











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Neoraimondia es un género de cactus arbóreo de América del Sur, perteneciente a la tribu Browningieae Buxbaum (Ostolaza 2014, Cáceres et al. 2005). Son conocidas solo dos especies, una es N. herzogiana endémica de Bolivia y N. arequipensis endémica de Perú, la cual tiene amplia distribución en la costa y sierra peruana (Ostolaza 2011, Cáceres et al. 2005). Asimismo, N. arequipensis presenta tres subespecies N. arequipensis subsp. arequipensis del sur del Perú, N. arequipensis subsp. roseiflora (Werd. & Backeberg) Ostolaza, distribuida en los valles y quebradas del departamento de Lima, Ancash y valles de Chincha y Pisco, y N. arequipensis subsp. gigantea, distribuida al norte del Perú, en los departamentos de La Libertad, Lambayeque, Piura y Cajamarca (Ostolaza 2011, Ostolaza 2014, Vila & Eastwood 2006).
El hábitat de esta cactácea columnar comprende la línea de costa rocosa, colinas costeras, matorral desértico y valles interandinos del Perú (Cáceres et al. 2005). En estos ambientes, cumplen un rol importante en la cadena trófica de diversos organismos, siendo sus flores y frutos utilizados como alimento de aves, roedores e insectos (Ceroni et al. 2007, Lerner et al. 2003, Novoa et al. 2005). Asimismo, los tallos son utilizados como soporte para nidos de algunas aves, refugio contra depredadores y también ofrecen condiciones adecuadas para el desarrollo de otros cactus y el establecimiento de plantas jóvenes (Novoa et al. 2005, Cáceres et al. 2005).
En el año 2014, en las provincias de Tumbes y Contralmirante Villar, del departamento de Tumbes, se observó la presencia de dos especímenes de cactus de la especie Neoraimondia arequipensis, cuya distribución geográfica registrada comprendía toda la costa del litoral peruano hasta solamente el departamento de Piura. Bajo este contexto, se procedió a realizar una búsqueda en toda la línea costera de Tumbes con la finalidad de registrar más individuos de esta especie. La presente investigación reporta los nuevos límites de distribución norte de esta especie de cactus arbustivo en el departamento de Tumbes.
Material y métodos
El estudio se realizó en las provincias de Contralmirante Villar (distritos de Canoas de Punta Sal, Zorritos y Casitas) y Tumbes (distritos de Tumbes, Corrales y La Cruz) del departamento de Tumbes, Perú. Esta ciudad limita por el norte y este con Ecuador, por el sur con Piura y el oeste con el Océano Pacífico. Según el Servicio Nacional de Meteorología e Hidrología (SENAMHI), Tumbes presenta dos climas marcados, uno de tipo templado sub-húmedo, situados entre 1000 a 3000 m de altitud, y precipitación anual que puede llegar hasta 1200 mm en las partes más elevadas, húmedas y orientales (SENAMHI 2019). Asimismo, presenta clima semi-cálido (desérticoárido- subtropical) y con lluvias escasas en la mayor parte del año que se acentúa hacia el territorio piurano. De manera general hay presencia de sol casi todo el año, con una temperatura máxima de 30 °C, mínima de 19 °C y con un promedio anual de 24 °C (MINAM 2009).
Se evaluó la presencia de ejemplares de cactus "gigantón" en diferentes puntos aledaños de la zona costera, desde el distrito de Canoas de Punta Sal (4°1'32.11"S; 80°57'50.13"W) hasta el distrito de Tumbes (3°35'7.36"S; 80°24'55.47"W). Cada punto de muestreo fue georeferenciado con un GPS Garmin modelo eTrex, se realizó el registro fotográfico de cada individuo identificado in situ (según Ostolaza 2014) y además se registró la presencia de fauna y flora acompañante. Para la identificación fenotípica se colectó partes de un espécimen de cactus Neoraimondia ubicado en Zorritos (03°40'48.43"S; 80°40'14,52"W), fue codificada como B (Fig. 1) y depositada en la colección de especímenes del Herbarium Truxillense (HUT), con el código de ingreso N° 59825.
