://dx.doi.org/10.15381/rpb.v24i4.14063
TRABAJOS ORIGINALES
Diferenciación morfológica de Carollia brevicauda y C. perspicillata (Chiroptera: Phyllostomidae) de Perú y Ecuador
Morphological differentiation of Carollia brevicauda and C. perspicillata (Chiroptera: Phyllostomidae) from Peru and Ecuador
Dennisse Ruelas
Universidad Nacional Mayor de San Marcos, Museo de Historia Natural, Av. Arenales 1256, Lima 11, Perú
E-mail Dennisse Ruelas: dennisse.ruelas@unmsm.edu.pe
Resumen
En el género Carollia, Carollia brevicauda y C. perspicillata son las especies más abundantes y ampliamente distribuidas del género en América del Sur, además de tener distribuciones casi complemente sobrepuestas. Debido a que estas especies son morfológicamente similares en algunas localidades son frecuentemente mal identificadas. El objetivo de este estudio fue determinar las diferencias morfológicas y morfométricas de C. brevicauda y C. perspicillata. para esto, se empleó un análisis morfológico detallado y diversos métodos estadísticos y multivariados. Se evaluaron 26 variables cráneo-dentales con distribución normal de 375 especímenes adultos entre ambas especies procedentes de poblaciones peruanas y ecuatorianas. Además, para efectos de comparación se incluyeron a 54 especímenes entre Carollia benkeithi, C. manu y Carollia sp. Se encontró que C. brevicauda y C. perspicillata son diferentes en 28 caracteres discretos y significativamente diferenciables en 25 variables morfométricas. Además, los análisis multivariados sugirieron que C. brevicauda, C. perspicillata y C. manu forman un grupo morfológico distinto del grupo de C. benkeithi y Carollia sp. Estos resultados son concordantes con estudios previos y adicionan varios caracteres que diferencian a C. brevicauda de C. perspicillata, los cuales necesitan ser probados en otras regiones.
Palabras clave: caracterización; discriminación; morfología; morfometría; similitud.
Abstract
In the bat genus Carollia, C. brevicauda and C. perspicillata are the most abundant and widely distributed in South America; also, their distributional ranges are almost completely overlapped. Because these species are morphologically very similar, they are frequently misidentified. The aims of this study were to determine the morphological and morphometric differences of C. brevicauda and C. perspicillata. For that reason, I employed a detail morphological assessment and several multivariate morphometric tools. I evaluated 26 normally craniodental variables in 375 adult specimens from Peruvian and Ecuadorian populations. In addition, 54 specimens of C. benkeithi, C. manu, and Carollia sp. were included for comparisons. I found that C. brevicauda and C. perspicillata are differentiated by 28 discrete characters, and are significantly different in 25 morphometric variables. Also, C. brevicauda, C. perspicillata, and C. manu are clustered in a different group from the group of C. benkeithi and Carollia sp. My results complement previous studies adding several characters that confidently differentiate C. brevicauda from C. perspicillata; which needs to be tested in other regions.
Keywords: characterization; discrimination; morphology; morphometry; similarity.
Introducción
Los murciélagos del género Carollia Gray, 1838 de la familia Phyllostomidae, son uno de los más comunes y abundantes del Neotrópico (Cloutier & Thomas 1992, McLellan & Koopman 2008), presentan una amplia distribución que va desde el centro de México hasta el sur de Brasil, y un rango de elevación desde el nivel del mar hasta 2500 m aproximadamente (McLellan & Koopman 2008, Eisenberg & Redford 1999). Actualmente, ocho especies son reconocidas dentro de Carollia: C. benkeithi Solari y Baker 2006; C. brevicauda (Schinz 1821); C. castanea Allen, 1890; C. manu Pacheco, Solari y Velazco 2004; C. monohernandezi Muñoz, Cuartas-Calle y Gonzáles 2004; C. perspicillata (Linnaeus 1758); C. sowelli Baker, Solari y Hoffmann 2002 y C. subrufa (Hahn 1905), además de dos especies candidatas dentro del complejo "castanea" (Solari & Baker 2006, Velazco 2013). Entre estas, C. brevicauda y C. perspicillata son las especies más abundantes y ampliamente distribuidas, además de ser casi totalmente simpátricas (McLellan & Koopman 2008).
Taxonómicamente, Carollia brevicauda y C. perspicillata tienen una historia larga y compleja, relacionada principalmente a determinaciones erróneas y confusiones taxonómicas. En el caso de C. brevicauda, la localidad tipo no fue indicada en la descripción original (Schinz 1821); no obstante, Wied (1826) la delimitó posteriormente a "Fazenda von Coroaba", cerca del Rio do Espírito Santo, Brasil. A pesar de las muchas muestras en colecciones de museos, C. brevicauda no era reconocida como especie plena y era confundida frecuentemente con C. perspicillata o C. subrufa (Hahn 1907, Thomas 1911, Miller 1924) debido a su tamaño y caracteres intermedios, hasta que Pine (1972) la delimitó morfológicamente a partir de la descripción realizada por Karl Koopman del sintipo presente en el Rijkmuseum en Leiden (Nº 17692), considerándola una especie válida y diferente de C. perspicillata y C. subrufa. Mientras que en el caso de perspicillata, Linnaeus (1766) no designó la localidad tipo, pero fue restringida por Tomas (1911) a Surinam como la más probable. Estos y otros aspectos taxonómicos fueron tratados ampliamente por Pine (1972), quien revisó el género a nivel morfológico reconociendo a cuatro especies: C. brevicauda, C. castanea, C. perspicillata y C. subrufa.
Si bien el estudio de Pine (1972) fue un gran aporte, algunas características que propuso resultaron difíciles de aplicar a nivel poblacional (Koopman 1978, Owen et al. 1984, Cháves 1985). Ante esta incertidumbre cualitativa, McLellan (1984) analizó en forma integral a nivel morfométrico las especies diferenciadas por Pine (1972) empleando métodos estadísticos y multivariados, sugiriendo que las especies de Carollia podían ser discriminadas morfométricamente y que la variación interespecífica era mayor a la intraespecífica. Aunque logró diferenciar a Carollia brevicauda y C. perspicillata, las poblaciones de Perú y Ecuador no estuvieron bien representadas, faltando C. brevicauda de Perú y C. perspicillata de Ecuador. Además, tanto Pine (1972) como McLellan (1984) advirtieron sobre la existencia de variación intraespecífica alta dentro de las especies de Carollia que podría dificultar la diferenciación entre algunas de las especies. Posteriormente, diversos autores dieron soporte a ambos estudios probando sus conjuntos de variables cualitativas y cuantitativas en distintas regiones (Owen et al. 1984, Cháves 1985, York & Papes 2007, Zurc & Velazco 2010, Jarrín-V et al. 2010, Murillo-García 2014); sin embargo, algunos reportaron similitud en morfología entre Carollia brevicauda y C. perspicillata en algunas localidades donde ambas ocurren (Koopman 1978, Pine 1972, Cháves 1985, Murillo-García 2014).
A pesar de los alcances morfológicos y morfométricos de diversos estudios, aún resulta complicado distinguir a Carollia brevicauda y C. perspicillata como resultado de la dificultad de encontrar caracteres cualitativos que discriminen consistentemente a ambas especies a lo largo de sus distribuciones (McLellan 1984, York & Papes 2007, Murillo-García 2014). Las claves taxonómicas diferencian a ambas especies, pero basadas en pocos caracteres, como el tamaño del antebrazo, coloración u otros que varían de uno a otro autor (Pine 1972, Cháves 1985, McLellan & Koopman 2008, Diaz et al. 2016).
El presente estudio tiene por objetivos contribuir a la diferenciación de Carollia brevicauda y C. perspicillata a nivel morfológico y morfométrico probando los caracteres propuestos en estudios previos en poblaciones peruanas y ecuatorianas, proveer descripciones detalladas de ambas especies para facilitar futuros estudios morfológicos en otras regiones y realizar comparaciones con otras especies del género co-distribuidas.
Material y métodos
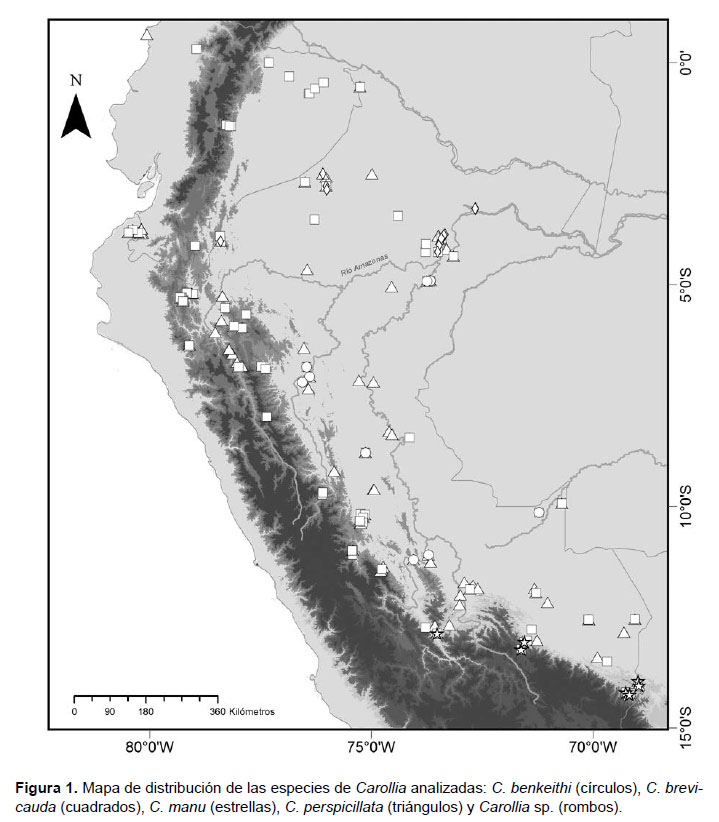
Se analizaron 375 ejemplares adultos entre Carollia brevicauda (n= 185) y C. perspicillata (n= 190), además de otros 54 ejemplares de tres especies de Carollia (C. benkeithi, n=20; C. manu, n= 14 y Carollia sp. n= 20) empleados con finalidad de comparación. Carollia sp. corresponde a las poblaciones del complejo Carollia castanea con distribución al norte del río Amazonas (Solari & Baker 2006). El material analizado se encuentra depositado en el Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MUSM), el Museo de Zoología de la Pontificia Universidad Católica del Ecuador (QCAZ) y el Museo Ecuatoriano de Ciencias Naturales (MECN). En el Apéndice 1 se incluyen detalles de los ejemplares examinados y su distribución geográfica en la Figura 1. Sólo se consideraron datos completos, tomados de especímenes en buen estado. La confirmación de las especies se realizó mediante la revisión de muestras y siguiendo la diagnosis de las especies (Hahn 1907, Pine 1972). La edad se determinó siguiendo a Brunet-Rossinni y Wilkinson (2009), en base al desarrollo de la dentadura y el grado de fusión de la epífisis de los huesos.
