











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los peces son el grupo más diverso de vertebrados (Nelson et al. 2016) y en la Región Neotropical la diversidad de este grupo es muy alta (Albert et al. 2011). La Cuenca Amazónica es el sistema hidrográfico más largo del mundo con un total de 6869999 km2 (Goulding et al. 2003) y 2257 especies reconocidas (Oberdorff et al. 2019), sin embargo, esta cifra podría estar subestimada debido a que cada año son descritas nuevas especies (Antonelli et al. 2018, (Antonelli et al. 2018, Machado et al. 2018). En el Perú la primera lista anotada de peces fue publicada por Ortega y Vari (1986) registrando 735 especies de aguas continentales, mientras que, en la más reciente publicación, Ortega et al. (2012), reconocen 1064 especies nativas válidas de las cuales el 90% corresponde a la Cuenca Amazónica.
En general se señala que los estudios ictiológicos en la Amazonia son realizados tomando en cuenta ríos importantes y de fácil acceso (Junk & Soares 2001). En la región amazónica peruana los estudios se han desarrollado principalmente en grandes cuencas como la de Madre de Dios (Goulding et al. 2010, Carvalho et al. 2012), Urubamba (Carvalho et al. 2011, Ortega et al. 2010), Huallaga (Ortega et al. 2007) y en tributarios directos del sistema Ucayali-Amazonas (e.g. Carvalho et al. 2009, Quezada García et al. 2017, Correa & Ortega 2010, Rengifo 2007) quedando sistemas hidrográficos poco estudiados (Ortega et al. 2012). Uno de estos sistemas es la quebrada Mayapo, en la cuenca del río Tambo. En el presente trabajo se reporta la composición taxonómica ictiológica de la quebrada Mayapo en base al material depositado en la Colección Ictiológica del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MUSM).
Material y métodos
Área de estudio
La quebrada Mayapo pertenece a la intercuenca 49951 (MINAGRI-ANA 2018), conocida también como cuenca del río Tambo, está ubicada en el distrito de Río Tambo, provincia de Satipo, Región Junín, entre los 256 y 560 m de altitud. Biogeográficamente el área está incluida en la ecorregión Piedemonte UcayaliUrubamba en la vertiente oriental de la Cordillera de los Andes, la cuenca comprende la zona de transición entre el llano Amazónico y los Andes en la parte oeste de la Amazonia (Abell et al. 2008). El río Tambo se forma por la confluencia de los ríos Perené y Ene, es caudaloso y de velocidad moderada, recibe a la quebrada Mayapo por su margen derecha y se une con el río Urubamba para formar el río Ucayali (Hinostrosa 2010).
La quebrada Mayapo es un cuerpo agua de primer orden (Strahler 1957) y uno de los principales tributarios del río Tambo en el sector correspondiente al llano amazónico de la cuenca, en este sector los ríos se caracterizan por ser torrentosos con abundantes rápidos y fondo de cauce rocoso-pedregoso; la temperatura promedio anual es de 28°C a más con precipitaciones anuales de 2000 a 3000 mm y se caracteriza por presentar un clima muy húmedo y cálido sin déficit de agua durante el año (Hinostrosa 2010).
Revisión de material ictiológico
Se analizó exclusivamente material depositado en la Colección Ictiológica del MUSM proveniente de colectas realizadas en la quebrada Mayapo y afluentes durante los años 2005, 2009 y 2010 en 30 puntos de muestreo, uno de los cuales estuvo ubicado cerca a la desembocadura al río Tambo (Figura 1).
