











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Senecio L. es uno de los géneros de angiospermas más ricos en especies (Nordenstam et al. 2009), aproximadamente 1250 (Bremer 1994, Pelser et al. 2007, Nordenstam 2007), la mayor parte repartidas por los Andes, surcentro de África, la cuenca Mediterránea y este de Asia (Pelser et al. 2007, Nordenstam et al. 2009, Calvo et al. 2015). Esta enorme zonación indica la complejidad del género en cuanto a su variabilidad morfológica (Salomón et al. 2018), desde hierbas anuales a perennes rizomatosas, subarbustos y arbustos. Sus principales caracteres los podemos encontrar en los capítulos con el involucro campanulado o cilíndrico con una sola fila de brácteas, generalmente con brácteas suplementarias; anteras con el conectivo prolongado en un apéndice cuspidado y la base con un collar generalmente balaustriforme; el estilo acaba en dos ramas lineares truncadas con una franja de tricomas penicilados, y en las que el área estigmática forma dos bandas longitudinales estrechas (Vision & Dillon 1996, Calvo & Aedo 2019).
En cuanto a su distribución, las especies de Senecio se pueden encontrar desde áreas desérticas, comunidades vegetales de bajas y medias altitudes, formando parte de matorrales, y comunidades de alta montaña (Beltrán & Galán de Mera 1996, 1998, Pelser et al. 2007, Montesinos- Tubée et al. 2021), características ecológicas que han sido tratadas en floras y catálogos florísticos atendiendo solamente a su distribución altitudinal y a zonas de vegetación muy amplias, aunque con una actualización taxonómica (Cabrera 1957 Brasil, Paraguay y Uruguay, Vision & Dillon 1996, Beltrán et al. 2007- Perú, Robinson 1999 Ecuador, Beck & Ibáñez 2014- Bolivia, Freire et al. 2014- Argentina, Ávila et al. 2015-Colombia, BFG 2015- Brasil, Rodríguez et al. 2018, Zuloaga et al. 2019- Chile).
En el Perú, las especies del género se distribuyen de norte a sur, con diferentes rangos altitudinales y una alta endemicidad ya observada en algunos trabajos locales (Galán de Mera & Linares Perea 2012, Beltrán & Roque 2015, Beltrán 2016, Gonzáles 2016, Whaley et al. 2019), y representada por muchas de las especies descritas recientemente, como S. carhuanishoensis H. Beltrán & Calvo (Beltrán & Calvo 2019), S. beltranii P. Gonzáles & Montesinos- Tubée (Montesinos-Tubée & Gonzáles 2020), S. huaynaputinaensis Montesinos-Tubée & Chicalla-Rios (Montesinos-Tubée & Chicalla-Rios 2021) o S. phlomidifolius H. Beltrán (Beltrán & Calvo 2021). Hasta el momento, no conocemos investigaciones sobre las relaciones biogeográficas entre las especies de este género, cuáles son las barreras de su distribución, qué especies son más frecuentes a lo largo del territorio peruano, y cómo se relacionan con el relieve y bioclimatología andinas.
Puesto que las especies se distribuyen a lo largo de la cordillera de los Andes, y están repartidas entre las distintas unidades de vegetación, parece evidente la influencia del cambio climático sobre su distribución y conservación. Por ello, además, incluimos observaciones sobre las modificaciones que se producirían entre las distintas cinturas de vegetación, y cómo afectarían a las especies de Senecio.
Material y métodos
Colecciones. - El presente trabajo está basado en las colectas del primer autor, depositadas en el herbario USM y la consulta de los principales herbarios donde se conserva material del Perú (B, CPUN, F, HUT, K, MO, NY, P, US, USM; abreviaturas de acuerdo con Thiers 2022). Además, se consultó la base de datos del portal GBIF (2022), que toma información de la mayor parte de los herbarios. Con todos estos datos, se obtuvo una matriz de 5263 pliegos de herbario, de los cuales en 4342 se pudo confirmar la localidad con coordenadas en grados decimales y la especie de Senecio que contienen, con lo que podemos conocer el rango altitudinal de cada especie, así como su distribución departamental y regional en los países adyacentes de América del Sur. Con esta matriz se obtuvo la tabla que se detalla en el Apéndice 1.