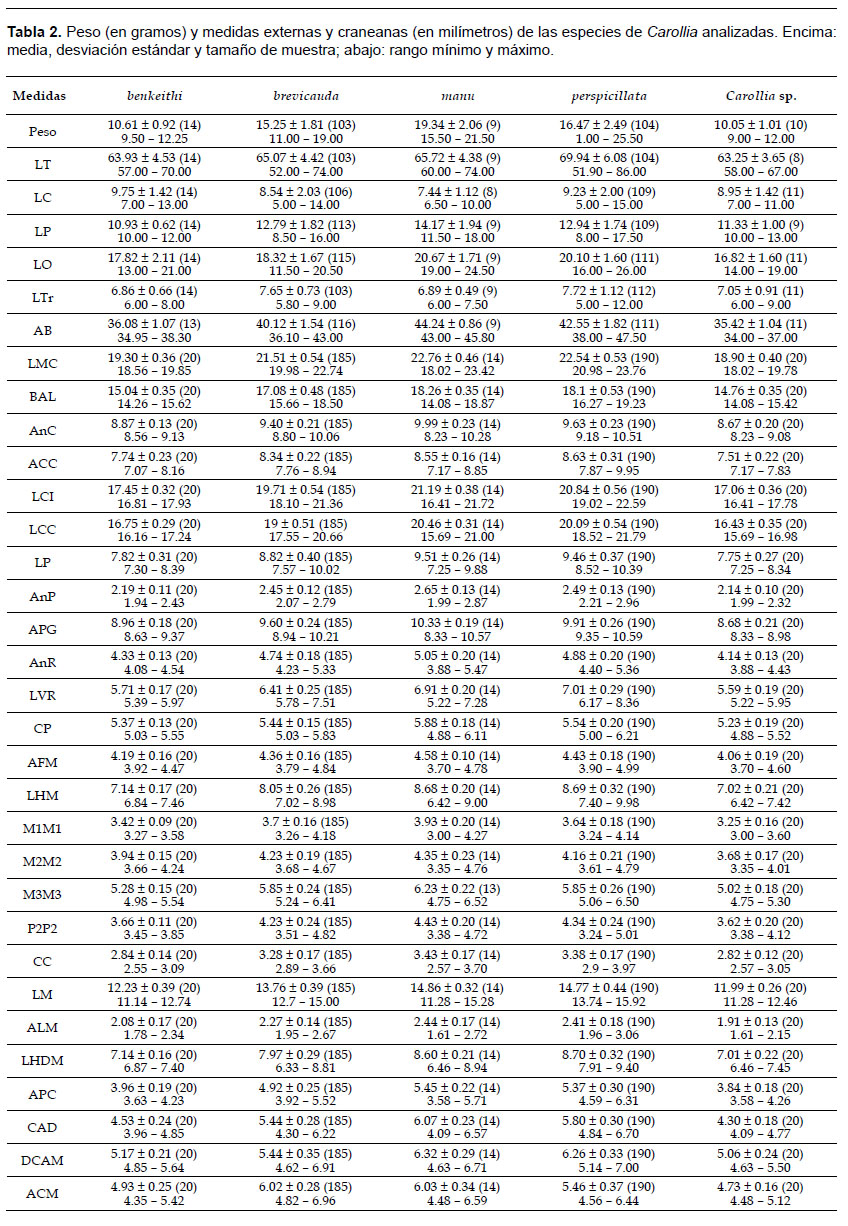
Para la caracterización morfológica se siguió a Wetterer et al. (2000), Velazco (2005), Giannini y Simmons (2007), Fracasso et al. (2011) y Amador et al. (2015). Para definir la coloración del pelaje se siguió la escala de colores de Smithe (1975) bajo luz natural difusa. Se tomaron 26 medidas cráneo-dentales previamente definidas por McLellan (1984) y Zurc y Velazco (2010): longitud máxima del cráneo (LMC), longitud basilar (BAL), ancho máximo de la caja craneal (AnC), altura de la caja craneal (ACC), longitud cóndilo-incisiva (LCI), longitud cóndilo-canina (LCC), longitud palatal (LP), ancho palatal (AnP), ancho postglenoideo (APG), ancho rostral (AnR), longitud ventral rostral (LVR), constricción postorbitaria (CP), ancho del foramen magnum (AFM), longitud de la hilera dental maxilar (LHM), distancia mínima interna entre los primeros molares superiores (M1M1), distancia mínima interna entre los segundos molares superiores (M2M2), distancia mínima interna entre los terceros molares superiores (M3M3), distancia máxima interna entre los segundos premolares superiores (P2P2), distancia máxima interna entre los caninos superiores (CC), longitud mandibular (LM), altura de la mandíbula (ALM), longitud de la hilera dental mandibular (LHDM), altura del coronoide (APC), distancia coronoide-angular (CAD), longitud de la rama postdentaria (DCAM) y ancho máximo condilar (ACM). Las medidas se tomaron con un calibrador Chalimex (±0.01 mm). El peso y las mediciones externas: longitud total (LT), longitud de la cola (LC), longitud de la pata (LP), longitud de la oreja (LO), longitud del trago (Ltr) y longitud del antebrazo (AB), se tomaron directamente de las etiquetas de los individuos con piel preparada y del resto se tomaron de la base de datos de las colecciones revisadas.
Todas las medidas fueron consideradas como variables dependientes; éstas fueron transformadas a logaritmo natural, y se puso a prueba la distribución normal mediante la prueba de Shapiro-Wilk (W). Para probar si existen diferencias significativas entre las especies del género Carollia, se realizó un análisis multivariado de varianza con 1000 permutaciones (PERMANOVA), sobre una matriz de distancias euclidianas con false discovery rate como método de ajuste del valor de p; luego un análisis de varianza de una vía (ANOVA) para cada variable empleando Tukey-Kramer (Lau 2013) como prueba post hoc para muestras de tamaño desigual, con el fin de evaluar cuáles variables eran significativas en la diferenciación de pares de especies. No se consideró la variación entre sexos ya que reportes previos sugieren que su aporte a este nivel es poco significativa (McLellan 1984, York & Papes 2007).
Posteriormente, se realizó un análisis de componentes principales (ACP) para identificar las principales tendencias de variación y un ANOVA del componente que explicó la mayor varianza. Finalmente, se realizó un análisis de función discriminante (AFD) para determinar las variables que permiten discriminar a las especies y se empleó el método de jackknife para evaluar la eficacia del mismo. El valor de confianza para los análisis estadísticos fue de α=0.05. Todos los análisis se realizaron en R 3.4.1 (R Core Team 2017).
Resultados
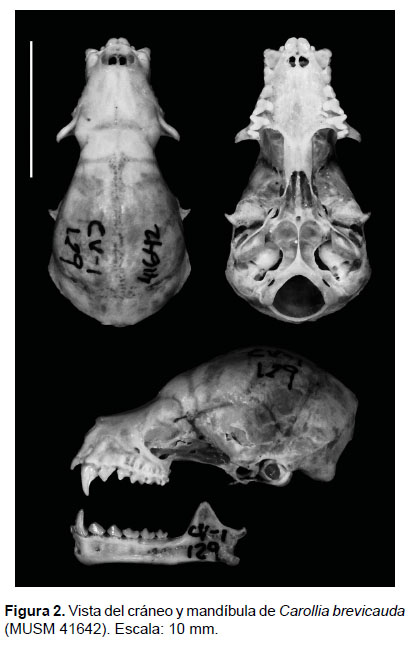
Morfología.-Se presentan y describen un conjunto de caracteres morfológicos cualitativos que permiten separar a Carollia brevicauda de C. perspicillata (Tabla 1), proponiéndose una rediagnosis.
Carollia brevicauda (Schinz, 1821)
"Murciélago de cola corta sedoso"
Rediagnosis: Carollia brevicauda es un murciélago de tamaño mediano que se puede distinguir de sus congéneres mediante la combinación de los siguientes caracteres: pelaje dorsal denso y largo con una longitud entre 7.0 a 8.6 mm y con un patrón tetracoloreado, pelaje del antebrazo denso y corto, tibia mediana, uropatagio relativamente estrecho (referente a la muesca), caja craneal globosa, rostro corto y robusto, pendiente de la frente ligeramente pronunciada, lacrimales hinchados, cresta sagital poco desarrollada, cresta supraoccipital desarrollada, paladar ligeramente curvado a nivel de P2, rama maxilar del cigoma corta y ligeramente robusta, hilera dental maxilar corta y divergente, P1 robusto, maxila curvada ligeramente a nivel del P1-P2, cíngulo posterior del P1 generalmente ausente, cíngulo anterior de P2 desarrollado y orientado internamente, cúspide de P2 1/2 o 2/3 más corta que P1, molares superiores esbeltos y ligeramente espaciados, metastilo de M1 alargado y delgado, protocono de M1 y M2 generalmente esbelto, posprotocrista de M1 cóncava, metastilo de M3 muy pequeño, mandíbula en forma de "U" con ramas divergentes, proceso coronoides mediano, incisivos de la mandíbula no oclusados por los caninos, m3 casi la mitad de ancho que m1.
Descripción (Tabla 1, Fig. 2). Tamaño. Es un murciélago de tamaño mediano entre sus congéneres, con un antebrazo que va de 36 mm hasta 43 mm y una tibia mediana (desde el tobillo hasta la rodilla) que va de 16 a 20 mm. Cabeza. El rostro es corto y ancho, la hoja nasal es pequeña y en forma de diamante con márgenes continuos; presenta numerosas vibrisas mistaciales pequeñas y delgadas, las vibrisas superciliares, suboculares, genales e interramales son muy pequeñas y delgadas, casi del tamaño del pelo y están dispersos; las orejas son pequeñas con bordes continuos de coloración oscura. El trago es pequeño y triangular. Los labios son simples y continuos. El mentón presenta verrugas pequeñas en forma de "V" rodeando a una verruga grande central. Cuerpo. El pelaje dorsal es ligeramente largo (7.0-8.6 mm a nivel de la espalda), los pelos son tretracoloreados: la base es clara, muy angosta y hasta casi imperceptible en algunos individuos, de coloración que va de "pale neutral" (color 86) a "drab-gray" (color 119D), la segunda banda es más amplia y va de "dark grayish" (color 20) a "burnt umber" (color 22), la tercera banda es casi tan amplia como la segunda y va de "pale neutral" (color 86) a "drab-gray" (color 119D), y la punta es generalmente más estrecha, que va de "raw umber" (color 23) a "vandyke brown" (color 121); el pelaje ventral es más corto (4.0-5.0 mm a nivel del pecho) y tricoloreado: la base es oscura y su coloración va de "olive-brown" (color 28) a "light drab" (color 119C), el centro es más claro, "olive-gray" (color 42) a "smoke gray" (color 45); y la punta es más oscura, su coloración va de "olive-brown" (color 28) a "ground cinnamon " (color 239). Los juveniles son de coloración más grisácea. El antebrazo presenta abundante pelo mediano y disperso; el primer dedo tiene la sección brevis poco desarrollada, el dactilopatagio brevis es muy angosto y está insertado entre la falange proximal y media; el plagiopatagio está insertado sobre el tobillo; el uropatagio es poco amplio, con una muesca generalmente poco profunda en forma de "C", el lado dorsal presenta pelos cortos, suaves y muy dispersos. La pata está completamente pigmentada y presenta pelos cortos, suaves y dispersos. El calcar es más pequeño que la pata. La cola es corta (8.54 mm) y está completamente insertada en el uropatagio, y es casi de la mitad de la longitud de ésta. Cráneo. La longitud máxima del cráneo va desde 19.98 mm hasta 22.74 mm. En vista dorsal, el rostro es corto y ligeramente estrecho anteriormente y se ensancha a la altura del segundo premolar, la caja craneana es globosa; la rama maxilar del cigoma es ligeramente poco alargada, divergente y robusta, el arco cigomático está ausente, los lacrimales son hinchados y están bien desarrollados. En vista lateral, la curvatura frontal es pronunciada; la cresta sagital y supraoccipital son poco desarrolladas pero perceptibles al tacto; a nivel del primer y segundo premolar se presenta una curvatura que inclina internamente al segundo premolar, las alas del pterigoides son delgadas, alargadas y dirigidas posteriormente; el cóndilo occipital está bien desarrollado. En vista ventral, el paladar es ligeramente corto y estrecho en su porción anterior y se ensancha a la altura del primer y segundo premolar; el proceso pospalatal es desarrollado, ligeramente ancho, el borde posterior en forma de V o U; las fosas del basioccipital son ligeramente profundas con un septum delgado; el margen anterior del foramen magnum generalmente en forma de C, aunque en algunos individuos es más parecido a una V; el proceso paraoccipital esta pobremente desarrollado; la bula auditiva es lateralmente comprimida y en dirección diagonal. Morfología dental. Fórmula I 2/2, C 1/1, P 2/2, M 3/3, total 34. Superior. Los incisivos centrales (I1) son más grandes y anchos que los externos (I2), tienen puntas romas dirigidas al centro, I2 es muy pequeña, con bordes casi rectos, entre I1 e I2 no hay espacios; el canino es alargado y robusto al inicio, ligeramente curvado hacia atrás, con un cíngulo posterior poco desarrollado; el primer premolar (P1) es alargado y puntiagudo, casi tan ancho en la base como alto, tiene el cíngulo anterior en contacto con el cíngulo posterior del canino; el segundo premolar (P2) es más corto que P1, aproximadamente la mitad de éste (2/3 o 1/2 del alto de P1), el borde anterior del ápice es más corto y recto que el borde posterior, aunque en algunos caso el borde anterior del ápice es tan curvado como el borde posterior, el cíngulo anterior (generalmente desarrollado) no está en contacto con el cíngulo posterior del P1 y está ubicado más internamente que éste, el cíngulo posterolingual está poco desarrollado; el primer molar (M1) es más largo que el segundo molar (M2), el parastilo está generalmente sobre el cíngulo posterior del P2, y el metastilo es largo y delgado y está en contacto con el parastilo de M2, el protocono es ligeramente esbelto y la posprotocrista es ligeramente cóncava; el M2 es más ancho y estrecho que M1, el protocono más estrecho que el protocono de M1; el tercer molar (M3) es la mitad del tamaño que M2, y aparentemente ha perdido el metacono, presenta un paracono bien desarrollado, el parastilo está en contacto con el metastilo de M2. Inferior. El primer incisivo (i1) es más grade y ancho que el segundo (i2), generalmente por lo menos la mitad de alto y ancho, ambos tienen cúspides romas y casi rectas o ligeramente bilobadas, no hay diastemas entre ellos; el canino es ligeramente delgado y puntiagudo, el cíngulo anterior no cubre los incisivos; el primer premolar (p1) es más ancho y ligeramente más alto que el segundo premolar inferior (p2), ambos presentan cúspides romas; el primer molar (m1) es similar en forma y pero ligeramente más estrecho que el segundo molar (m2); el tercer molar (m3) es más estrecho (aproximadamente la mitad de ancho del m1) y bajo pero similar en forma al segundo molar. Mandíbula. En vista oclusal, las ramas mandibulares son divergentes y están dispuestas en forma de "U", el proceso angular es divergente a la rama mandibular. En vista lateral, el proceso coronoides generalmente es alto, el borde anterior es casi recto y más corto que el borde posterior que es más cóncavo; el proceso angular es ligeramente corto y delgado.