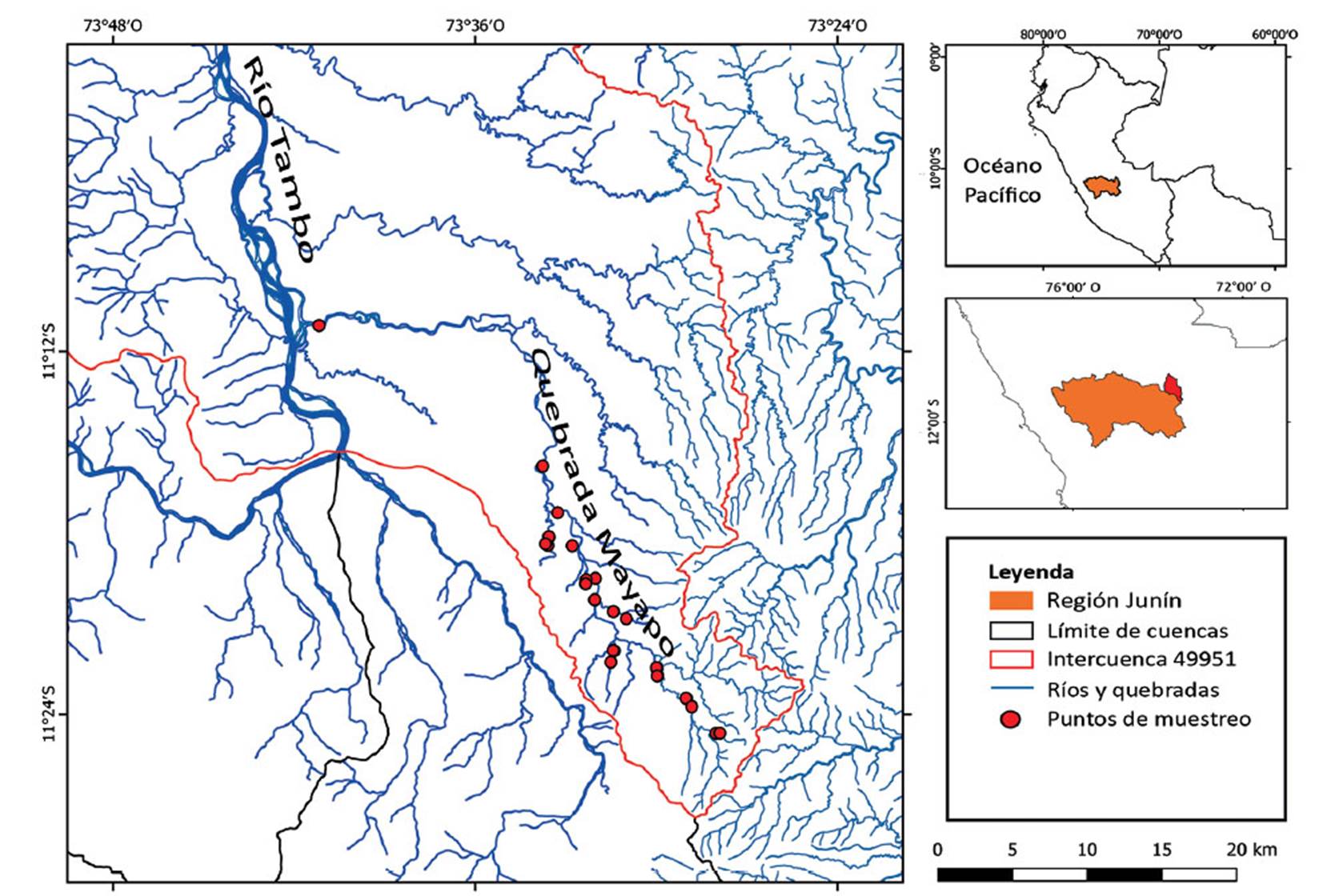
Figura 1 Mapa de ubicación de los puntos de muestreo y procedencia del material ictiológico de la quebrada Mayapo y afluentes.
El material fue recolectado con redes de arrastre a la orilla de 10 m de longitud x 3 m de altura y abertura de malla de 5 mm, formando un semicírculo a favor de la corriente; adicionalmente fueron empleadas redes de lance (atarrayas). Las muestras fueron fijadas en formol al 10% por un periodo de 24 a 48 horas para posteriormente ser preservadas en etanol al 70% y colocadas en bolsas plásticas envueltas en gasa humedecida para su traslado e identificación en el Departamento de Ictiología del MUSM.