Bioclimatología. - Para conocer la influencia de distintas variables climáticas sobre la distribución de las especies de Senecio se usó el modelo de los pisos bioclimáticos de Rivas-Martínez (Rivas-Martínez et al. 1999), analizados y cartografiados en el Perú por Galán de Mera et al. (2017). Los pisos bioclimáticos están basados en intervalos del Índice de Termicidad, que suma diferentes valores de temperatura en grados Celcius, (It = (T + M + m) 10; T = temperatura media anual, M = temperatura media de las máximas del mes más frío, m = temperatura media de las mínimas del mes más frío) en conjunción con plantas bioindicadoras, que guardan una tendencia hacia un piso bioclimático, y comunidades vegetales. En el Perú reconocemos 6 pisos bioclimáticos: Infratropical (It > 690), Termotropical (It 491 a 690), Mesotropical (It 321 a 490), Supratropical (It 161 a 320), Orotropical (It 50 a 160), y Criorotropical (It < 50). Los pisos bioclimáticos se matizan con intervalos de precipitaciones (P anual en mm) ‒Ultrahiperárido (P < 5), Hiperárido (P 5 a 30), Árido (P 31 a 100), Semiárido (P 101 a 300), Seco (P 301 a 500), Subhúmedo (P 501 a 900), Húmedo (P 901 a 1500), Hiperhúmedo (P 1501 a 2500), y Ultrahiperhúmedo (P > 2500)‒ de tal forma que podemos encontrarnos con un piso termotropical muy seco o muy húmedo. Los valores de temperatura y precipitación fueron obtenidos del Servicio Nacional de Meteorología e Hidrología del Perú (SENAMHI).
Los pisos bioclimáticos se pueden asociar a las características climáticas y tipos de vegetación del modelo de Köppen-Geiger (Kottek et al. 2006, Rubel & Kottek 2010). El mapa de Köppen-Geiger es posible visualizarlo con el portal Google Earth (http://koeppen-geiger. vu-wien.ac.at/shifts.htm) a partir de datos climáticos entre 1986 y 2010, y a una resolución de 5 arcos de minuto, y además establecer hipótesis de cambio climático hasta el año 2100 (Climate Change & Infectious Diseases Group 2019).
Análisis estadístico. - Los análisis estadísticos han sido realizados a partir de los datos numéricos del Apéndice 1 con el programa PAST 4.07b (Hammer et al. 2001), donde figuran las especies de Senecio, su distribución departamental y en países de alrededor en caso de no ser endemismos peruanos, rango altitudinal medio, variables climáticas medias e Índice de Termicidad medio. Con el rango altitudinal podemos conocer la altitud media (Hm en m sobre el nivel del mar) a la que crece una especie, y además las estaciones meteorológicas más próximas a una determinada especie en un departamento del Perú, y con ellas los valores de P, M, m e It. Dichas estaciones fueron tomadas de Galán de Mera et al. (2017). Los valores medios de las estaciones para una especie (Pm, Mm, mx e Itm) ofrecen su tendencia como bioindicador de un piso bioclimático junto con un valor de precipitaciones.
La presencia o ausencia de las especies de Senecio en los distintos departamentos del Perú, y la similitud existente entre ellos sirve para apreciar la distribución geográfica y, por tanto, conocer algunas sus características biogeográficas en cuanto a la proximidad entre áreas (Galán de Mera et al. 1997, Freitas et al. 2019, Hansen et al. 2021). Para ello se construyó un clúster basado en valores de similitud por pares (UPGMA) utilizando el coeficiente de Sørensen para presencia-ausencia entre los diferentes departamentos del Perú.
