











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El estudio de la dieta de las aves rapaces a partir del análisis de sus egagrópilas puede ayudar a conocer mejor la distribución, abundancia, conducta y vulnerabilidad de las especies presa (Fulk 1976, Marti 1987). Además, en algunos casos ha permitido el descubrimiento de nuevas especies de roedores (e.g., Massoia 1979) y la extensión de la distribución conocida de otras (e.g., Pardiñas & Galliari 1998, Podestá et al. 2000, Teta et al. 2006a, Teta & Pardiñas 2007, Udrizar Sauthier & Carrera 2013).
Las egagrópilas son aglomerados de restos de presas no digeridos, como huesos, pelos, plumas, exoesqueletos de artrópodos, entre otros, consumidas por aves y regurgitadas periódicamente, en general en sitios de descanso y anidación (Errington 1930, Andrews 1990). La recolección y estudio de éstas, resulta un método adecuado en el estudio de presas pequeñas, debido a que permite el estudio de áreas extensas, genera gran cantidad y variedad de individuos por lote. En varias contribuciones, se ha considerado que el número y frecuencia de presas halladas en las egagrópilas son suficientemente representativas de la disponibilidad de presas en una determinada área (Errington 1930, Bonvicino & Bezerra 2003, Millán de la Peña et al. 2003, Pardiñas et al. 2003, Trejo & Lambertucci 2007, Gonzalez-Fischer et al. 2012, Lyman 2012).
El orden Strigiformes cuenta con el mayor número de estudios de dieta de aves rapaces a nivel mundial (Bó et al. 2007). En esta rama de la ecología, los estudios tróficos en aves rapaces son realizados comúnmente para la familia Tytonidae, y se destaca a la lechuza de campanario (Tyto furcata) por estar entre las especies con mayor número de contribuciones sobre el análisis de sus regurgitados (Gonzáles-Calderón 2017).
Tyto furcata es una especie que habita en gran variedad de ambientes, incluso muy antropizados (Tylor 2004). El estudio de las regurgitaciones de esta especie ha ayudado a realizar nuevas interpretaciones sobre la distribución, taxonomía y conservación de muchas especies de roedores (Massa et al. 2014a). Por ejemplo, se ha registrado que la variación de la densidad de una especie presa en ecosistemas templados puede correlacionarse con la proporción de esa especie presa en la dieta de Tyto furcata, e incluso dependiendo de la densidad de otras especies presentes en el área (Bernard et al. 2010). En general, estos mamíferos constituyen su principal fuente de alimentos, aunque también se alimentan de pequeños marsupiales y murciélagos (Massoia 1988, Massa et al. 2014b, Idoeta et al. 2020), y en menor proporción de pequeñas aves, anuros y reptiles (Massoia & Fornes 1964).
Holochilus chacarius es una especie de roedor cricétido, que muestra una distribución por la zona chaqueña de Argentina, Paraguay, Bolivia y parte del Cerrado de Brasil, llegando hasta el delta inferior de la desembocadura de los ríos Paraná y Uruguay (Massa et al. 2019, Prado et al. 2021). Esta especie no presenta registros en las cuencas fluviales del interior de las provincias de Buenos Aires y Santa Fe, donde sí aparece H. vulpinus (Torres & Massa 2019).
En este trabajo se utiliza el análisis de dieta de Tyto furcata como método indirecto, pero con alto grado de fiabilidad, para establecer la presencia de Holochilus chacarius en distintas localidades del sur y oeste de la provincia de Santa Fe. El objetivo de la presente contribución es presentar nuevos registros de H. chacarius para la provincia de Santa Fe, en un área donde previamente no había sido registrado, y discutir las posibles hipótesis acerca de la presencia de ésta y la ausencia de H. vulpinus en los muestreos.
Materiales y métodos
Área de estudio. - La provincia de Santa Fe, Argentina, se ubica en la región del centro-este del país, en una zona de clima templado con estacionalidad marcada, observando variaciones relacionadas a su extensión territorial con predominio de su eje norte-sur, que determina que en el norte el clima se aproxima más a un perfil subtropical, mientras que al sur es más bien de tipo templado-frío (Lewis & Collantes 1974, Coronel & Sacchi 2006).