Comparación (Figs. 3, 4 y 5, Tablas 1 y 2). Carollia brevicauda comparada con C. benkeithi presenta el pelaje dorsal más largo (8.6 mm versus 6.3 mm a nivel de la espalda) y más denso, el patrón de bandas tetracoloreado, el antebrazo en promedio más largo (40.12 mm versus 36.08 mm) y con pelaje más denso y largo, la tibia más larga, el uropatagio más estrecho, el cráneo más grande (LMC: 21.51 mm versus 19.13 mm), la caja craneal menos globosa, el rostro más robusto, la pendiente de la frente más pronunciada, los lacrimales más hinchados, las crestas sagital y supraoccipital ligeramente más desarrolladas, el paladar curvado a nivel de P1-P2 (en benkeithi a nivel de P2-M1), las fosas de basioccipital ligeramente más profundas, la rama maxilar del cigoma más corta, robusta y más divergente, los caninos y P1 más robustos, el cíngulo anterior de P2 más desarrollado, el metastilo de M1 más alargado, la posprotocrista de M1 menos cóncava, el M1 alineado al borde labial de la hilera dental, las ramas mandibulares más robustas y más divergentes, el proceso coronoides más alto (4.93 mm versus 3.96 mm), el i2 más ancho y alto que i1, el p1 casi del mismo alto que p2, el m1 con cúspides pronunciadas, los protocónidos de m2 y m3 más anchos.
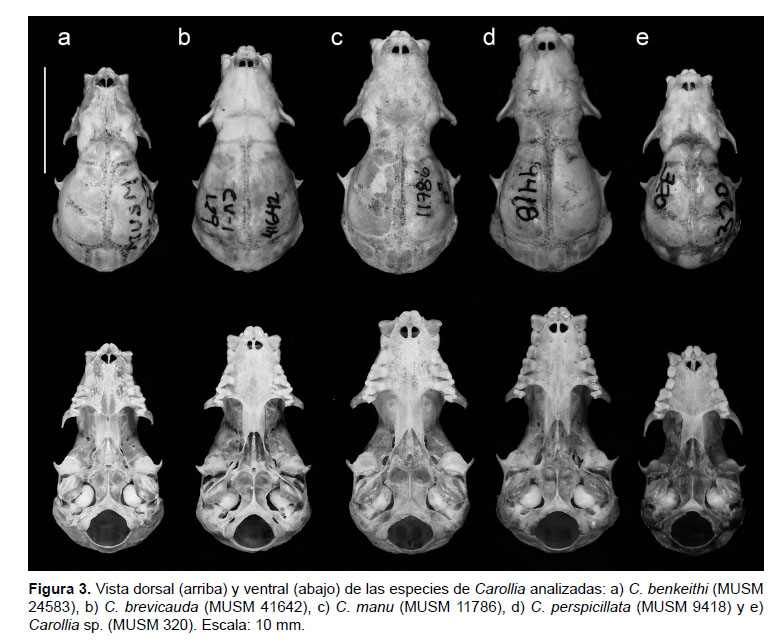
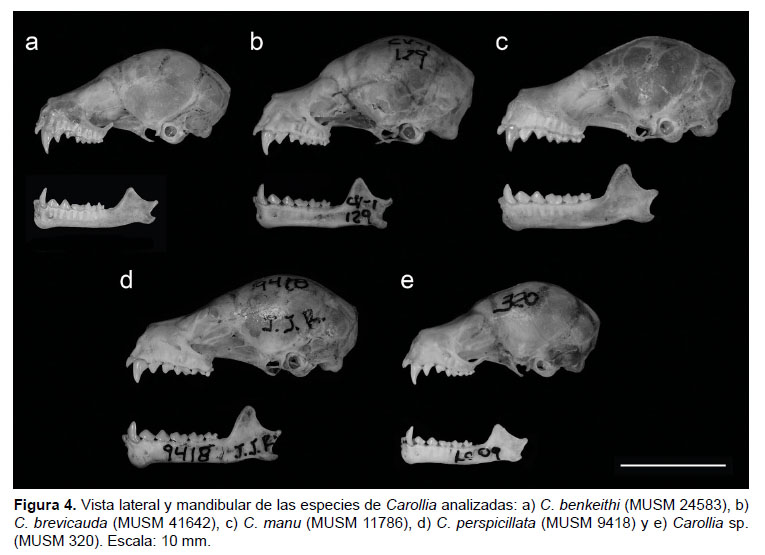
Carollia brevicauda comparada con C. manu presenta el pelaje más corto (8.6 mm versus 10.1 mm a nivel de la espalda) y menos denso, la tercera banda del pelo ligeramente menos marcada, el antebrazo más corto (40.12 mm versus 44.24 mm) con pelaje menos denso y largo, el cráneo más pequeño (LMC: 21.54 mm versus 22.76 mm), el rostro más corto y menos robusto, la pendiente de la frente más pronunciada, los lacrimales menos hinchados, las crestas sagital y supraoccipital menos pronunciadas, el proceso pospalatal ligeramente más corto, las fosas del basioccipital más profundas, la hilera dental maxilar más corta y divergente, los caninos menos robustos, el cíngulo posterior de P1 generalmente poco desarrollado o ausente, el diastema entre P1 y P2 presente, la cúspide de P2 menos alargada, los molares superiores más esbeltos, el M1 mejor alineado al borde labial de la hilera dental, el metastilo de M1 más alargado y delgado, el protocono de M1 más esbelto, la protocrista de M1 más estrecha, el proceso coronoides más bajo (4.92 mm versus 5.45 mm).
Carollia brevicauda comparada con C. perspicillata presenta el pelaje más largo (8.6 mm versus 7.6 mm a nivel de la espalda) y denso, el patrón de bandas tetracoloreado, el antebrazo más corto (40.12 mm versus 42.55) con pelaje más denso y conspicuo, la tibia más corta, el uropatagio más estrecho, el cráneo más pequeño (LMC: 21.51 mm versus 22.54 mm), la caja craneal más globosa, el rostro más corto y robusto, la pendiente de la frente más pronunciada, los lacrimales más hinchados, las crestas sagital y supraoccipital menos desarrolladas, el paladar curvado a nivel del P1-P2, el proceso pospalatal más corto, las fosas del basioccipital generalmente menos profundas, la rama maxilar del cigoma más divergente, la hilera dental maxilar más corta y divergente, la maxila curvada a nivel de P2, el cíngulo anterior de P2 más desarrollado, la cúspide de P2 más corta (1/2 o 2/3 de P1), los molares más esbeltos, el metastilo de M1 más largo y delgado, el protocono de M1 y M2 más esbelto, la protocrista de M1 más estrecha, la mandíbula con ramas más divergentes, el proceso coronoides menos alto (4.93 mm versus 5.37 mm), el i2 no cubierto parcial o totalmente por el cíngulo del canino.
Carollia brevicauda comparada con Carollia sp. presenta pelaje dorsal más largo (8.6 mm versus 7.3 mm a nivel de la espalda) y denso, el patrón de bandas tetracoloreado, antebrazo más largo (40.12 mm versus 35.42 mm) con pelo más denso y largo, la tibia más larga, el uropatagio más estrecho, el cráneo más grande (LMC: 21.51 mm versus 18.90 mm), la caja craneal menos globosa, el rostro más robusto, la pendiente de la frente más pronunciada, los lacrimales más hinchados, la cresta sagital ligeramente más desarrollada, el paladar curvado a nivel de P1-P2 (en benkeithi a nivel de P2-M1), la rama maxilar del cigoma más corta y ancha, los caninos más anchos, el cíngulo anterior de P2 más desarrollado, el M1 alineado al borde labial de la hilera dental, el metastilo de M1 más alargado, la rama mandibular más ancha (2.27 mm versus 1.91 mm) y más divergente, el proceso coronoides más alto (4.92 mm versus
3.84 mm), el i2 más ancho y alto que i1, el m1 con cúspides pronunciadas, el protocónido de m2 y m3 más ancho.
Carollia perspicillata (Linnaeus, 1758)
"Murciélago frutero común"
Rediagnosis: Carollia perspicillata se puede distinguir de sus congéneres mediante la combinación de los siguientes caracteres: tamaño grande, pelaje dosal corto entre 5.0 mm a 7.0 mm, pelos dorsales generalmente tricoloreados con la banda media muy marcada, pelaje del antebrazo disperso y corto, tibia larga, uropatagio amplio, caja craneal ligeramente alargada y poco globosa, rostro alargado y ligeramente robusto, pendiente de la frente poco pronunciada o casi recta, lacrimales poco hinchados, cresta sagital desarrollada, cresta supraoccipital muy desarrollada, paladar casi recto lateralmente, proceso pospalatal ligeramente largo, fosas del basioccipital profundas, rama maxilar del cigoma poco divergente, corta a muy corta y ligeramente robusta, hilera dental maxilar larga y poco divergente, caninos robustos, cíngulo posterior del P1 poco desarrollado, diastema presente entre P1 y P2, cíngulo anterior de P2 poco desarrollado o ausente, cúspide de P2 alargada y casi del mismo alto que P1, molares superiores robustos, M1 alineado al borde labial de la hilera dental, metastilo de M1 alargado y ancho, protocono de M1 y M2 robusto, posprotocrista de M1 ensanchada, mandíbula poco divergente y en forma de "V", rama mandibular robusta, proceso coronoides alto, incisivos de la mandíbula oclusados por los caninos, i2 más del doble de ancho y alto que i1, p1 y p2 casi del mismo tamaño, cúspide de m1 pronunciada, protocónidos de m2 y m3 anchos.
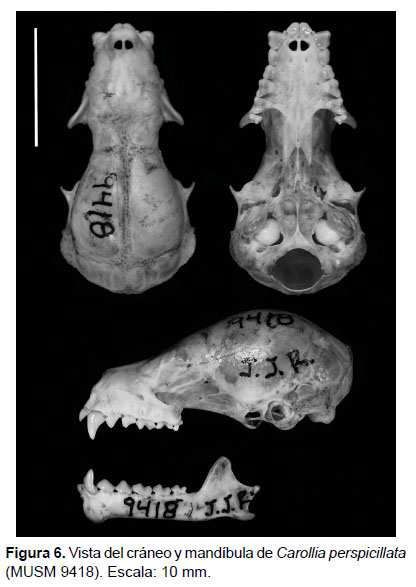
Descripción (Tabla 1, Fig. 6). Tamaño. Es un murciélago de tamaño grande entre sus congéneres, con un antebrazo de 38.00 a 47.50 mm y una tibia de 18.00 a 22.00 mm (desde el tobillo hasta la rodilla). Cabeza. El rostro es ligeramente alargado; la hoja nasal es pequeña y en forma de diamante con márgenes continuos; presenta numerosas vibrisas mistaciales pequeñas y delgadas, las vibrisas superciliares, suboculares, genales e interramales son muy pequeñas y delgadas, casi del tamaño del pelo y están dispersos; las orejas son pequeñas con bordes continuos y de coloración oscura; el trago es pequeño; los labios son simples y continuos; el mentón presenta verrugas pequeñas en forma de "V" rodeando a una verruga grande central. Cuerpo. El pelaje dorsal es ligeramente corto (5.0-7.0 mm), los pelos son generalmente tricoloreados: la base es amplia y su coloración va de "dusky brown" (color 19) a "fuscous" (color 21), y también "mars brown" (color 223A); la banda media es casi tan amplia como la segunda y va de "pale neutral" (color 86) a "cinnamon" (color 123B) y en algunos pocos individuos es "verona brown" (color 223D); y la punta es generalmente más estrecha, que va de una coloración de "army brown" (color 219B) a "vandyke brown" (color 121) y también "tawny" (color 38); cuando el pelo es ligeramente más largo se presenta una banda más basal que es de coloración blanquecina. El pelaje ventral es más corto (3.0-4.0 mm), los pelos son tenuemente tricoloreados: la base es muy amplia y oscura con una coloración "dark grayish brown" (color 20), el centro es "smoke gray" (color 45) o "drab" (color 27) y la punta de color "drab" (color 27) a "fawn color" (color 25); por lo general, la banda media y la punta son poco diferenciables dándole apariencia bicoloreada. Los juveniles son de coloración más grisácea. El antebrazo es comparativamente largo con poco pelo bastante disperso, el primer dedo tiene la sección brevis poco desarrollada, el dactilopatagio brevis es muy angosto y está insertado entre la falange proximal y media. El plagiopatagio está insertado sobre el tobillo. El uropatagio es amplio con una muesca generalmente superficial y generalmente en forma de "V", el lado dorsal presenta pelos cortos, suaves y muy dispersos. La pata está completamente pigmentada y presenta pelos cortos, suaves y dispersos. El calcar es más pequeño que la pata. La cola es corta (9.23 mm) y está completamente insertada en el uropatagio y es casi la mitad de la longitud de ésta. Cráneo. La longitud máxima del cráneo va desde 20.98 mm hasta 23.76 mm. En vista dorsal, el rostro es ligeramente alargado y se ensancha proporcionalmente en forma divergente; la caja craneana es alargada y redondeada; la rama maxilar del cigoma en general no es muy alargada, es robusta, divergente y poco curvada, aunque en algunos individuos la curvatura es más pronunciada; el arco cigomático está ausente; los lacrimales generalmente son poco desarrollados pero evidentes. En vista lateral, la curvatura frontal es poco pronunciada, incluso se muestra casi recta en algunos especímenes; la cresta sagital y supraoccipital son desarrolladas en diferente grado, en general los machos lo tienen ligeramente más desarrollados que las hembras; las alas del pterigoides son delgadas, muy alargadas y dirigidas posteriormente; el cóndilo occipital está bien desarrollado. En vista ventral, el paladar es ligeramente alargado y estrecho en su porción anterior y se ensancha proporcionalmente hasta el tercer molar; el proceso pospalatal es desarrollado, alargado, ligeramente ancho, el borde posterior en forma de "V" o "U"; las fosas del basioccipital son profundas con un septum desarrollado; el margen anterior del foramen magnum generalmente tiene forma de "C", aunque en algunos individuos es más parecido a una "V" con la muesca redondeada; el proceso paraoccipital esta pobremente desarrollado; la bula auditiva está comprimida lateralmente y en dirección diagonal. Morfología dental. Fórmula I 2/2, C 1/1, P 2/2, M 3/3, total 34. Superior. Los incisivos centrales (I1) son más grandes y anchos que los externos (I2), tienen puntas romas dirigidas al centro, I2 es diminuto, con bordes casi rectos, entre I1 e I2 no hay espacios; el canino es alargado y robusto al inicio, ligeramente curvado hacia atrás, con un cíngulo posterior poco desarrollado; el primer premolar (P1) es alargado y puntiagudo, ligeramente más alto que ancho, tiene el cíngulo anterior poco desarrollado ubicado inmediatamente por debajo del cíngulo posterior del canino, en algunos individuos el cíngulo anterior de P1 parece estar ausente; el segundo premolar (P2) es casi tan largo como P1, aproximadamente 3/4 de éste, el borde anterior del ápice es más corto y recto que el borde posterior, el cíngulo anterior, si está desarrollado, generalmente está en contacto con el cíngulo posterior del P1 y ambos se encuentran alineados, el cíngulo posterolingual está poco desarrollado; el primer molar (M1) es ligeramente más ancho que el segundo molar (M2), el parastilo de M1 está generalmente en contacto con el cíngulo posterior del P2 y el metastilo en contacto con el parastilo de M2; el protocono de M1 es robusto y la posprotocrista es ensanchada; el M2 es ligeramente más alargado o casi del mismo tamaño que M1 pero con el protocono más estrecho que el protocono de M1; el tercer molar (M3) es pequeño, casi la mitad de M2 y aparentemente ha perdido el metacono, el paracono está bien desarrollado y el parastilo está generalmente en contacto con el metastilo de M2. Inferior. El primer incisivo (i1) es más grade y ancho que el segundo (i2), siendo i2 por lo menos la tercera parte del primero; ambos incisivos son similares en forma con cúspides bilobadas o casi rectas y no hay espacio entre ellos; el canino es delgado y puntiagudo, el cíngulo anterior generalmente cubre completamente i2; el primer premolar inferior (p1) es similar en forma y tamaño al segundo premolar inferior (p2) y ambos presentan cúspides romas; el primer molar (m1) es similar en forma y tamaño al segundo molar (m2), el protocónido de m1 es más alto que el hipocónido; el tercer molar (m3) es más estrecho (aproximadamente ¾ de m1) y bajo pero similar en form a al segundo molar. Mandíbula. En vista oclusal, las ramas mandibulares están dispuestas en forma de "V" y poco divergentes, el proceso angular es ligeramente divergente, puede ser alargado o no. En vista lateral, el proceso coronoide es generalmente alto, el borde anterior es casi recto y más corto que el borde posterior que es cóncavo. El proceso angular es ligeramente corto y delgado.