Para observar la tendencia de cada especie en relación con la altitud (Hm) y las variables e índices bioclimáticos (Pm, Mm, mx e Itm) se realizó un análisis de componentes principales (ACP).
En los análisis estadísticos no se incluye a S. vulgaris L. por ser una especie introducida con una gran amplitudaltitudinal, de 50 a 4000 m, que crece en zonas antropizadas. Tampoco a S. tridentatus DC., que solo se conoce deuna colecta de Haenke (P) con el departamento dudoso.
Resultados
Análisis estadístico y cartografía. -El género Se-necio en Perú está conformado por 153 especies, de las cuales 94 son endémicas.
La Figura 1 muestra el análisis de agrupamientobasado en los registros de las especies de cada departamento. Lambayeque (LA) no tiene ninguna especiepropia, pero su rama se encuentra unida a la de Piura(PI), que contiene a S. josei Sklenář (especie común conEcuador, ver Apéndice 1) y S. otophorus Wedd. (distribuida entre Venezuela y Perú). Ambos departamentos son próximos a Cajamarca (CA), La Libertad (LL),Amazonas (AM) y San Martín (SM), donde se distribuye un grupo de especies que no alcanzan la CordilleraBlanca hacia el sur, como S. piurensis Sagást. & Zardini (endemismo de Cajamarca y Piura), S. woodsonianus Cuatrec. (endemismo de Amazonas), S. involucratus DC. (que penetra desde Colombia hasta el departamento deAmazonas), S. recurvatus Kunth (endemismo de Caja-marca), S. ishcaivilcanus Cuatrec. (endemismo de Caja-marca y La Libertad), S. laricifolius Kunth (endemismode Amazonas, Cajamarca, La Libertad y San Martín), S. szyszylowiczii Hieron. (endemismo de Amazonas, Caja-marca y Piura) y S. jungioides Cabrera (endemismo deAmazonas, Cajamarca y La Libertad). Las ramas de Arequipa (AR), Moquegua (MO) y Tacna (TA) recogen especies sobre todo occidentales del sur del Perú, aunque con algunas disyunciones interiores (Fig. 2A), como S. trifurcifolius Hieron. (que se adentra desde Chile y Bolivia hasta Arequipa y algunas localidades de Cusco), S. beltranii (endemismo de Arequipa y Cusco), S. ticsanicus Montesinos & Trinidad (endemismo de Moquegua), S. tacorensis Cabrera (que alcanza Tacna desde Bolivia), S. behnii Ricardi & Martic. (que se adentra hastaMoquegua desde Chile), S. phylloleptus Cuatrec. (que sedistribuye por la puna seca de Bolivia, Chile, Arequipa,Moquegua y Tacna), S. cumingii Hook. & Arn. (que crece en Chile y Moquegua), S. neoviscosus Cuatrec. (endemismo de Arequipa y Moquegua), y S. ctenophyllus Phil. (deChile y Tacna).
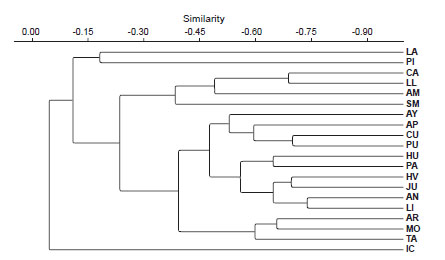
Figura 1: Dendrograma para mostrar la similitud de presencia-ausencia de las diferentes especies de Senecio entre los departamentos del Perú. El clúster está basado en valores de similitud por pares (UPGMA) usando el coeficiente de Sorensen. Las abreviaturas departamentales coinciden con Lamas & Encarnación (1976).