La zona de estudio se ubica en el tercio más austral de su territorio con precipitaciones que oscilan entre los 800 y 1000 mm anuales, ubicándose entre las isotermas anuales de entre los 16 y 18 °C, variables entre días con heladas en época invernal y temperaturas estivales que pueden alcanzar más de 35 °C (Coronel & Sacchi 2006). Se corresponde con el bioma denominado “pampa húmeda argentina”, caracterizado por ser una extensa planicie con escasa pendiente que, originalmente fue un ecosistema de pastizal prácticamente continuo, sin especies leñosas excepto en algunos corredores, asociados principalmente a los cuerpos de agua de tipo lótico (ríos y arroyos) (Cabrera 1976).
En la actualidad presenta un grado de subdivisión y utilización del suelo que modificó su aspecto original, convirtiéndose en una planicie donde predominan los cultivos de granos y oleaginosas. En general, se puede considerar a la zona de estudio como un área de alta producción agrícola, con un crecimiento en detrimento de la ganadera y en una sobreexplotación del suelo, con prácticas intensivas de dobles cultivos anuales, como por ejemplo trigo y soja. Esto ha provocado una disminución importante de los ambientes naturales, un cambio en la estructura y funcionamiento de los ecosistemas y en la mayoría de los casos su fragmentación (Martínez 2010, Bilenca et al. 2012).
Para esta investigación se establecieron distintos puntos de muestreo (a partir de la presencia de perchas activas de Tyto furcata) en siete localidades del sur de Santa Fe. De esta manera quedó establecido para el departamento Caseros las localidades de Casilda, Los Molinos, Arequito, Los Nogales y Los Quirquinchos; para el departamento Iriondo, la localidad de Totoras; y para el departamento Belgrano, la localidad Montes de Oca.
A fin de establecer asociaciones entre los registros de presencia y el tipo de ambiente se estableció una categorización según uso del suelo siguiendo a Rimoldi y Curti (2021), el cual quedó establecido de la siguiente manera: Ambiente Urbano (U), donde hay dominancia de viviendas humanas, y los espacios verdes se reducen a los patios de las mismas y a las veredas donde se disponen árboles y arbustos mayoritariamente exóticos; Ambiente Rural Antropizados (RA), donde la producción agrícola crece en detrimento de la ganadería con una sobreexplotación del suelo con prácticas intensivas de dobles cultivos anuales, como por ejemplo maíz y soja; y Ambiente Rural No Antropizado o Seminatural (RNAN), donde las limitantes edáficas o geomorfológicas permiten la presencia de formaciones vegetales autóctonas sin agricultura o cría intensiva de ganado.
Colecta de datos. - Entre enero y octubre de 2020 se colectaron egagrópilas mensualmente en puntos previamente establecidos en los sitios de muestreo. En todos los casos se recogió el total de estas dejando la percha limpia. Tratándose de ámbitos restringidos, resultó relativamente sencillo rastrear en toda la superficie expuesta bajo las perchas, obteniéndose todo el material disponible.
Las egagrópilas se colocaron en bolsas de papel rotuladas (con el nombre del ambiente relevado, coordenadas, fecha y cantidad) y luego en bolsas de polietileno herméticamente selladas. Se respetaron todas las medidas de bioseguridad (uso de guantes de látex y barbijo en la colecta) hasta el secado del material, el cual se realizó en laboratorio con una estufa, a 70 °C durante 48 h (Muñoz-Pedreros & Yañez 2004). Posteriormente fueron procesadas, utilizando instrumental quirúrgico para extraer los restos de mandíbulas y cráneos de los animales encontrados. Aquellas egagrópilas que estaban compactadas fueron remojadas en agua durante tres horas para realizar la separación.
Las mandíbulas y cráneos de las presas capturadas se compararon con muestras identificadas en distintas colecciones osteológicas y literatura especializada (Voglino et al. 2004, Gómez Villafañe et al. 2005, Courtalon et al. 2013, Patton et al. 2015, Brandao & Nascimento 2015). Se consideró como un individuo a los pares de mandíbulas de la misma especie y/o cráneo.
Los materiales estudiados están depositados en la Colección de Zoología General de la Facultad de Ciencias Agrarias, Universidad Nacional de Rosario.
Resultados
Se obtuvo un total de 6236 individuos recuperados de 2598 egagrópilas. Del total, el 96% pertenece a micromamíferos, mientras que el 4% restante corresponde a ejemplares de anfibios, reptiles y aves. El 91% (n = 5448) del total de micromamíferos recuperados, correspondieron a roedores nativos de la familia Cricetidae, seguida por los roedores introducidos de la familia Muridae n = 476 (7.96%). La menor proporción de presas estuvo representada por las familias Caviidae n = 45 (0.76%), Molossidae n = 6 (0.1%) y Leporidae (0.016%).