Comparación (Figs. 3, 4 y 5, Tablas 1 y 2). Carollia perspicillata comparada con C. benkeithi presenta el pelaje dorsal ligeramente más largo (7.6 mm versus 6.3 mm), patrón de bandeo más marcado, la banda basal del pelo dorsal más ancha, antebrazo más largo (42.55 mm versus 36.08 mm) y ligeramente más peludo, la tibia más larga, el cráneo más grande (LMC: 22.54 mm versus 19.30 mm), la caja craneal menos globosa, el rostro más alargado y robusto, la pendiente de la frente menos pronunciada, las crestas sagital y supraoccipital más desarrolladas, el proceso pospalatal más largo, el borde lateral del paladar casi recto, la rama maxilar del cigoma más corto y robusto, las fosas del basioccipital más profundas, la hilera dental más larga y menos divergente, los caninos más robustos, el borde maxilar casi recto, el P1 más ancho, el cíngulo posterior de P1 más desarrollado, la cúspide de P2 más alargada, los molares superiores más robustos, el M1 mejor alineado al borde labial de la hilera maxilar, el metastilo de M1 más ancho, el protocono de M1 y M2 más robusto, la posprotocrista de M1 más ensanchado, las ramas mandibulares menos divergentes y en forma de "V", la mandíbula más robusta (2.44 mm versus 2.08 mm), el proceso coronoides más alto (5.37 mm versus 3.96 mm), el i2 cubierto parcial o totalmente por el cíngulo de los caninos, el i1 más ancho y alto que i2, el p2 casi del mismo tamaño que p1, el protocónido de m2 y m3 más ancho.
Ver la comparación de Carollia perspicillata con C. brevicauda en la sección de comparaciones de C. brevicauda.
Carollia perspicillata comparada con C. manu presenta el pelaje dorsal más corto (7.6 mm versus 10.1 mm) y menos denso, el patrón de bandas del pelo dorsal generalmente tricoloreado, el antebrazo más corto (42.55 mm versus 44.24 mm) y con pelaje más corto y disperso, la tibia más larga, la caja craneal menos globosa, el rostro ligeramente más delgado, los lacrimales menos hinchados, los bordes laterales del paladar casi rectos y menos divergentes, el borde dental del maxilar casi recto, el cíngulo posterior de P1 menos desarrollado, el diastema entre P1 y P2 presente, el cíngulo anterior de P2 menos desarrollado, los molares superiores menos robustos, el M1 alineado al borde labial de la hilera dental, el metastilo de M1 más delgado, el protocono de M1 y M2 menos robustos, las ramas mandibulares menos divergentes y fusionadas en forma de "V", incisivos inferiores total o parcialmente cubiertos por el cíngulo anterior de los caninos, i1 más ancho y alto que i2.
Carollia perspicillata comparada con Carollia sp. presenta el pelaje dorsal ligeramente más denso, la banda basal del pelo dorsal más ancha y más marcada, el antebrazo más largo (42.55 mm versus 35.42 mm) con pelaje más conspicuo, la tibia más larga, el cráneo más grande (LMC: 22.54 mm versus 18.95 mm), la caja craneal menos globosa, el rostro más alargado, la pendiente de la frente menos pronunciada, las crestas sagital y supraoccipital más desarrolladas, los lacrimales menos hinchados, el borde lateral del paladar casi recto, el proceso pospalatal más largo, la rama maxilar del cigoma más corto y ancho, la hilera dental más larga, los caninos más robustos, el borde maxilar casi recto, la cúspide de P2 más alargada, los molares superiores más robustos, el M1 mejor alineado al borde labial de la hilera maxilar, el metastilo de M1 más ancho, el metacono de M1 y M2 más robusto, las ramas mandibulares menos divergentes y fusionadas en forma de "V", la mandíbula más robusta (2.41 mm versus 1.91 mm), el proceso coronoides más alto (5.37 mm versus 3.84 mm), el i2 cubierto parcial o totalmente por el cíngulo de los caninos, el i1 más ancho y alto que i2, el protocónido de m2 y m3 más ancho.
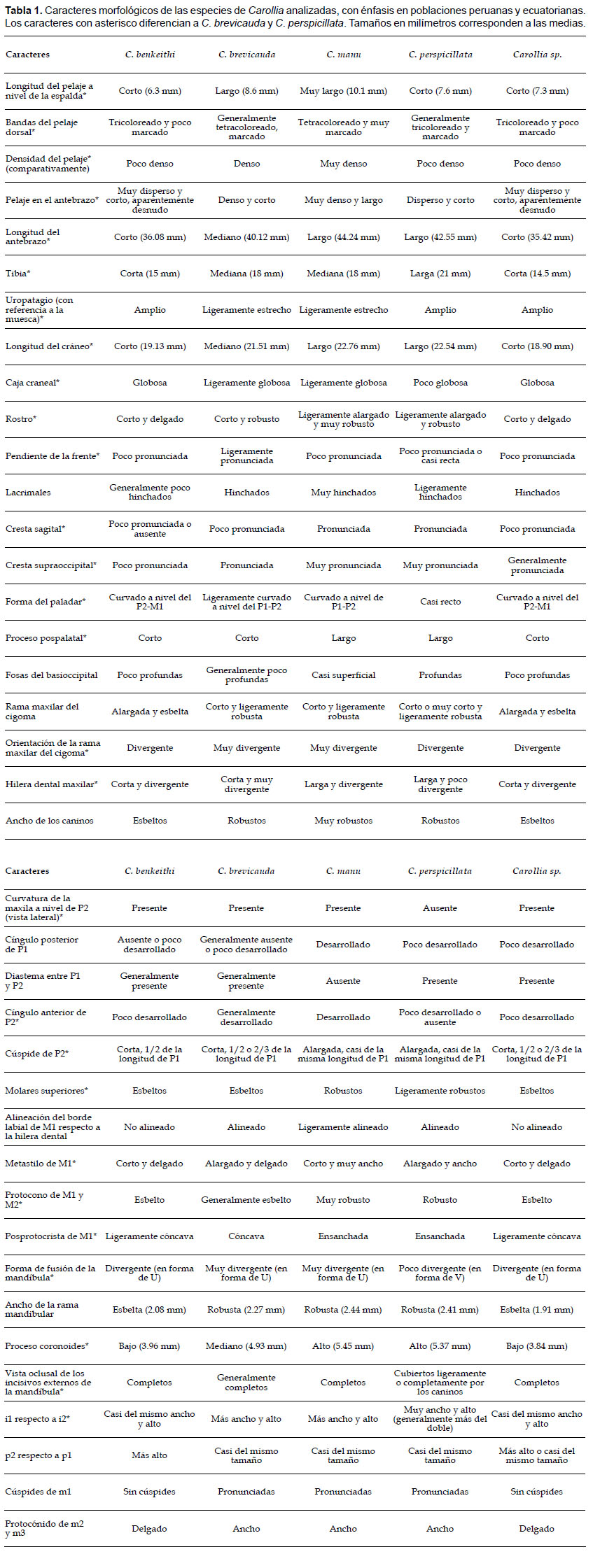
Morfometría. De las especies analizadas, Carollia sp. seguida por C. benkeithi presentaron las medias más bajas en todas las variables, sugiriendo que ambas son las especies más pequeñas, mientras que las medias más altas se presentaron en C. manu (LP, LO, AB, LMC, BAL, AnC, LCI, LCC, LP, AnP, PGW, AnR, CP, AFM, M1M1, M2M2, M3M3, P2P2, CC, LM, ALM, APC, CAD y DCAM) y C. perspicillata (LT, LC, LTr, ACC, LVR, LHM y LHDM) sugiriendo que ambas representan a las especies más grandes, mientras que C. brevicauda se presentó como intermedia entre éstas, excepto en la variable ACM la cual presentó una media similar a C. manu, sugiriendo que C. brevicauda es una especie mediana (Tabla 2).
Todas las variables empleadas tuvieron una distribución normal (p< 0.05) y fueron empleadas en los análisis posteriores: LMC (W= 0.90), BAL (W= 0.91), AnC (W= 0.97), ACC (W= 0.97), LCI (W= 0.90), LCC (W= 0.90), LP (W= 0.96), AnP (W= 0.98), PGW (W= 0.96), AnR (W= 0.97), LVR (W= 0.97), CP (W= 0.99), AFM (W= 0.99), LHM (W= 0.96), M1M1 (W= 0.99), M2M2 (W= 0.99), M3M3 (W= 0.96), P2P2 (W= 0.96), CC (W= 0.96), LM (W= 0.94), ALM (W= 0.99), LHDM (W= 0.97), APC (W= 0.91), CAD (W= 0.91), ACM (W= 0.97) y DCAM (W= 0.96).
Los resultados de PERMANOVA mostraron diferencias significativas entre las especies evaluadas (F= 291.13, pperm<0.05). Además, se encontraron diferencias significativas (p<0.05) a nivel interespecífico para cada una de las variables empleadas: LMC (F= 390.15), BAL (F= 388.43), AnC (F= 158.74), ACC (F= 128.64), LCI (F= 416.44), LCC (F= 444.46), LP (F= 195.19), AnP (F= 67.74), PGW (F= 15.15), AnR (F= 112.50), LVR (F= 273.71), CP (F= 38.36), AFM (F= 32.35), LHM (F= 315.64), M1M1 (F= 49.67), M2M2 (F= 43.92), M3M3 (F= 90.09), P2P2 (F= 78.92), CC (F= 94.49), LM (F= 410.85), ALM (F= 67.39), LHDM (F= 318.20), APC (F= 275.53), CAD (F= 216.80), ACM (F= 178.88) y DCAM (F= 148.35). También se hallaron diferencias significativas (p<0.05) entre pares de especies en la mayoría de comparaciones; sin embargo, no se halló diferencias significativas al analizar las variables: LMC entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, BAL entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, ACC entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, LCI entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, LCC entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, LP entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, AnP entre C. benkeithi-Carollia sp., LVR entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, AFM entre C. benkeithi-Carollia sp., CP entre C. benkeithi-Carollia sp. y C. brevicauda-C. benkeithi, LHM entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, P2P2 entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, CC entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, M2M2 entre C. brevicauda-C. manu, M3M3 entre C. brevicauda-C. perspicillata, LM entre C. benkeithi-Carollia sp., y C. manu-C. perspicillata, ALM entre C. manu-C. perspicillata, LHDM entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, APC entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata, CAD entre C. benkeithi-Carollia sp., ACM entre C. benkeithi-Carollia sp. y C. manu-C. perspicillata y C. brevicauda-C. benkeithi y DCAM entre C. benkeithi-Carollia sp. y C. brevicauda-C. manu
Los tres primeros componentes del ACP explicaron el 81.84% de la varianza total. El componente principal 1 explicó el mayor porcentaje de esta varianza (CP1: 67.38%), los autovectores variaron negativamente y estuvo influenciado principalmente por ALM, APC y CAD (Tabla 3). El componente principal 2 explicó el 10.12% de la varianza y estuvo influenciado principalmente por M2M2, ACM y DCAM, y el Componente principal 3 explicó el 4.34% de la varianza y fue influenciado también por M2M2, ACM y DCAM (Tabla 3). La distribución a lo largo del morfoespacio del CP1 mostró la separación de dos grupos, uno conformado por las especies pequeñas (Carollia benkeithi y Carollia sp.) y otro por las especies más grandes (C. brevicauda, C. manu y C. perspicillata) (Fig. 7a). En el CP1, tanto C. brevicauda como C. perspicillata se muestran ligeramente sobrepuestas y C. manu solapada casi completamente por ambas; mientras que C. benkeithi y Carollia sp. son casi indistinguibles. El resultado del ANOVA del CP1 encontró diferencias significativas entre las especies analizadas (F= 460.59, p<0.05).