La rama del departamento de Ica (IC) se encuentraseparada al abarcar especies termotropicales, como S. fernandinus H. Beltrán & J. Calvo (endemismo de Ica), aveces compartidas, con Arequipa, en el caso de S. calcicola Meyen & Walp. (endemismo de Arequipa e Ica), o conáreas más interiores de Ayacucho (AY) o Huancavelica(HV), como S. acarinus Cabrera (endemismo de Arequipa,Ayacucho, Huancavelica e Ica). Las ramas de Ancash (AN),Junín (JU) y Lima (LI) también aparecen juntas debido aalgunas especies de distribución circunscrita a la Cordillera Blanca y a la cordillera de La Viuda, como ocurre con S. pickeringii A.Gray (endemismo de Ancash, Junín y Lima,que también alcanza Ayacucho), S. casapaltensis Ball (endemismo de Ancash, Junín y Lima) (Fig. 2B), S. rauranus Cuatrec. (endemismo de Ancash), S. rauhii Cuatrec. (endemismo de Ancash), S. tubicapillosus Cuatrec. (endemismo de Ancash), S. carhuanishoensis (endemismo de Lima), y S. phlomidifolius (endemismo de Lima).
El resto de las especies están repartidas por la puna húmeda, como ocurre con S. melanandrus (Wedd.) J.Calvo, A.Granda & V.A.Funk (que se encuentra también en Bolivia), S. burkartii Cabrera (que sube desde Argentina hasta Piura y San Martín), S. betonicifolius DC. (cuya distribución se extiende desde Ecuador a Bolivia), S. nutans (que se distribuye por Argentina, Bolivia, Chile y Perú pero no supera la Cordillera Blanca hacia el norte) (Fig. 2B), y S. hebetatus Wedd. (endemismo de Ayacucho, Cusco y Junín). Dos ramas orientales bastante bien definidas en el dendrograma (Fig. 1) son las de Apurímac (AP) y Cusco (CU), que comparten especies como S. panticallensis Cabrera (endemismo de Apurímac y Cusco) y S. tenuicaulis Sch.Bip. ex Klatt (también presente en Bolivia); la rama de Cusco queda bien representada por endemismos como S. qosqoensis H. Beltrán & A. Granda, S. parvocapitatus Cabrera y S. calcensis Cabrera & Zardini.
Al observar la Figura 1 también podemos deducir que la mayor parte de las especies de Senecio en el Perú se ubican en los departamentos que comparten la puna húmeda.
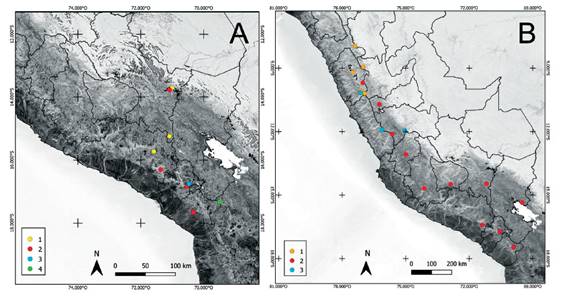
Figura 2: Distribución de algunas especies de Senecio: A) especies criorotropicales del sur del Perú, en localidades con inviernos muy fríos, 1. S. beltranii, 2. S. trifurcifolius, 3. S. ticsanicus, 4. S. tacorensis, B) especies supratropicales de localidades con inviernos fríos, 1. S. sulinicus, 2. S. nutans, 3. S. casapaltensis.
En la Figura 3A se representa el número de especies por departamento que comparte el Perú con otros países, el número de endemismos nacionales (endemismos peruanos compartidos por varios departamentos), y el número de endemismos exclusivamente departamentales. Mientras que la Figura 3B indica la frecuencia de especies por departamento. La puna húmeda es un bioma andino caracterizado por grandes espacios de Gramíneas situados por encima de los 3500 - 3800 m, ubicado sobre todo en el centro del país y entorno del lago Titicaca (Galán de Mera et al. 2017), donde se distribuye la mayor parte de las especies de Senecio (Fig. 4A). Esto coincide con una mayor distribución supra- y orotropical (Fig. 4B), como también se demuestra en el ACP (Fig. 5).