El material, cada ejemplar, fue revisado haciendo uso de un microscopio estereoscópico en base caracteres externos morfológicos, morfométricos (con ayuda de un calibrador digital de 150 mm con 0.05 mm de precisión) y merísticos, procurando llegar hasta el nivel de especie. Se emplearon claves taxonómicas, descripciones originales de especies y literatura ictiológica especializada. Para el orden Characiformes se usaron como referencia los trabajos de Bertaco y Malabarba (2010), Bührnheim y Malabarba (2006), Chernoff y Machado-Allison (1990), Ferreira (2007), Gery (1977), de Lucena y Malabarba (2010), Malabarba (2004), Pavanelli (1999), VanegasRíos y Urbano-Bonilla (2017), Vari y Ortega (2000), Vari (1991), Vari y Harold (2001) y Weitzman y Ortega (1995); para Siluriformes a Burgess (1989), Covain y Fisch-Muller (2007), Eigenmann (1917), Nijssen y Isbrücker (1986), Schultz (1944), Vari et al. (2005); para Beloniformes a Collete (1974); y para Cichliformes a Kullander (1986).
El material se encuentra debidamente catalogado y depositado en la Colección Ictiológica del MUSM. La lista de especies fue organizada por órdenes y familias de acuerdo con lo propuesto por Eschmeyer y Fong (http:// researcharchive.calacademy.org/research/ichthyology/ catalog/SpeciesByFamily.asp), dentro de cada familia los géneros y especies fueron ordenados alfabéticamente.
Resultados y discusión
Fueron analizados un total de 8316 individuos provenientes de la quebrada Mayapo y afluentes de las colectas de los años 2005, 2009 y 2010. Se identificaron 60 especies agrupadas en 39 géneros, 16 familias y cuatro órdenes (Tabla 1). El orden Characiformes presentó la mayor riqueza específica con 42 especies (70.0% del total), seguido por Siluriformes con 13 especies (21.7%), teniendo los órdenes Cichliformes y Beloniformes en conjunto cinco especies (8.3%). A nivel de familias, las mejores representadas fueron Characidae con 32 especies (54.1%) seguida de Loricariidae con ocho (26.1%). Los géneros con mayor predominancia de especies fueron Astyanax y Knodus, ambos pertenecientes a la familia Characidae, con cinco y cuatro especies, respectivamente. No se registraron especies introducidas o invasoras.
En algunos casos no fue posible identificar hasta el nivel de especie a algunos ejemplares debido a su condición de juveniles, que no permite el desarrollo de las características distintivas de cada especie, o a que no coincidieron con las descripciones y/o claves taxonómicas vigentes.
Se observa la predominancia de especies de los órdenes Characiformes (peces con escamas y sin espinas en las aletas) y Siluriformes (peces con placas o piel desnuda), lo que concuerda con el patrón descrito para la Región Neotropical (Albert et al. 2011, Reis et al. 2003). Este patrón también ha sido observado en otras regiones de la Amazonia Peruana como en la cuenca de los ríos Perené (Salcedo 1998), Bajo Pachitea (Palacios et al. 2008), Urubamba (Carvalho et al. 2011, Ortega et al. 2010), Madre de Dios (Goulding et al. 2010, Carvalho et al. 2012), Aguaytía (Quezada García et al., 2017), Yuruá (Carvalho et al. 2009, Rengifo 2007) y la parte baja del río Nanay (Correa & Ortega 2010) (Figura 2). La familia Characidae es la más diversa y compleja de peces neotropicales (Mirande 2010, de Lima et al. 2013, Britski et al. 1999, Nelson et al. 2016) y en nuestro estudio también fue la más diversa.