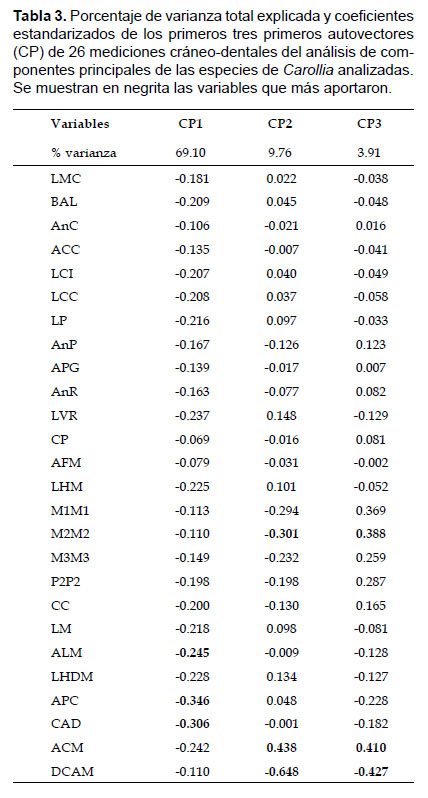
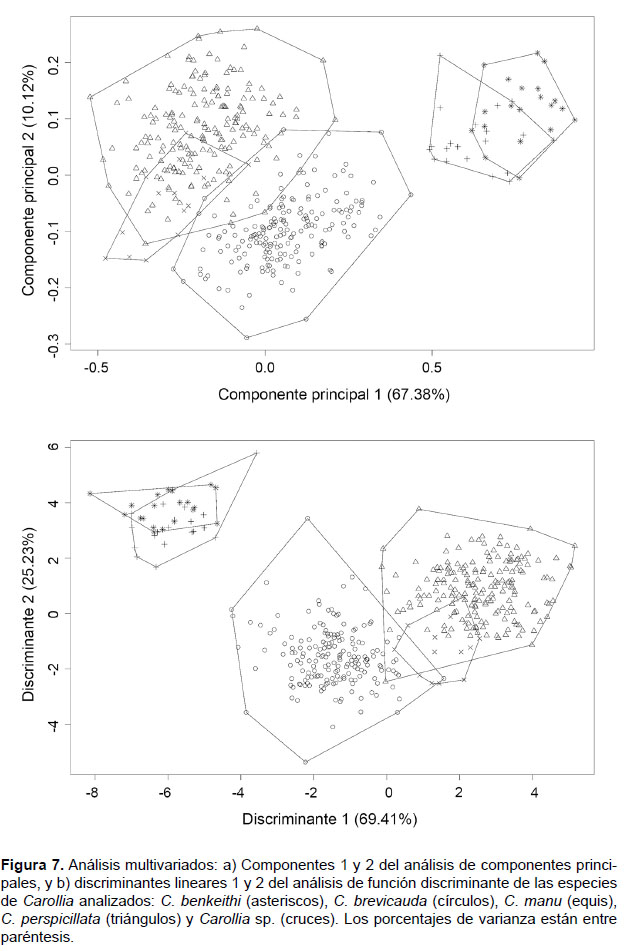
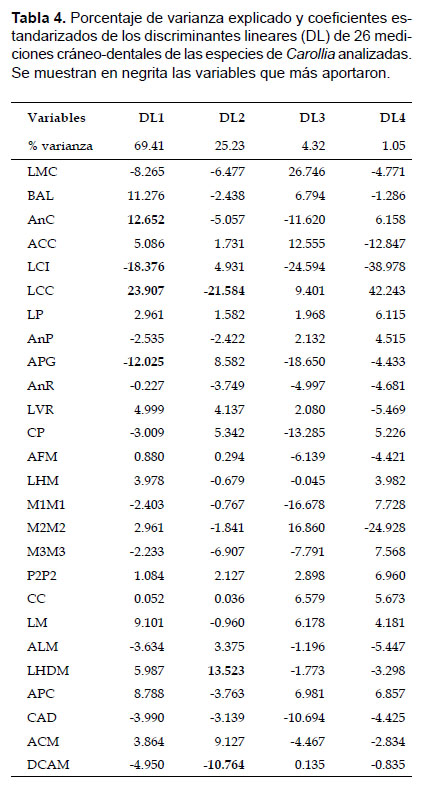
Los resultados del AFD mostraron dos grupos definidos, uno conteniendo a las especies mas grandes, donde Carollia brevicauda se encuentra separada y alejada de C. perspicillata, mientras que C. manu continúa sobrepuesta con ambas especies, y el otro conteniendo a las especies pequeñas mostrando a C. benkeithi y Carollia sp. sobrepuestas casi en su totalidad (Fig. 7b). El discriminante linear 1 (DL1) explicó el mayor porcentaje de éste (69.41%), y las variables que más contribuyeron en la discriminación de las especies fueron AnC, LCI, LCC y PGW; el discriminante linear 2 (DL2) explicó el 25.23% de la varianza, y las variables que más aportaron fueron LCC, LHDM y DCAM; mientras que los discriminantes 3 y 4 (DL3 y DL4) solo explicaron el 5.37% de la varianza total (Tabla 4). El 94.87% de los especímenes fueron correctamente asignados a las especies determinadas a priori (empleando los caracteres propuestos) en función a los cuatro discriminantes lineares, donde C. brevicauda es el más distintivo con 97.84% de especímenes correctamente clasificados, seguido por C. perspicillata con 96.32%, C. manu con 84.71%, Carollia sp. con 80.00% y C. benkeithi con 75.00%. No se registraron especímenes de C. brevicauda clasificados por los discriminantes como C. perspicillata, sin embargo, especímenes de C. perspicillata clasificados como C. brevicauda representaron el 2.11%.
Discusión
Variación morfológica y morfométrica
A pesar de que Carollia brevicauda y C. perspicillata figuran entre los murciélagos más comunes y abundantes en Perú y Ecuador (Hice et al. 2004, Pacheco et al. 2007, Arias et al. 2016, Medina et al. 2015), existe aún mucha ambigüedad sobre su delimitación, conllevando a identificaciones erróneas, basadas principalmente en morfología externa y en consecuencia, afectan directamente los análisis de diversidad y abundancia en los que son empleados.
De acuerdo con los resultados de este estudio, Carollia brevicauda y C. perspicillata son discriminadas mediante los análisis morfológicos y morfométricos empleados, dando soporte a lo reportado por otros autores (Pine 1972, McLellan 1984, York & Papes 2007, Zurc & Velasco 2010, Murillo-García 2014) quienes también las diferenciaron empleando estas aproximaciones, y difiriendo de Cháves (1985) quien analizó poblaciones colombianas de Carollia y sugirió que los caracteres morfológicos de Pine (1985) eran imposibles de aplicar y que morfométricamente C. brevicauda y C. perspicillata se sobreponían ampliamente. En el presente trabajo, 28 caracteres externos y cráneo-dentales permitieron distinguir a Carollia brevicauda de C. perspicillata, estos incluyen algunos presentados por primera vez y otros reportados anteriormente (Pine 1972, McLellan & Koopman 2008, Zurc & Velazco 2010) y confirmados en este estudio.
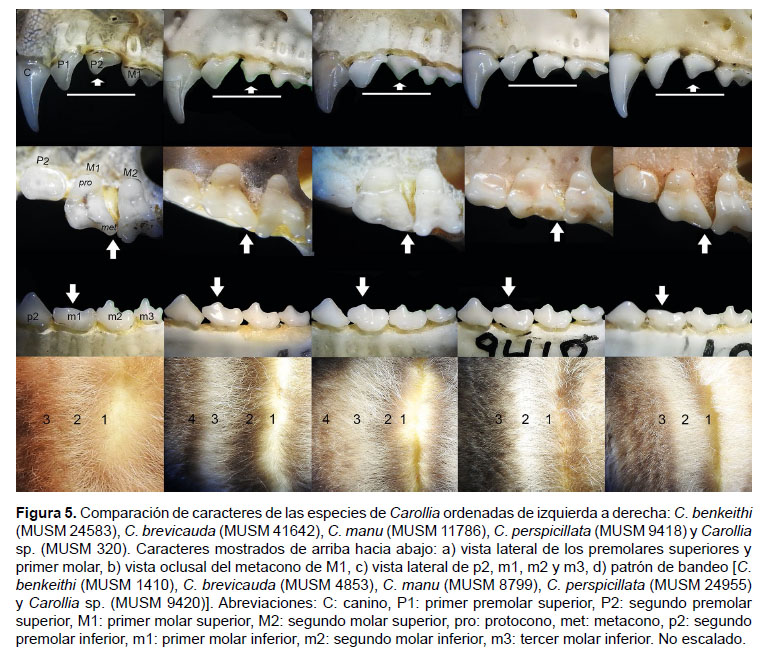
Con respecto a la coloración del pelaje dorsal, Carollia sp. y C. benkeithi son las más diferenciables de sus congéneres por presentar diversas tonalidades de castaño a rojizo y un patrón de bandas poco marcado, mientras que C. brevicauda, C. manu y C. perspicillata presentan patrones de coloración más parecidos y generalmente en tonalidades marrones a grises. Además, se considera al igual que Pine (1972) que el tamaño del pelo es un carácter relevante, ya que en general, C. brevicauda tiene el pelo más largo que C. perspicillata en las localidades donde se les ha registrado en simpatría. El pelo disperso en el antebrazo es un carácter que puede variar individualmente, ya que se ha notado que algunos especímenes de C. brevicauda presentan pelo casi de forma similar que C. perspicillata, pero en general, C. brevicauda presenta el pelo más denso y visible que C. perspicillata. El pelo disperso en las patas y uropatagio de C. brevicauda puede variar en tamaño y visibilidad, mientras que en C. perspicillata es menos evidente; sin embargo, en C. manu este es un carácter indiscutible y diagnóstico.
Con respecto al patrón de bandas del pelaje dorsal, en este estudio se encontró que tanto Carollia brevicauda como C. manu presentan pelos tetracoloreados como patrón más frecuente (95 y 100% de casos respectivamente), contrario a otros estudios que sugieren un patrón tricoloreado para éstas (Cuartas et al. 2001, Baker et al. 2002, Pacheco et al. 2004, Zurc & Velazco 2010); mientras que C. perspicillata presentó generalmente un patrón tricoloreado (90% de casos) y raramente una diminuta banda basal adicional. Con respecto a C. benkeithi y Carollia sp., se coincide con Solari y Baker (2006) que reportan un patrón tricoloreado tenue (85% de casos).
La curvatura del paladar a nivel de P2 fue muy conspicua en más del 85% de los especímenes de Carollia brevicauda y se considera como un carácter diagnóstico, ya que en C. perspicillata, el cíngulo posterior de P1 y el cíngulo anterior de P2 están mejor alineados (90% de los casos) dando al paladar una apariencia casi recta en los bordes laterales (Fig. 5). Esta curvatura del paladar a nivel de P2 es también muy evidente en C. manu, y en menor medida en C. benkeithi y Carollia sp. Contrariamente, Jarrín-V et al. (2010), en un análisis de morfometría geométrica, sugirieron que este carácter es subjetivo entre C. brevicauda y C. perspicillata.
Los caracteres encontrados por Zurc y Velazco (2010) para la distinción de ambas especies no fueron constantes en todos los especímenes analizados. Tanto en Carollia brevicauda y C. perspicillata la forma de la fosa mesopterigoidea varía entre "U" a "V", el metastilo del tercer molar superior es muy pequeño y en algunos casos parece ausente, el proceso paracondilar está desarrollado en diferente grado pero no ausente, generalmente el trigónido tiene la misma proporción que el talónido en el primer molar inferior, la cúspide de primer molar inferior es desarrollada en diferente grado y propensa al desgaste y la alineación del tercer molar inferior respecto al eje longitudinal de la hilera dental en C. perspicillata es muy sutil.
Si bien ambas especies presentan algunos caracteres morfológicos en común, se les puede diferenciar mediante la combinación de los caracteres propuestos en este estudio. Se ha notado que los caracteres cualitativos menos variables que permiten distinguir a ambas especies son la longitud del pelaje dorsal, la longitud de la tibia, el largo del rostro, la curvatura de la frente, la longitud del proceso pospatal, la forma, orientación y tamaño relativo del segundo premolar superior, el desarrollo del metastilo, protocono y posprotocrista de primer molar superior, la divergencia de las hileras maxilares y mandibulares y la oclusión de segundo incisivo inferior por el canino (Tabla 1). También se ha notado un mayor desarrollo de las crestas de los molares inferiores en C. perspicillata respecto a C. brevicauda; sin embargo, este carácter es susceptible de desgaste y debido a ello no se puede evidenciar en todos los especímenes.