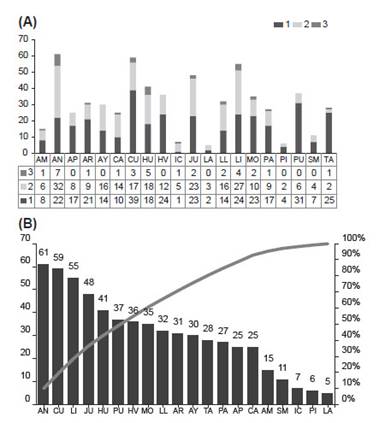
Figura 3: Diversidad y endemicidad de Senecio por departamento: A) 1. Especies que exceden el territorio peruano, 2. Endemismos nacionales, 3. Endemismos departamentales. B) Distribución de especies en orden decreciente de frecuencia por departamento. La línea significa el porcentaje de acumulación de especies.
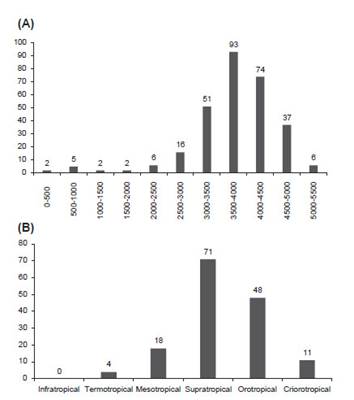
Figura 4: Diversidad altitudinal y bioclimática de Senecio en el Perú: A) Número de especies por rango altitudinal (intervalo de 500 m), B) número de especies por piso bioclimático.
En el ACP de la Figura 5 se muestra la correlación entre las especies de Senecio con la altitud media (Hm), valores medios de variables climáticas (Pm, Mm, mx) y del Índice de Termicidad (Itm), y distribución geográfica. Podemos reconocer 10 grupos de especies:
- A- Son especies criorotropicales del S del Perú, con inviernos muy fríos (mx con valores muy bajos de alrededor de -12 °C), y ambientes subhúmedos de la puna seca por debajo de 600 mm de precipitación (Fig. 2A),
- B- Son también especies criorotropicales con inviernos muy fríos (mx con valores de alrededor de -7 a -10 °C) y ambientes más húmedos correspondientes a la puna húmeda.
- C, D y E- Son grupos supratropicales. El piso supratropical está diversificado en tres grupos, pues es donde se encuentra el mayor número de especies (Fig. 4B). C. Son las del norte del Perú que no alcanzan hacia el sur de la Cordillera Blanca, y se ubican en áreas con un intervalo de lluvias húmedo. Senecio minesinus Cuatrec. solo alcanza una Pm de 892.8 mm (intervalo subhúmedo), siendo muy cercana en el gráfico a S. ayapatensis Baker que ya se encuentra además en la Cordillera Blanca. D. Son las especies localizadas en áreas con inviernos más templados, donde mx es positiva, y esto coincide con una distribución andina oriental. E. Es un grupo bastante heterogéneo que abarca desde especies localizadas en lugares húmedos, como S. casapaltensis (Pm 1055 mm), como en lugares secos, como S. hebetatus (Pm 445,5 mm), con mx negativa (Fig. 2B). La parte de E que está situada por debajo del eje de abscisas está compuesta por especies de lugares con bajas precipitaciones, de intervalo seco-subhúmedo (Pm alrededor de 500 - 600 mm), donde la temperatura máxima del mes más frío (Mm) es más elevada, aunque los valores de las temperaturas mínimas (mx) se mantienen negativos. Suelen ser especies propias de depresiones interandinas e incluso de zonas occidentales, como S. yurensis Rusby. En general, las especies que se encuentran próximas entre un grupo y otro, pertenecen a distintas áreas biogeográficas, piso bioclimático, como S. behnii y S. neoviscosus, o intervalo de precipitaciones, como S. minesinus (dentro del intervalo subhúmedo) y S. ochoanus (dentro del intervalo húmedo).