El género Astyanax tiene una amplia distribución que incluye la región tropical y subtropical de América desde Estados Unidos hasta Argentina (Gery 1977). Las especies Astyanax abramis, Astyanax bimaculatus y Astyanax maximus, presentes en la quebrada Mayapo, han sido reportadas en otras evaluaciones ictiológicas como en el Bajo Urubamba (Carvalho et al. 2011), Las Piedras (Carvalho et al. 2012) y Aguaytía (Quezada et al. 2017). El género Knodus presenta una amplia distribución en la Amazonia, desde el noreste de Sudamérica en el piedemonte amazónico hasta el sudoeste de Brasil (Ferreira 2007, Gery 1977). Las especies Knodus hypopterus y Knodus savannensis, presentes en nuestros resultados, también han sido reportadas en áreas relativamente cercanas como en los ríos Aguaytía (Quezada et al. 2017) y Bajo Urubamba (Carvalho et al. 2011). La especie Ancistrus tamboensis presenta una distribución más restringida, fue descrita para la cuenca del río Tambo (Fowler 1945), habiéndose registrado hasta el momento en el presente estudio y en la cuenca del río Perené (Salcedo 1998) que se encuentra muy próxima al área de estudio.
Las especies Hoplias malabaricus "fasaco", Prochilodus nigricans "boquichico" y Triportheus angulatus "sardina" son componentes importantes de los desembarques pesqueros con fines de consumo en toda la región amazónica (Barletta et al. 2016 (Barletta et al. 2016, García-Dávila et al. 2018, Garcia et al. 2009, Doria et al. 2018, Barthem & Goulding 2007, Diaz-Sarmiento & Alvarez-León 2003) y presentan una amplia distribución en toda la Amazonia. Otros géneros de importancia de consumo, aunque en menor grado, son Serrasalmus (Doria et al. 2018, GarcíaDávila et al. 2018, Diaz-Sarmiento & Alvarez-León 2003) y Crenicichla (García-Dávila et al. 2018). Además, en varias comunidades amazónicas son consumidas diversas especies robustas que lleguen a un tamaño regular (aproximadamente 10 cm de longitud) como algunos individuos de los géneros Astyanax, Parodon, Steindachnerina, Pimelodus y Bujurquina. Las especies de los géneros Prochilodus, Serrasalmus y Triportheus son consideradas como migradoras, siendo las migraciones de los géneros Serrasalmus y Triportheus estacionales y menores a 100 km (Diaz-Sarmiento & Alvarez-León 2003), mientras que en P. nigricans las migraciones son más largas, pudiendo consistir en varios cientos de kilómetros, durante la época de aguas altas con fines reproductivos (García-Dávila et al. 2018, Diaz-Sarmiento & Alvarez-León 2003).
La riqueza de especies encontrada en la quebrada Mayapo (60) se encuentra por debajo de otras evaluaciones como las realizadas en los ríos Bajo Urubamba (Carvalho et al. 2011), Madre de Dios (Carvalho et al. 2012), Aguaytía (Quezada et al. 2017), Yuruá (Carvalho et al. 2009) y Nanay (Correa & Ortega 2010), donde el número de especies varió entre 86 y 211. Un factor que influye en la variación de la riqueza de especies es el esfuerzo de pesca (Oberdorff et al. 2019), lo que explicaría el bajo número de especies en el río Nanay con cuatro puntos de muestreo (durante dos temporadas) frente un rango de 11 a 30 puntos de muestreo en las otras evaluaciones mencionadas (Figura 3).