Los análisis morfométricos, en general, diferenciaron estadísticamente a las especies con las variables empleadas coincidiendo con otros estudios (McLellan 1984, York & Papes 2007, Zurc & Velazco 2010, Murillo-García 2014). Los resultados del análisis de función discriminante sugieren que el ancho del cráneo, la longitud cóndilo-incisivo, la longitud cóndilo-canino, el ancho postglenoideo, la longitud de la hilera dental mandibular y la longitud de la rama postdentaria permiten discriminar a las cinco especies. Mientras que McLellan (1984) encontró que la longitud basilar, la longitud de la hilera dental maxilar, la longitud ventral del rostro, distancia coronoides-angular, la longitud de la mandíbula y la altura del coronoides eran las variables más útiles para discriminar a C. brevicauda (incluyendo a C. sowelli), C. perspicillata, C. subrufa y C. castanea (incluyendo a Carollia benkeithi). Por otro lado, Cháves (1985) encontró que las variables útiles en la discriminación de ambas especies eran las medidas de las series dentales maxilar y mandibular.
Los análisis multivariados recuperaron un patrón comparable al encontrado por Ruelas (2014) empleando morfometría geométrica, donde las especies grandes y medianas (Carollia brevicauda, C. manu y C. perspicillata) forman un grupo ligeramente diferenciable del de las pequeñas (C. benkeithi y Carollia sp.) y con un soporte estadístico significativo (Fig. 7). Si bien Pacheco et al. (2004) sugirieron que C. manu era la especie más grande y más parecida a C. brevicauda, en este estudio sugerimos que morfológicamente esta especie es intermedia entre C. brevicauda y C. perspicillata, dado que, con respecto al tamaño del cráneo y mandíbula es más similar a C. perspicillata, coincidiendo en esto con Ruelas (2014), mientras externamente C. manu es más parecida a C. brevicauda; no obstante, C. manu comparte algunos caracteres morfológicos con ambas especies (Tabla 1). En relación con otros estudios morfométricos, se encontró esta misma coincidencia en la distinción de las especies grandes de las pequeñas (York & Papes 2007, Jarrín-V et al. 2010, López-Aguirre et al. 2015).
Este estudio también sugiere que de las 26 variables craneodentales empleadas, 25 son significativamente diferentes entre C. brevicauda y C. perspicillata y pueden ser empleados en otros estudios. Sin embargo, aunque Carollia perspicillata, en general, es más grande y de coloración más oscura que C. brevicauda, usar solo estos aspectos como clave en la determinación conlleva a errores frecuentes, ya que individuos grandes y más oscuros de C. brevicauda pueden confundirse y determinarse como individuos pequeños de C. perspicillata. En consecuencia, se ha notado que es más frecuente confundir a C. brevicauda con C. perspicillata que viceversa, principalmente en poblaciones de la vertiente oriental, mientras que en poblaciones occidentales es más frecuente confundir a C. perspicillata con C. brevicauda, así por ejemplo, del total de muestras empleadas, 17 especímenes de C. perspicillata fueron reidentificadas como C. brevicauda y 25 de C. brevicauda fueron reidentificados como C. perspicillata, 12 de los cuales fueron de Tumbes (occidente de Perú). Adicionalmente, de las muestras que no se emplearon en este estudio debido a que se encontraban en mal estado o correspondían a individuos juveniles, cinco fueron reidentificadas como C. brevicauda y 34 como C. perspicillata, todas de la vertiente oriental de Perú.
Las claves taxonómicas generalmente distinguen a Carollia brevicauda y C. perspicillata de acuerdo a la medida del antebrazo, si es mayor de 39.0 o 40.0 mm corresponde a C. perspicillata, pero si es menor, a C. brevicauda (McLellan & Koopman 2008, Diaz et al. 2016); sin embargo, no es raro encontrar especímenes de C. brevicauda con un antebrazo de hasta 43 mm de longitud (Tabla 1); por lo que trabajos que han usado solo el tamaño del antebrazo para diferenciar especies pueden producir resultados equívocos. Aunque los resultados morfométricos en este trabajo sugieren que el tamaño, en general, es un factor importante en la determinación de las especies de Carollia, no es preciso en todos los casos debido a la sobreposición de éstas en los límites inferiores y superiores entre las especies pequeñas (C. benkeithi y Carollia sp.) y entre las medianas y grandes (C. brevicauda, C. manu y C. perspicillata) (Tabla 2).
Si bien en este estudio se notó que las variaciones morfológicas son generalmente individuales, se sugiere realizar un estudio enfocado en este tópico que permita dilucidar, si hubiere, un patrón de variación relacionado a aspectos como la geografía, clinas, o tipos de ecosistemas. Se sugiere también realizar descripciones detalladas, a nivel local, de otras especies de amplia distribución como Desmodus rotundus, Diaemus youngii, Glossophaga soricina o Chrotopterus auritus, entre otros, cuando no se tenga disponibilidad de muestras de todo su rango de distribución, a fin de permitir a otros investigadores hacer comparaciones y analizar los patrones de variación morfológica y morfométrica.
Distribución.
La revisión de especímenes permitió una mejor delimitación de la distribución de ambas especies. En Perú y Ecuador, Carollia brevicauda y C. perspicillata son simpátricos en casi toda su distribución, abarcando distintos tipos de bosques húmedos y secos tanto en el oriente y occidente de los Andes, incluyendo las Yungas, y un rango de elevación desde 27 m (Tumbes, MUSM 20633) hasta 2407 m (Cajamarca, MUSM 25024) para C. brevicauda, y desde 27 m (Tumbes, MUSM 20635) hasta 2200 m (Cajamarca, MUSM 40303) para C. perspicillata.
En el occidente, el límite más austral para Carollia brevicauda y C. perspicillata se ha reportado en el bosque seco de Tumbes (Pacheco et al. 2007); sin embargo, nuestra revisión da cuenta que de 16 especímenes determinados como Carollia brevicauda de Tumbes, 12 correspondían a C. perspicillata, y varios de los especímenes del lado occidental de Ecuador, no empleados en este estudio, también fueron reidentificados como C. perspicillata (aproximadamente 30 especímenes). Esto sugiere que probablemente, el registro de C. brevicauda al occidente de los Andes, entre Perú y Ecuador, es menos frecuente que lo que se ha reportado (Pacheco et al. 2007, Novoa et al. 2011) debido a determinaciones erróneas de especímenes de C. perspicillata como C. brevicauda. Debido a ello se sugiere revisar los especímenes registrados en localidades occidentales a fin de confirmar las determinaciones y realizar un análisis poblacional de ambas especies y analizar el rango de variación geográfica de las poblaciones occidentales no tratado en este estudio.
Situación taxonómica.
Diversos estudios moleculares se han desarrollado con la finalidad de dilucidar las relaciones filogenéticas entre las especies de Carollia. Así, por ejemplo, Wright et al. (1999), usando el gen mitocondrial citocromo b, sugirió que C. brevicauda y C. perspicillata eran especies hermanas y el clado de divergencia más reciente del género, calculado en 1.036 millones de años (Pavan 2011), contradiciendo la hipótesis de Lim y Engstrom (1998), quienes usando enzimas de restricción mitocondrial sugirieron que C. perspicillata era grupo hermano de C. subrufa. En general, la hipótesis más aceptada es que C. brevicauda y C. perspicillata son monofiléticas recíprocas (Wright et al. 1999, Ditchfield 2000, Hoffmann & Baker 2003, Pavan et al. 2011, Velazco 2013).
Aunque sus complejas historias taxonómicas hayan ocasionado anteriormente inconsistencias en las delimitaciones de Carollia brevicauda y C. perspicillata, en la actualidad ambas son consideradas especies plenas, no obstante, presentan un cariotipo similar (2n= 20, FN= 36 [Noguera-Urbano & Muñoz-Montenegro 2014] y 2n= 20-21, NF= 36 [Baker & Bleier 1971] respectivamente) y una distancia genética estimada entre 3.67% (Hoffmann & Baker 2003) a 3.75% (Velazco 2013) entre haplotipos del gen mitocondrial citocromo b. Si bien la distancia genética es menor al 5%, valor propuesto por Baker y Bradley (2006) para la diferenciación entre especies, éste aún está dentro del rango mínimo interespecífico reportado por Bradley y Baker (2001). Otras especies de murciélagos filostómidos con una distancia genética menor al 5% pero morfológicamente diferenciables son Platyrrhinus angustirostris y P. fusciventris con 2.03% (Velazco et al. 2010), Platyrrhinus helleri y P. matapalensis con 3.1% (Velazco et al. 2010), Platyrrhinus albericoi y P. vittatus con 1.2% (Velazco & Patterson 2008), Chiroderma doriae y C. trinitatum con 3.2% (Baker et al. 1994), Chiroderma improvisum y C. trinitatum con 4.1% (Baker et al. 1994).
Agradecimientos
Agradezco profundamente a Víctor Pacheco (MUSM, Museo de Historia Natural UNMSM), Santiago Burneo (QCAZ, Museo de Zoología de la Pontificia Universidad Católica del Ecuador) y Jorge Brito (MECN, Museo Ecuatoriano de Ciencias Naturales) por permitirme acceder a los especímenes de las Colecciones científicas de las que son responsables. A Víctor Pacheco, Natali Hurtado y Richard Cadenillas por sus comentarios y sugerencias. Este trabajo fue financiado por el Programa Nacional de Innovación para la Competitividad yProductividad (INNÓVATE PERÚ), de acuerdo con el contrato 402-PNICP-PIBA-2014.
Literatura citada
Amador L.I, V. Abdala & N.P. Giannini. 2015. Homology of thechiroptera "dactylopatagium" brevis. Mammalian biology 80(6):447-450. doi: http://dx.doi.org/10.1016/j.mambio.2015.06.003 [ Links ]
Arias E., V. Pacheco, K. Cervantes, A. Aguilar & J. Álvarez. 2016.Diversidad y composición de murciélagos en los bosquesmontanos del Santuario Nacional Pampa Hermosa, Junín,Perú. Revista peruana de biología 23(2):103-116. doi:http://dx.doi.org/10.15381/rpb.v23i2.12381 [ Links ]
Baker R.J., V.A. Taddei, J.L. Hudgeons & R.A. Van Den Bussche.1994. Systematic relationships within Chiroderma (Chiroptera: Phyllostomidae) based on cytochrome b sequencevariation. Journal of mammalogy 75(2):321-327. doi:http://dx.doi.org/10.2307/1382550 [ Links ]
Baker R.J., S. Solari & F.G. Hoffmann. 2002. A new Central American species from de Carollia brevicauda Complex. Occasional Papers, Museum of Texas Tech University 217:1-11. [ Links ]
Baker R.J. & R.D. Bradley. 2006. Speciation in mammals and thegenetic species concept. Journal of Mammalogy 87(4):643-662. doi: http://dx.doi.org/10.1644/06-MAMM-F038R2.1 [ Links ]
Baker R.J. & W.J. Bleier. 1971. Karyotypes of bats of the subfamilyCarollinae (Mammalia: Phyllostomatidae) and their evolutionary implications. Experientia 27:220-222. [ Links ]
Bradley R.D. & R.J. Baker. 2001. A test of the genetic speciesconcept: cytochrome-b sequences and mammals. Journal of Mammalogy 82:960-973. doi: http://dx.doi.org/10.1644/1545-1542(2001)082<0960:ATOTGS>2.0.CO;2 [ Links ]
Brunet-Rossinni A.K. & G.S. Wilkinson. 2009. Methods for AgeEstimation and the Study of Senescence in Bats. En: Kunz, T.H. & S. Parsons, eds. Ecological and behavioral methodsfor the study of bats. Johns Hopkins University Press,Baltimore, Maryland. pp. 315-325. [ Links ]
Cháves J.G. 1985. Morfometría y sistemática de murciélagos delgénero Carollia Gray, 1838 (Phyllostomidae, Carolliinae).Tesis de licenciatura inédita, Universidad Nacional de Colombia. Bogotá, Colombia. [ Links ]
Cloutier D. & D.W. Thomas. 1992. Carollia perspicillata. Mammalian species 417:1-9. [ Links ]
Cuartas C.A., J. Muñoz & M. González. 2001. Una nueva especiede Carollia Gray, 1838 (Chiroptera: Phyllostomidae) deColombia. Actualidades biológicas 23(75):63-73. [ Links ]
Díaz M.M., S. Solari, L.F. Aguirre, L.M.S. Aguiar & R. Barquez.2016. Clave de identificación de los murciélagos de Sudamérica. Publicación especial N°2, PCMA (Programa deConservación de los Murciélagos de Argentina). Argentina. [ Links ]
Ditchfield A.D. 2000. The comparative phylogeography of Neotropical mammals: patterns of intraespecific mitochondrialDNA variation among bats contrasted to nonvolant smallmammals. Molecular ecology 9(9):1307-1318. doi: http://dx.doi.org/10.1046/j.1365-294x.2000.01013.x [ Links ]
Eisenberg J.F. & K.H. Redford. 1999. Mammals of the Neotropics:the central Neotropics. Ecuador, Peru, Bolivia, Brazil.University of Chicago Press. pp. 609. [ Links ]
Fracasso M.P., L. Salles & F. Perini. 2011. Upper molar morphology and relationships among higher taxa in bats. Journal of mammalogy 92(2):421-432. doi: http://dx.doi.org/10.1644/09-MAMM-A-415.1 [ Links ]
Giannini N.P. & N.B. Simmons. 2007. The chiropteran premaxilla:A reanalysis of morphological variation and its phylogeneticinterpretation. American Museum Novitates 3585:1-44. [ Links ]
Hahn W.L. 1907. A review of the bats of genus Hemiderma. Proceedings of the United States National Museum 32:103-118. [ Links ]
Hice C.L., P.M. Velazco & M.R. Willig. 2004. Bats of the Reserva Nacional Allpahuayo-Mishana, northeastern Peru, with noteson community structure. Acta chiropterologica 6(2):319-334. doi: http://dx.doi.org/10.3161/001.006.0210 [ Links ]
Hoffmann F.G. & R.J. Baker. 2003. Comparative phylogeography of short-tailed fruit bats (Carollia: Phyllostomidae). Molecular Ecology 12(12):3403-3414. doi: http://dx.doi.org/10.1046/j.1365-294X.2003.02009.x [ Links ]
Jarrín-V P., C. Flores & J. Salcedo. 2010. Morphological variationin the short-tailed fruit bat (Carollia) in Ecuador, withcomments on the practical and philosophical aspects ofboundaries among species. Integrative Zoology 5:226-240.doi: http://dx.doi.org/10.1111/j.1749-4877.2010.00208.x [ Links ]
Koopman K.F. 1978. Zoogeography of Peruvian bats with specialemphasis on the role of the Andes. American MuseumNovitates 2651:1-33. [ Links ]
Lau M.K. 2013. DTK: Dunnett-Tukey-Kramer pairwise multiplecomparison test adjusted for unequal variances and unequalsample sizes. http://CRAN.R-project.org/package=DTK [ Links ]
Lim B.K. & M.D. Engstrom. 1998. Phylogeny of Neotropical shorttailed fruit bats, Carollia spp. Phylogenetic analysis ofrestriction site variation in mtDNA. En: Kunz T.H. & P.A. Racey, eds. Bat biology and conservation, ed. Washington, DC: The Smithsonian Institution Press. pp. 43−58.