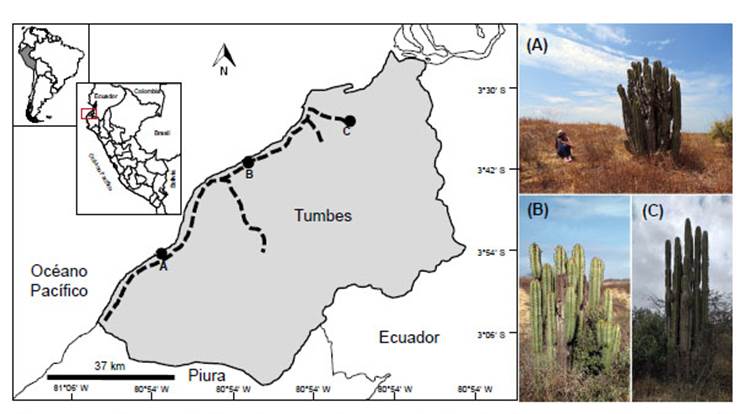
Figura 1 Mapa de recorrido (líneas discontinuas) y puntos de colecta de muestra para análisis molecular de Neoraimondia arequipensis. A, distrito de Canoas de Punta Sal; B, distrito de Zorritos y C, distrito de Tumbes.
Por otro lado, tres muestras de tejido floral codificadas como A, B y C (Fig. 1), fueron colectadas para identificación molecular mediante secuenciación del gen matK (Maturase K gene). Para esto, la extracción de ADN se realizó mediante el método CTAB-DTAB (Gustincich et al. 1991), luego la reacción en cadena de la polimerasa (PCR por sus siglas en inglés) se preparó en 20 µL de volumen final de reacción con PCR 2X Taq master mix (GeneOn GmbH), 0.5 µM de los cebadores matK-xf (TAATTTACGATCAATTCATTC) y matK-MALP (ACAAGAAAGTCGAAGTAT) (Ford et al. 2009; Dunning & Savolainen 2010) y 1.0 µL de ADN (10 µg/mL). El proceso de amplificación se realizó en un termociclador (Blue-Ray Biotech), iniciando con 98 °C durante 5 minutos, seguida de 35 ciclos de desnaturalización (98 °C por 10 s), hibridación (54 °C por 30 s) y elongación (72 °C por 40 s), seguido por una extensión final de 10 min a 72 °C. Los productos de amplificación de la PCR fueron analizados en gel de agarosa al 1.8% con buffer de migración TAE 1X a 120V por 30 min, coloreados posteriormente en solución de bromuro de etidio (1 mg/mL) y visualizados empleando un transiluminador UV Thermo Scientific. Para la secuenciación de ambas cadenas de los productos de amplificación, 10 µL de cada una fueron empacadas y enviadas a la empresa Macrogen de Korea, incluyendo 5 µL de cada cebador para el gen matK. Las secuencias forward y reverse obtenidas de cada producto de la PCR fueron procesadas mediante el programa Mega 7 (Kumar et al. 2016), con la finalidad de visualizar los cromatogramas y generar las secuencias consenso. Una vez obtenidas dichas secuencias se utilizó la herramienta Nucleotide BLAST (Basic Local Alignment Search Tool) para verificar la identidad de espécimen colectado. Las secuencias fueron depositadas en el GenBank con los números de accesión:MW160169, MW160170 y MW160171.
Resultados
Se evaluaron diferentes zonas naturales de los distritos de Tumbes, Corrales, La Cruz, Zorritos, Casitas y Canoas de Punta Sal; identificándose un total de 70 individuos de Neoraimondia arequipensis. El ambiente donde fue encontrada con más frecuencia esta especie se caracteriza por tener suelo de tipo pedregoso y rocoso, aledaños a zonas de quebrada y en altitudes entre 20 a 60 m de altitud (Tabla 1; Fig. 2E).
Tabla 1 Número de especímenes de Neoraimondia arequipensis identificados fenotípicamente en el departamento de Tumbes, durante los años 2014 y 2019.

Figura 2 Neoraimondia arequipensis subsp. gigantea. (A) y (B) Flores rojas; (C) fruto en proceso de maduración; (D) vista superior del extremo de una rama mostrando 6 costillas; (E) hábitat rocoso; (F) ardilla nuca blanca (Sciurus stramineus) alimentándose de areolas; (G) Polistes sp. alimentándose de fruto; (H) presencia de hormiga Crematogaster sp. (flecha roja); (I) Fregata magnificens reposando en cactus "gigantón"; (J) utilización de cactus "gigantón" como protección de nidos de aves (flecha roja).