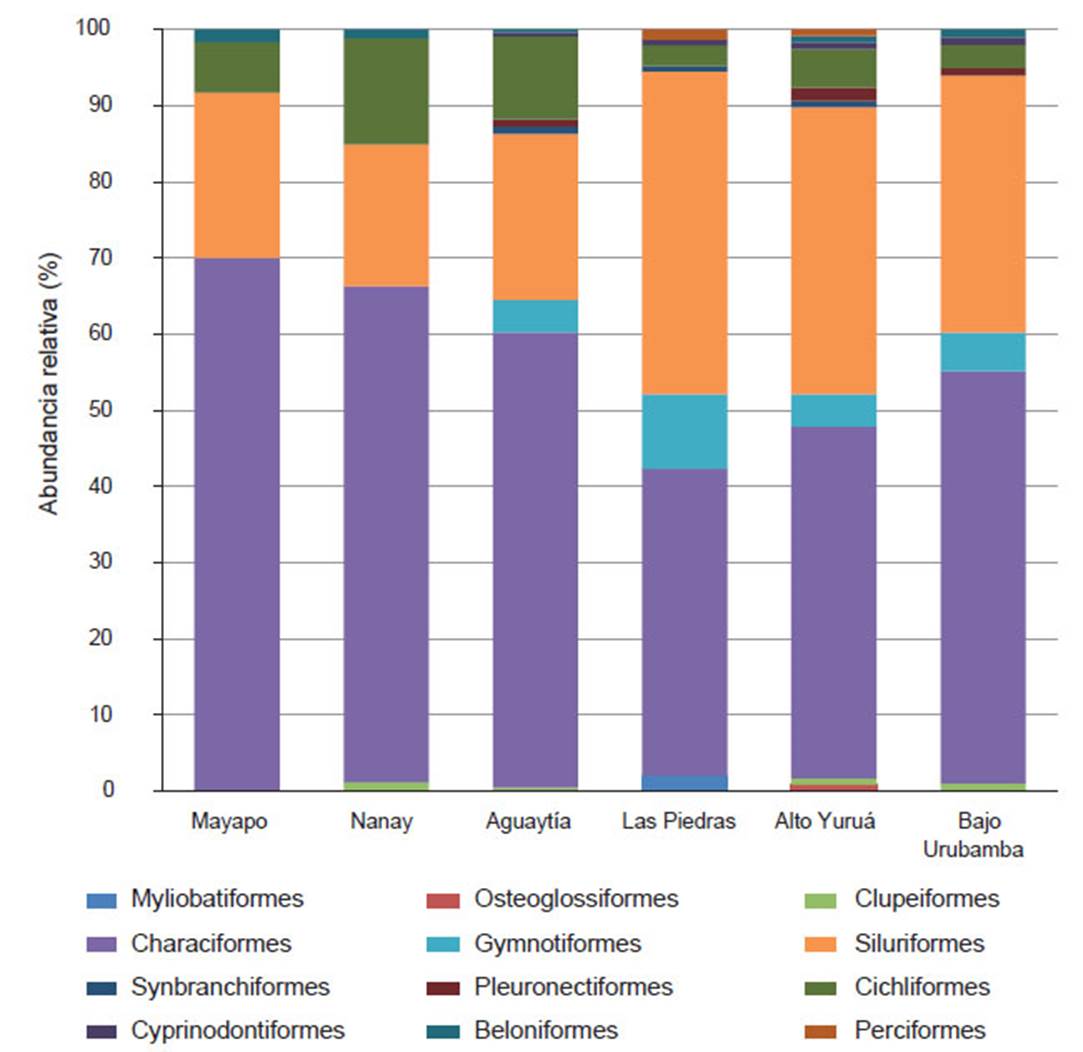
Figura 2 Abundancia relativa de especies por órdenes presentes en distintas evaluaciones ictiológicas realizadas en la Amazonia Peruana: Mayapo (presente evaluación), Nanay (Correa & Ortega 2010), Aguaytía (Quezada et al. 2017), Las Piedras (Carvalho et al. 2012), Alto Yuruá (Carvalho et al., 2009) y Bajo Urubamba (Carvalho et al. 2011).