López-Aguirre C., J. Pérez-Torres & L.A.B. Wilson. 2015. Cranial andmandibular shape variation in the genus Carollia (Mammalia: Chiroptera) from Colombia: biogeographic patterns and morphological modularity. PeerJ 3:e1197. doi: http://dx.doi.org/10.7717/peerj.1197 [ Links ]
McLellan L.J. & K.F. Koopman. 2008. Subfamily Carolliinae Miller, 1924. En: Gardner A.L., ed. Mammals of South America, Volume 1 Marsupials, Xenarthrans, shrews, and Bats.Chicago and London: The University of Chicago Press.pp. 208-218. [ Links ]
McLellan L.J. 1984. A morphometric analysis of Carollia (Mammalia,Chiroptera). American Museum Novitates 2791:1-35. [ Links ]
Medina C.E., E. López, K. Pino, A. Pari & H. Zeballos. 2015. Biodiversidad de la Zona Reservada Sierra del Divisor (Perú): una visión desde los mamíferos pequeños. Revista peruana de biología 22(2):199-212. doi: http://dx.doi.org/10.15381/rpb.v22i2.11354 [ Links ]
Miller G.S. Jr. 1924. List of North American recent mammals. 1923. Bulletin of the United States National Museum 128:1-673. [ Links ]
Murillo-García O.E. 2014. Murciélagos de cola corta (Carollia:Phyllostomidae) del Parque Nacional Natural Gorgona(Colombia) y sus implicaciones biogeográficas. Revista de Biología Tropical 62(1):435-445. [ Links ]
Noguera-Urbano E.A. & S. Muñoz-Montenegro. 2014. Un cariotipo del murciélago sedoso de cola corta (Carollia brevicauda[Schinz, 1821], Chiroptera: Phyllostomidae) de los andes de Colombia. Therya 5(2):559-566. doi: http://dx.doi.org/10.12933/therya-14-145 [ Links ]
Novoa S., R. Cadenillas & V. Pacheco. 2011. Dispersión de semillas por murciélagos frugívoros en bosques del Parque Nacional Cerros de Amotape, Tumbes, Perú. Mastozoología Neotropical 18(1):81-93. [ Links ]
Owen J.G., D.J. Schmidly & W.B. Davis. 1984. A morphometric analysis of three species of Carollia (Chiroptera, Glossophaginae) from Middle America. Mammalia 48(1):85-93. doi: http://dx.doi.org/10.1515/mamm.1984.48.1.85 [ Links ]
Pacheco V., S. Solari & P.M. Velazco. 2004. A New Species of Carollia (Chiroptera: Phyllostomidae) from the Andes of Peru and Bolivia. Occasional Papers, Museum of Texas Tech University 236:1-15. [ Links ]
Pacheco V., R. Cadenillas, S. Velazco, E. Salas & U. Fajardo. 2007. Noteworthy bat records from the Pacific Tropical rainforest region and adjacent dry forest in northwestern Peru. Acta Chiropterologica 9(2):409-422. doi: http://dx.doi.org/10.3161/1733-5329(2007)9[409:NBRFTP]2.0.CO;2 [ Links ]
Pavan A.C., F. Martins, F.R. Santos, A. Ditchfield & R.A.F. Redondo. 2011. Patterns of diversification in two species of shorttailed bats (Carollia Gray, 1838): the effects of historical fragmentation of Brazilian rainforests. Biological journalof the Linnean Society 102(3):527-539. doi: http://dx.doi.org/10.1111/j.1095-8312.2010.01601.x [ Links ]
Pine R.H. 1972. The bats of the genus Carollia. Technical Monograph, Texas Agricultural Experimental Station, Texas A & M University 8:1-125. [ Links ]
Ruelas D. 2014. Análisis morfo-geométrico del género Carollia Gray, 1838 (Phyllostomidae: Chiroptera) en el Perú. Tesis para para optar por el Título Profesional de Biólogo. Facultad de Ciencias Biológicas, Universidad Nacional de San Agustín. [ Links ]
Smithe F.B. 1975. Naturalist’s color guide. New York: AmericanMuseum of Natural History.
Solari S. & R.J. Baker. 2006. Mitochondrial DNA sequence, karyotypicand morphological variation in the Carollia castanea speciescomplex (Chiroptera: Phyllostomidae) with description of anew species. Occasional Paper The Museum of Texas Tech University 254:1-16. [ Links ]
Thomas O. 1911. The Mammals of the Tenth Edition of Linnæeus; an attempt to fix the types of the genera and the exact basesand localities of the species. Proceedings of the Zoological Society of London 1911:120-158. [ Links ]
Velazco P.M. 2005. Morphological phylogeny of the bat genus Platyrrhinus Saussure, 1860 (Chiroptera: Phyllostomidae) with the description of four new species. Fieldiana, Zoology105:1-54. [ Links ]
Velazco P.M. 2013. On the phylogenetic position of Carollia manu Pa-checo et al., 2004 (Chiroptera: Phyllostomidae: Carolliinae).Zootaxa 3718:267-276. doi: http://dx.doi.org/10.11646/zootaxa.3718.3.3 [ Links ]
Velazco P.M. & B.D. Patterson. 2008. Phylogenetics and biogeography of the broad-nosed bats, genus Platyrrhinus (Chiroptera: Phyllostomidae). Molecular Phylogenetics andEvolution 49:749-759. doi: http://dx.doi.org/10.1016/j.ympev.2008.09.015 [ Links ]
Velazco P.M., A.L. Gardner & B.D. Patterson. 2010. Systematics of thePlatyrrhinus helleri species complex (Chiroptera: Phyllostomidae), with descriptions of two new species. Zoological Journal of the Linnean Society 159:785-812. doi: http://dx.doi.org/10.1111/j.1096-3642.2009.00610.x [ Links ]
Wetterer A.L., M.V. Rockman & N.B. Simmons. 2000. Phylogeny of Phyllostomid bats (Mammalia: Chiroptera): data from diverse morphological systems, sex chromosomes, and restriction sites. Bulletin of the American Museum of Natural History 248:1-200. [ Links ]
Wright A.J., R.A. Van Den Bussche, B.K. Lim, M.D. Engstrom& R.J. Baker. 1999. Systematics of the genera Carolliaand Rhinophylla based on the cytochrome-b gene. Journal of mammalogy 80:1202-1213. doi: http://dx.doi.org/10.2307/1383171 [ Links ]
York H.A. & M. Papes 2007. Limiting similarity and species assemblages in the short-tailed fruit bats. Journal of zoology273:249-256. doi: http://dx.doi.org/10.1111/j.14697998.2007.00321.x [ Links ]
Zurc D. & P.M. Velazco. 2010. Análisis morfológico y morfométrico de Carollia colombiana Cuartas et al. 2001 y C. monohernandezi Muñoz et al. 2004 (Phyllostomidae: Carollinae) en Colombia. Chiroptera neotropical 16(1):567-572. [ Links ]
Apéndice 1. Especímenes examinados y mapeados en este estudio.
Carollia benkeithi (n= 20): PERÚ: Junín: Satipo, Capitiri, 2 (MUSM 35497, 35498), Satipo, Coriteni, 1 (MUSM 35499). Loreto: Requena, Jenaro Herrera, 3 (MUSM 852, 853, 5552), San Martín: Bellavista, C.C. El Quinillal, 2 (MUSM 43792, 43794), Bellavista, Concesión para conservación "Valle del Biavo", 7 (MUSM 43797-43803), Picota, Concesión para conservación "Ojos de agua" (ABOFOA), 2 (MUSM 43795, 43796). Ucayali: Padre Abad, Padre Abad, B.N. Von Humboldt, 2 (MUSM 8608, 8602), Purús, Río Curanja, Balta, 1 (MUSM 1435)
Carollia brevicauda (n= 185): ECUADOR: Orellana: La Joya de los Sachas, 5 (QCAZ 2506, 3294, 3312, 3315, 3506), Orellana, Parque Nacional Yasuní, 1 (QCAZ 3507), Orellana, 1 (QCAZ 3511), Orellana, Sacha Lodge, 1 (QCAZ 3898), Pichincha: La Unión del Toachi, 1 (QCAZ 5412), Sucumbíos: Bosque Los Cedros, 1 (QCAZ 110), Pañacocha, 4 (QCAZ 3900-3902, 3905), Tungurahua: Río Negro, 4 (QCAZ 4546, 4548, 4592, 4684), Tungurahua, Río Machay, 2 (QCAZ 4543, 4545), Zamora Chinchipe: Parque Nacional Podocarpus, 1m (QCAZ 15378). PERÚ: Amazonas: Bongará, Yambrasbamba, ACP Abra patricia-Alto Nieva, 1 (MUSM 25163), San Carlos, Río Utcubamba, entre Churuja y Pedro Ruiz, 1 (MUSM 4852), Chachapoyas, Balsas, Balsas, 3 km E, 1 (MUSM 4850), Condorcanqui, El Cenepa, Condorcanqui, Puesto de Vigilancia 22 (Falso Paquisha), Cordillera el Cóndor, 10 (MUSM 309-311, 319, 331, 342, 352, 353, 364, 366), Condorcanqui, El Cenepa, Cordillera del Condor, Río Comaina, Puesto de Vigilancia 3 Alfonso Ugarte, 1 (MUSM 10593), Luya, San Jerónimo, Río Utcubamba, 11 km N by road Pedro Ruiz, 1 (MUSM 4853), Utcubamba, Cajaruro, ca 20 trail km E- de La Peca, 2 (MUSM 121, 193). Ayacucho: La Mar, Santa Rosa, Huanhuachayo, 1 (MUSM 1408). Cajamarca: Chota, Querocoto, La Granja, Quebrada Checos, 3 (MUSM 39381, 40304, 41527), San Ignacio, Tabaconas, Anexo Cerro Grande, 2 (MUSM 24944, 24945), San Ignacio, Tabaconas, Anexo La Lama, 12 (MUSM 24946-24954, 37900-37902), San Ignacio, Tabaconas, Cachango, 9 (MUSM 25014-25019, 37903-37905), San Ignacio, Tabaconas, Manchara, 5 (MUSM 25120-25124), San Ignacio, Tabaconas, Palmapampa, 14 (MUSM 25020, 25021, 25125-25135, 37906), San Ignacio, Tabaconas, Pueblo de Tabaconas, 3 (MUSM 25136-25138), San Ignacio, San Ignacio, San Ignacio, Chirinos, Nuevo Chalaquito "El Chaupe", 3 (MUSM 12633-12635), San Ignacio, Namballe, San Ignacio, El Sauce, 7 (MUSM 18164, 18211-18216), San Ignacio, Tabaconas, Tierra Blanca, 7 (MUSM 25022-25026, 25139, 25140). Cusco: La Convención, Echarate, La Convención, Camisea, Armihuari, 1 (MUSM 13540), La Convención, Quimbiri, La Convención, Kimbiri, 1 (MUSM 14529), Paucartambo, Challabamba, Bosque de las Nubes, 1 (MUSM 8788), Paucartambo, Kosñipata, Consuelo, 2 (MUSM 19638, 19639), Paucartambo, Kosñipata, San Pedro, 2 (MUSM 8793, 19425). Huánuco: Huánuco, Chinchao, Caserío de San Pedro de Carpish, 2 (MUSM 18900, 18901), Huánuco, Chinchao, trail to Hacienda Patty below Carpish Pass, 1 (MUSM 1410). Junín: Chanchamayo, San Ramón, Bosque Puyu Sacha, 1 (MUSM 39013), Chanchamayo, San Ramón, Nueva Italia, 2 (MUSM 40379, 