A partir de los resultados obtenidos se pudo establecer la presencia de 31 individuos de Holochilus chacarius brindando de esta forma los primeros registros de esta especie para los departamentos Caseros, Iriondo y Belgrano (provincia de Santa Fe, Tabla 1 y Figura 1).
Tabla 1 Localidades muestreadas en el sur de las provincias de Santa Fe. Se indican, en cada caso, el número de individuos (n) de Holochilus chacarius y el tipo de ambiente en el cual se obtuvieron los registros. Referencias: RU: Rural antropizado; RNAN: Rural no antropizado o natural; U: Urbano.
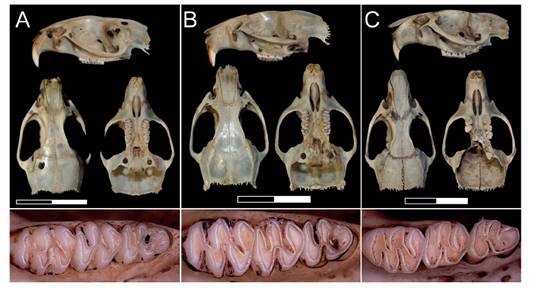
Figura 1 Restos craneanos de Holochilus chacarius hallados en distintas localidades de la Cuenca del Río Carcarañá, en vistas lateral izquierda, dorsal, ventral, y detalle de la serie molar derecha. Los ejemplares mostrados corresponden a las localidades de 1, 6 y 9 de la Figura 2. Es decir, A, individuo de Casilda; B, individuo de Los Quirquinchos; C, individuo de Montes de Oca 2. Escalas: 20 mm.
Los restos craneales fueron asignados a Holochilus chacarius dada la presencia de la siguiente combinación de caracteres: 1) mesolofo ausente en los molares superiores/inferiores; 2) áreas de los proto e hipocono de contorno subrectangular (Hershkovitz 1955, Massoia 1976, Pardiñas & Galliari 1998, Voglino et al. 2004, Pardiñas et al. 2013). Además, las medidas tomadas a los individuos (ver Tabla 2) se corresponden con las indicadas por otros autores para esta especie (Pardiñas et al. 2013, Brandao & Nascimento 2015). Los ejemplares colectados en los distintos puntos de muestreo (ver Tabla 1) distan entre 50 Km y 115 Km al oeste del área de distribución conocida para la especie (ver Figura 2).
Tabla 2 Medidas craneodentarias (en mm) de los individuos de Holochilus chacarius mostrados en la Figura 1. M, media; DE, desvío estándar. Abreviaturas de las medidas (tomadas de Brandao & Nascimento, 2015): LD, longitud del diastema; MTR, longitud de la serie molar; BM1, ancho del M1; LIF, longitud del foramen incisivo; BIF, ancho del foramen incisivo; BZP, ancho de la placa zigomática; LIB, ancho interorbitario.
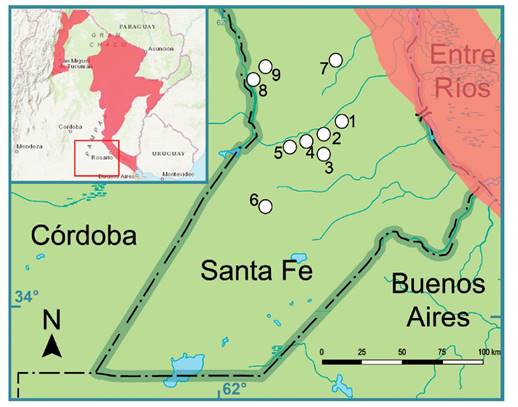
Figura 2 Área de estudio. En la esquina superior izquierda del recuadro se indica en rosado el área de distribución reconocida para la Holochilus chacarius en Argentina, según la última categorización nacional (Massa et al. 2019). El recuadro rojo indica el área de estudio. Los puntos numerados indican las localidades referenciadas en la Tabla 1.
Discusión
Los registros aquí presentados extienden el rango de distribución de H. chacarius por hasta 115 Km (Montes de Oca) hacia el oeste del río Paraná. Además, no se obtuvo ningún registro de H. vulpinus.