Las características fenotípicas encontradas incluyen cactus arbóreos de 40 cm hasta 5 m de altura máxima, tallos verdes y lignificados en los cactus más adultos, ramas erectas con 5 a 7 costillas, areolas apretadas ymuchas de ellas elongadas. Algunos cactus en proceso de floración presentaron flor y fruto rojo (Fig. 2A, B, C, D).
Los análisis moleculares de las muestras de cactus basados en el gen matK, en todos los casos muestran similitud superior a 99.87% para N. arequipensis (Tabla 2), comparada con la secuencia genética con número de accesión AY015299.1 de la base de datos del GenBank; que corresponde al estudio de relaciones filogenéticas en la familia Cactaceae basados en secuencias trnK/matK y trnL-trnF (Nyffeler 2002).
Tabla 2 Identificación molecular mediante secuenciación del gen matK (Maturase K gene) en muestras de tejido floral colectadas en el departamento de Tumbes.

Se observaron algunas interrelaciones con otros organismos, siendo N. arequipensis fuente de alimento para roedores (Sciurus stramineus) e insectos (Polistes sp. y Crematogaster sp.), protección para nidos de aves y soporte para el reposo de Fregata magnificens (Fig. 2F, G, H,I, J). En cuanto a la flora acompañante, se identificaronlas siguientes géneros Chloris, Pennisetum, Cenchrus, Bouteloua, Alternanthera; y las especies Cordia lutea, Bursera graveolens, Capparis angulata, Caesalpinia paipai, Ipomoea pes-caprae, Luffa operculata, Parkinsonia aculeata, Prosopis pallida, Armatocereus cartwrightianus, Melocactus peruvianus, Monvillea diffusa y Haageocereus versicolor.
Discusión
Neoraimondia arequipensis ha sido reportada por el norte hasta el departamento de Piura (Ostolaza 2019a, 2014b, 2011c, Ostalaza & Cáceres 2017), con registros en las áreas naturales del Coto de Caza El Angolo (Sullana), tablazos de Máncora (Talara) (Sabogal & Zerbe 2005; IGP 2017; La Torre-Cuadros & Linares-Palomino 2008), bosque de Jambur del distrito de Suyo y bosques de La Menta y Timbes, ambos en la provincia de Ayabaca, en la zona este de Piura (Fiestas 2016, Rasal et al. 2011). Mientras que, para Tumbes solamente se cuenta con una mención sin mayores detalles (Aguirre et al. 2006), en un estudio de especies leñosas en los bosques estacionalmente secos de Ecuador y Perú, y un reporte fotográfico en la provincia de Contralmirante Villar, con ubicación georeferenciada, pero sin caracterizar el ejemplar evaluado (Flanagan 2018).
Por otro lado, Tumbes cuenta con zonas naturales ycondiciones ambientales propicias para el establecimiento de N. arequipensis, sobre todo en la parte sur correspondiente a la línea costera del distrito de Canoas de Punta Sal, que se encuentra dentro de la zona de vida matorraldesértico tropical md-T (MINAN 2009), donde se reportóla mayor cantidad de individuos de "gigantón". Esta zonase caracteriza por presentar suelos rocosos y pedregosos,con alto drenaje de agua, e intensas lluvias estacionales;así tenemos que estas condiciones son adecuadas para eldesarrollo de N. arequipensis (Cáceres et al. 2005). Por otraparte, al norte de Canoas de Punta Sal se presenta la zonade vida matorral desértico Premontano Tropical (md-PT), caracterizada por un relieve topográfico entre ondulado y quebrado con algunas áreas de pendientes suaves, conpredominio de suelos arcillosos (ONERN 1976).
Neoraimondia arequipensis subsp. gigantea cumpliría importantes funciones ecológicas en los ambientes áridosde Tumbes, como brindar estructuras (areolas, flores yfrutos) que son utilizadas como alimento por las ardillasde nuca blanca, avispas y hormigas (Figs. 2F, 2G y 2H), quea la vez favorecen la polinización y dispersión de semillas, así también, sirven como medio de protección de nidos enaves y para descanso de aves marinas (Figs. 2I y 2J).
Las características fenotípicas y genotípicas evaluadas en los individuos de cactus analizados, indican la presencia de N. arequipensis subsp. gigantea en dos provincias del departamento de Tumbes y es necesario realizar estudios complementarios que permitan conocer con mayor detalle la dinámica de esta población de cactácea con su entorno, proveyendo información básica para fomentar medidas de conservación a nivel regional.