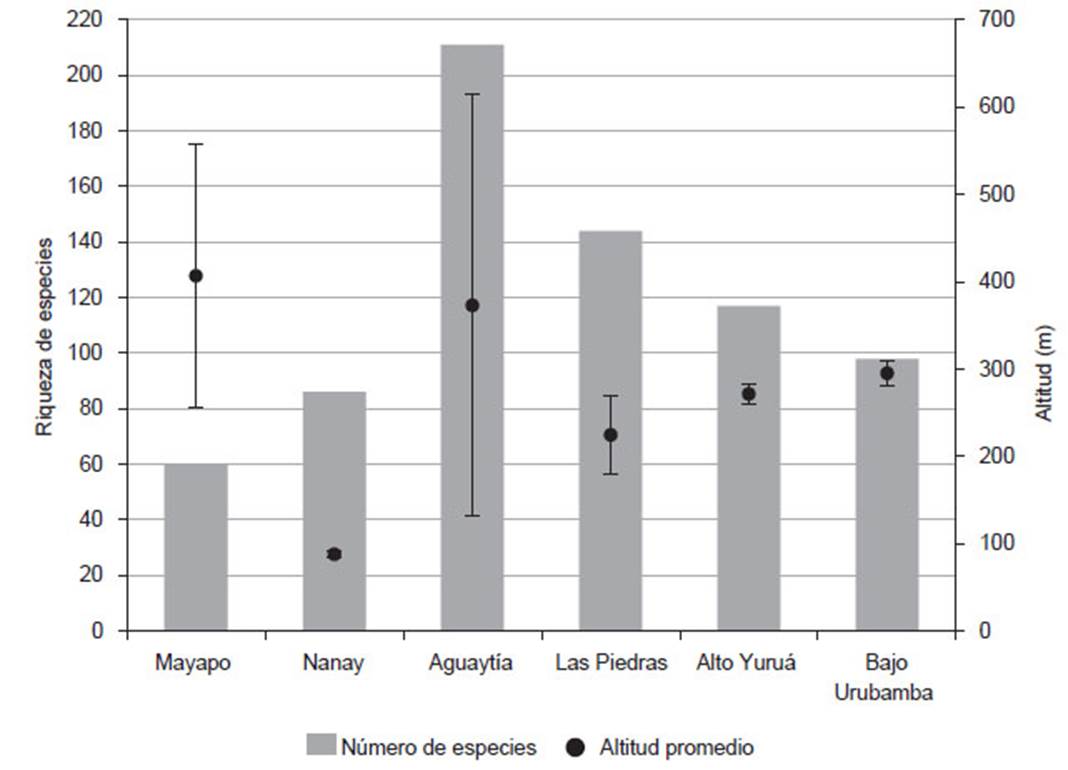
Figura 3 Riqueza de especies reportadas y rango altitudinal (ɪ) de distintas evaluaciones ictiológicas realizadas en la Amazonia Peruana: Mayapo (presente evaluación), Nanay (Correa & Ortega 2010), Aguaytía (Quezada et al. 2017), Las Piedras (Carvalho et al. 2012), Alto Yuruá (Carvalho et al. 2009) y Bajo Urubamba (Carvalho et al. 2011).
Otro factor que estaría influenciando nuestros resultados sería la variación altitudinal donde, de acuerdo con el concepto del río continuo, la diversidad es menor en las cabeceras de los ríos y mayor en las desembocaduras (Tomanova et al. 2007, Vannote et al. 1980). Este patrón ha sido también observado en la región amazónica (Lujan et al., 2013). Las evaluaciones en la quebrada Mayapo (entre 256 y 557 m de altitud) y en la cuenca del río Perené (45 especies registradas a 600 m de altitud aproximadamente) presentaron una moderada cantidad de especies, lo que estaría relacionado a su ubicación altitudinal. Por otro lado, la alta riqueza de especies registradas en el río Aguaytía (211) estaría relacionada a la amplia variación altitudinal en la que fueron realizados los muestreos (entre 132 y 614 m de altitud). De manera similar, la menor variedad de órdenes se presentó a mayor altitud, habiéndose reportado cuatro órdenes en la quebrada Mayapo (presente evaluación) y en la cuenca del río Perené (Salcedo 1998)