El río Paraná y sus grandes afluentes, como el Paraguay, son importantes “corredores” (es decir, rutas efectivas para la migración activa o pasiva) de la flora y fauna tropical hacia zonas templadas, donde pueden coexistir con especies locales (Ringuelet 1961). En el área estudiada existe un importante cuerpo de agua, el río Carcarañá, donde en sus riberas hay una mezcla de ambientes propios de pastizal pampeano (espartillares, ambientes halófitos, etc.), parcialmente modificados por actividad antrópica (mayormente áreas de cultivos y campos ganaderos), y ambientes boscosos típicos de la Ecorregión del Espinal. En este aspecto, H. chacarius y H. vulpinus han sido registrados en distintos sectores de la llanura Chaco-pampeana, a ambientes con alta diversidad de paisajes (Massa et al. 2014a), y ambas especies han sido halladas en egagrópilas de Tyto furcata, procedentes de agroecosistemas o cercanas a ellos (Scheibler & Christoff 2004, Teta et al. 2010, Gonzalez-Fisher et al. 2012, 2017, Massa 2015).
Aunque el área de distribución de ambas especies es parcialmente simpátrica, son muy raras las referencias sobre hallazgos de ambos taxones en la misma localidad (Contreras et al. 2003, Voglino et al. 2004, Teta & Pardiñas 2010, Massa et al. 2014a). Además, Prado et al. (2021) plantearon que las especies de Holochilus no sólo están separadas genéticamente, sino también ambientalmente, diferenciando a H. vulpinus como típicamente de pampas, mientras que H. chacarius sería más frecuentes en ambientes tipo pantanal. Por lo tanto, aunque en los mapas de distribución parecen ser especies simpátricas en distintos puntos de sus áreas de distribución, es probable que frecuenten diferentes ambientes. En los reportes de ambas especies en la misma localidad, en general H. vulpinus suele ser muy abundante, mientras que H. chacarius está poco representada (Chebez et al. 2005). Sólo en localidades del Chaco Húmedo se ha observado la tendencia contraria (Massoia 1971, 1976, Teta & Pardiñas 2010, Udrizar Sauthier & Carrera 2013), donde H. chacarius parece ser más abundante que H. vulpinus, y en pocas localidades se han reportado ambas especies en cantidades similares (Voglino et al. 2004). Por otro lado, son muy abundantes las citas de una de las dos especies, y la ausencia de la otra, particularmente cuando la muestra presenta una gran cantidad de individuos de una de las dos especies (Massoia et al. 1989, 1995, 1997, 1999, Teta & Contreras 2003, Teta et al. 2006b, Massa et al. 2014a, Brandao & Nascimento 2015).
Observando las áreas de distribución de H. vulpinus y H. chacarius, puede verse que esta última muestra una distribución por la zona chaqueña de Argentina, Paraguay, Bolivia, y parte del Cerrado de Brasil. Por otro lado, H. vulpinus parece habitar en un rango mayor de paisajes, ya que sus registros van desde el sur de la provincia de Buenos Aires, por el este de Argentina, hasta Paraguay y el sur de Brasil (Weksler et al. 2016a, 2016b). Esto es importante, porque según la última categorización de mamíferos de Argentina, H. vulpinus debería encontrarse en la zona centro y sur de la provincia de Santa Fe (Torres & Massa 2019), mientras que H. chacarius llegaría hasta el delta inferior del Paraná en la provincia de Buenos Aires, a través de la cuenca del río Paraná (Massa et al. 2019). En los registros aquí presentados, se muestra que en el centro-sur de Santa Fe, atravesando de oeste a este el ancho de la provincia habita H. chacarius, y no hay reportes concretos de H. vulpinus.
Se ha planteado que H. chacarius y H. vulpinus habrían sufrido retracciones en sus áreas de distribución en las últimas décadas, posiblemente en asociación al cambio climático por actividad antrópica (Teta et al. 2014). Los nuevos hallazgos aquí reportados podrían deberse a la ausencia de muestreos detallados previos, como se ha sugerido para otras áreas donde se han reportado gran cantidad de ejemplares de H. chacarius (Voglino et al. 2004, Courtalon et al. 2013).
En conclusión, los registros que aquí se suman de H. chacarius, extienden su área de distribución en la provincia de Santa Fe, casi hasta el límite con la provincia de Córdoba. La intensificación de muestreos en áreas poco exploradas, con nuevas técnicas de muestreo y de identificación de taxones, ayudarán al mejor conocimiento de estos taxones y otros grupos con problemas similares.