40380), Chanchamayo, Santuario Nacional Pampa Hermosa, Los Cedros, 2 (MUSM 41159, 41161), Chanchamayo, Santuario Nacional Pampa Hermosa, Podocarpus, 1 (MUSM 41162-41164, 41166-41168) Satipo, Pampa Hermosa, San Antonio, 1 (MUSM 40546). Loreto: Alto Amazonas, Andoas, Lote 1AB, Capahuari Norte, 1 (MUSM 41642), Datem del Marañón, Andoas, Soplin, 2 (MUSM 25849, 25850), Loreto, Trompeteros, Lote 1AB, 1 (MUSM 41606), Maynas, Iquitos, 25 km S de Iquitos, Estación Biológica Allpahuayo, 1 (MUSM 16467), Maynas, Alto Nanay, a 17.35 Km norte de la localidad de Albarenga, 1 (MUSM 26522), Maynas, Alto Nanay, A 20 Km sur de la localidad de Diamante Azul, 1 (MUSM 26568), Maynas, Iquitos, A 20.4 Km oeste de la localidad de Nueva Villa Belen, 1 (MUSM 26523), Maynas, San Juan Bautista, Bélgica km 62 carretera Iquitos-Nauta E del camino, 1 (MUSM 30996), Maynas, Fernando Lores, Estación Biológica Quebrada Blanco, 1 (MUSM 21095), Maynas, San Juan Bautista, Mishana Km 25 carretera Iquitos-Nauta, Reserva Allpahuayo-Mishana, 1 (MUSM 31116), Maynas, Torres Causana, Río Lagartococha, Campamento Catalino, 3 (MUSM 21107-21109), Requena, Jenaro Herrera, Centro de Investigación Jenaro Herrera, 4 (MUSM 849-851, 5543). Madre de Dios: Santuario Nacional Pampas del Heath, 1 (MUSM 11659), Manu, CICRA, 1 (MUSM 26080, 26081), Manu, Maskoitania, 13.4 km NNW Atalaya, 1 (MUSM 19635), Manu, Parque Nacional Manu, Estación Biológica Pakitza, 2 (MUSM 12279, 12292), Tambopata, Las Piedras, Reserva de Cusco Amazónico, 2 (MUSM 6132, 6133). Pasco: Oxapampa, Palcazú, Parque Nacional Yanachaga-Chemillen, 6 (MUSM 10194, 10195, 10200, 10206, 10210, 10213), Palcazú, Huancabamba, Comunidad Nativa Castillo, 1 (MUSM 392), Palcazú, Iscozacin, 2 (MUSM 718, 719), Sandia, San Juan del Oro, Challohuma, 1 (MUSM 26762), San Juan del Oro, Yanacocha, 2 (MUSM 26766, 26767), Alto Inambari, Yanacocha, 2 (MUSM 34963, 34967), San Juan del Oro, Yanahuaya, 1 (MUSM 26772). San Martín: Mariscal Cáceres, Huicungo, Añazco, 1 (MUSM 24364), Mariscal Cáceres, Huicungo, La Meseta, 1 (MUSM 24366), Parque Nacional Río Abiseo, Las Palmas, 1 (MUSM 7246, 7247), Picota, Pucacaca,
C.C. Ojos de agua (ABOFOA), 1 (MUSM 43805). Tumbes: Tumbes, Pampas de Hospital, Angostura, Platanal, 1 (MUSM 22101), Pampas de Hospital, Quebrada Faical, 2 (MUSM 19169, 19280), San Jacinto, Rica Playa, Rio Tumbes, 1 (MUSM 22108). Ucayali: Coronel Portillo, Callaria, Lado SE Cerro Tahuayo, 1 (MUSM 1523), Padre Abad, Irazola, Bosque Nacional Von Humboldt, 1 (MUSM 8601), Purús, Purús, Concesión de Conservación Río La Novia, 2 (MUSM 44169, 44467), Purús, Río Curanja, Balta, 1 (MUSM 1416).
Carollia manu (n= 14): PERÚ: Cusco: La Convención, La Convención, Kimbiri, Camp. Llactahuaman, 1 (MUSM 14535), Paucartambo, Bosque de las Nubes, carretera Paucartambo-Pillcopata, km. 150, puente Union, 2 (MUSM 8799, 8801), Paucartambo, Estación Biológica Bosque Nublado, San Pedro, 2 (MUSM 11784, 11786). Puno: Sandia, 3 km N Yanahuaya, 4 (MUSM 34968-34970, 34978), Sandia, Curva Alegre, 1 (MUSM 26618), Sandia, San Fermín, 1 (MUSM 26612), Sandia, Yanacocha, 3 (MUSM 34971, 34975, 34977)
Carollia perspicillata (n= 190): ECUADOR: Esmeraldas, Esmeraldas, 1 (MECN 2035), Esmeraldas, Muisne, Río San Francisco, Estero La Ines, 1 (MECN 3024). PERÚ: Amazonas: Bagua, ca 20 road km SW Chiriaco, 2 (MUSM 1455, 1456), Amazonas, Bongará, Río Utcubamba, entre Churuja y Pedro Ruiz, 1 (MUSM 4858), Balsas, 19 km by road E (MUSM 4857), Balsas, 3 km E (MUSM 4849, 4851, 4856), Condorcanqui, Puesto de Vigilancia 22 (Falso Paquisha) Cordillera el Cóndor, 8 (MUSM 312-314, 316-318, 332, 339), Luya, Mendán, 1 (MUSM 38582), Luya, Río Utcubamba, 11 km N by road Pedro Ruiz, 2 (MUSM 4855, 4859), Luya, San Lucas, 2 (MUSM 38584, 38585), Luya, Tuén, 1 (MUSM 38587), Utcubamba, Buenos Aires (Bagua Grande), 1 (MUSM 1453), Utcubamba, Chiñuña, 1 (MUSM 38580). Ayacucho: La Mar, Ccentabamba, 2 (MUSM 21369, 21371). Cajamarca: Chota, La Granja, Quebrada Checos, 2 (MUSM 39382, 40303), San Ignacio, Anexo La Lama, Quebrada el Salado, 2 (MUSM 24955, 24956), San Ignacio, Palmapampa, 5 (MUSM 25027, 25141-25143, 37833). Cusco: La Convención, Kp 127, 1 (MUSM 40485), La Convención, KP 22, 1 (MUSM 38782), La Convención, Camisea, Armihuari, 1 (MUSM 13591), La Convención, Camisea, Pagoreni, 1 (MUSM 13600), La Convención, Camisea, San Martín, 1 (MUSM 13602), La Convención, Camisea, Segakiato, 1 (MUSM 14733), La Convención, Kimbiri, Campamento Llactahuaman, 2 (MUSM 14533, 14536), La Convención, Vilcabamba 2, 1 (MUSM 38783), Paucartambo, Sabaluyo (Pillcopata), 2 (MUSM 8559, 8560). Huánuco: Leoncio Prado, ca 35 km NE de Tingo Maria, Hacienda Santa Elena, 2 (MUSM 155, 156), Puerto Inca, Estación Biológica Panguana, 2 (MUSM 1470, 36691). Junín: Chanchamayo, Centro poblado Nueva Italia, 5 (MUSM 40384, 40385, 41169, 41171, 41317), Satipo, Calabaza, 1 (MUSM 42844), Satipo, Capitiri, 1 (MUSM 35503), Satipo, Coriteni, 1 (MUSM 35509), Satipo, Poyeni, 3 (MUSM 35510, 35512, 35516), Satipo, San Antonio, 3 (MUSM 40551, 40554, 40555), Satipo, Santa Bibiana, 2 (MUSM 40562, 40563). Loreto: Rio Yarapa, Albergue Yacumama, SE Iquitos, 2 (MUSM 9418, 9419), Alto Amazonas, Lote 1AB, Capahuari Norte, 1 (MUSM 41643), Alto Amazonas, Trueno, 1 (MUSM 16357), Loreto, Lote 1AB, Jibarito, 1 (MUSM 41609), Loreto, Lote 1AB, Shiviyacu, 4 (MUSM 41613, 41644, 41646, 41649), Loreto, Río Samiria (Flor de Yarina), Estación Biológica Pithecia, 1 (MUSM 1415), Maynas, 25 km S de Iquitos, Estación Biológica Allpahuayo, 6 (MUSM 16466, 16468, 16469, 16471-16473), Maynas, 25 km S de Iquitos, Estación Biológica Allpahuayo, 1 (MUSM 16475), Maynas, Estación Biológica Quebrada Blanco, 1 (MUSM 21094), Maynas, Mishana, río Nanay, 2 (MUSM 31123, 31583), Maynas, Quebrada Agua Negra, 1 (MUSM 24418), Maynas, Río Lagartococha, Campamento Catalino, 3 (MUSM 21118, 21122, 21125), Requena, Centro de Investigación Jenaro Herrera, 2 (MUSM 5544, 5548), Requena, Jenaro Herrera, 6 (MUSM 854-857, 1054, 5920), Ucayali, Sierra de Contamana, Cerros de Canchaguaya, Aguas Calientes, 1 (MUSM 17958). Madre de Dios: Manu, CICRA, Cruce Sachavaca, Bajío, 1 (MUSM 26082), Manu, Estación Biológica Cocha Cashu, 1 (MUSM 5107), Manu, Pakitza, 3 (MUSM 189, 6852, 26122), Tambopata, Explorer's Inn Lodge, 2 (MUSM 1487, 1500), Tambopata, Reserva de Cusco Amazónico, 2 (MUSM 6163, 6164). Pasco: Oxapampa, Estación Biológica Paujil, 1 (MUSM 24192), Oxapampa, Hacienda Roca-Lux, Lontananza, Rio Mucñis, 1 (MUSM 10202), Oxapampa, Iscozacin, 1 (MUSM 717), Oxapampa, Río Pescado, 3 (MUSM 24193, 24195, 24196). Puno: Carabaya, Carabaya, Coasa, Boca de Quebrada Ursulinda con Rio Candamo, 1 (MUSM 15799), Sandia, Challohuma, 1 (MUSM 26773), Sandia, Curva Alegre, 1 (MUSM 26619), Sandia, San Fermin, 3 (MUSM 26620, 26622, 26623), Sandia, Yanahuaya, 1 (MUSM 26774). San Martín: Bellavista, Concesión para conservación El Incaico, 2 (MUSM 43807, 43809), Bellavista, C.C. El Quinillal, 8 (MUSM 43804, 43810-43813, 43817, 43820, 43822), Lamas, Lamas, 1 (MUSM 1503). Tumbes: Tumbes, Angostura, Platanal, 5 (MUSM 22102-22106), Tumbes, Quebrada Angostura, 8 (MUSM 19273-19276, 19278, 19287, 19288, 19290), Tumbes, Quebrada Faical, 2 (MUSM 22096, 24477), Tumbes, Quebrada Las Pavas, 1 (MUSM 24478), Tumbes, Rica Playa, Rio Tumbes,1 (MUSM 22109), Zarumilla, Campo Verde, 5 (MUSM 19270, 19272, 19284, 22110, 22111), Zarumilla, Naranjal, 4 (MUSM 22097, 22098, 22112, 22113), Zarumilla, Quebrada Campo Verde, 1 (MUSM 22114), Zarumilla, Quebrada Naranjos, 4 (MUSM 19173, 19174, 19282, 19292), Zarumilla, Rio Zarumilla, Cabaña INRENA, 1 (MUSM 22115), Zarumilla, Rio Zarumilla, Carrizalillo, 4 (MUSM 22099, 22117, 22118, 22120). Ucayali: Ucayali, afluente del Río Mamanchita, 1 (MUSM 630), Coronel Portillo, Pucallpa, 4 (MUSM 1511, 1516, 1471, 1497), Padre Abad, Padre Abad, Bosque Nacional Von Humboldt, 2 (MUSM 8611, 8613), Purús, Concesión de Conservación Río La Novia, 7 (MUSM 44170, 44171, 44175-44178, 44182)
Carollia sp. (n= 20): Amazonas, Condorcanqui, Condorcanqui, Puesto de Vigilancia 22 (Falso Paquisha) Cordillera el Cóndor, 2 (MUSM 320, 359), Río Yarapa, Albergue Yacumama, SE Iquitos, 1 (MUSM 9420), Loreto, Lote 1AB, 3 (41607, 41608, 41640), Maynas, Bélgica, 2 (MUSM 31168, 31174), Maynas, Camino a El Paujil, 1,8 km al W del km 35 de la carretera Iquitos-Nauta, 1 (MUSM 27592), Maynas, Reserva Allpahuayo-Mishana, 1 (MUSM 31248), Maynas, Paujil, W km 37.45 de la carretera Iquitos-Nauta, 1 (MUSM 27612), Maynas, Peña Negra, 1 (MUSM 27614), Maynas, Quebrada Oran, ca 5 km N Río Amazonas, 3 (MUSM 1437, 1438, 1443), Maynas, San Lucas, 1 (MUSM 27628), Maynas, Varillal, 1 (MUSM 27630).
Fuentes de financiamiento: Proyecto INNÓVATE-PERÚ mediante el contrato N° 402-PNICP-PIBA-2014
Información sobre los autores: La autora no incurre en conflictos de intereses.
Presentado: 13/08/2017
Aceptado: 04/11/2017
Publicado online: 20/12/2017