- F- Son todas las especies orotropicales, situadas a la izquierda del eje de ordenadas, donde las podemos observar en áreas hiperhúmedas (Pm > 1600 mm), como S. pascoensis Cabrera, a áreas subhúmedas (Pm < 700 mm), como S. mathewsii Wedd. Entre ellas, además podemos observar especies muy dispersas por el territorio, como S. evacoides Sch.Bip. que no supera la Cordillera Blanca hacia el N, S. chachaniensis Cuatrec., que la encontramos distribuida por el S del Perú, o S. humillimus Sch.Bip., de la zona oriental, pero que alcanza las montañas de Arequipa, Moquegua y Tacna. Por el contrario, hay especies muy restringidas, como S. qosqoensis en Cusco, S. pascoensis en Pasco, S. crassiandinus Montesinos & Pino y S. roseoandinus Montesinos & R. Zárate en Huánuco.
- G- Son especies mesotropicales que no superan hacia el sur de la Cordillera Blanca. S. jungioides y S. laricifolius son de ambientes subhúmedos (Pm ~ 700 mm) sobre todo orientales en los departamentos de Amazonas, Cajamarca, La Libertad y San Martín.
- H- Son especies de Senecio mesotropicales que superan o se localizan al sur de la Cordillera Blanca. Senecio axillifoliatus Cuatrec., S. lancidentatus Cuatrec. y S. weberbaueri Cuatrec. son de ambientes subhúmedos orientales (Pm > 500 mm), mientras que S. gracilipes A. Gray y S. huaynaputinaensis son de localidades occidentales secas (Pm < 500 mm).
- I- Son igualmente mesotropicales, pero de los ambientes semiáridos que se alcanzan en el sur del Perú. Senecio acarinus llega a desarrollarse a un intervalo de precipitaciones casi árido (Pm ~ 150 mm).
- J- Reúne especies termotropicales. S. calcicola, S. fernandinus y S. smithianus son de las lomas occidentales del desierto peruano, mientras que S. inghamii Montesinos es de valles térmicos semiáridos del centro- norte del país.
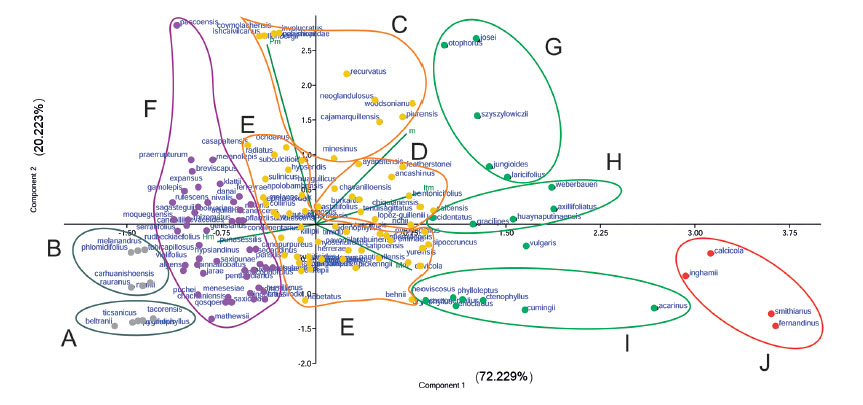
Figura 5: Análisis de Componentes Principales (ACP) correlacionando las especies de Senecio con la altitud media (Hm), y valores medios de variables e índices climáticos (Pm: precipitación anual media, Mm: valor medio de las temperaturas máximas del mes más frío, mx: valor medio de las temperaturas mínimas del mes más frío, Itm: índice de termicidad medio). Los porcentajes entre paréntesis junto a las componentes significan la varianza comprendida por el eje respectivo. Componente 1: frío a cálido, componente 2: seco a húmedo. Agrupaciones de especies: A) Criorotropicales de la puna seca, B) criorotropicales de la puna húmeda, C) supratropicales que no superan hacia el sur la Cordillera Blanca, D) supratropicales de localidades con inviernos templados, E) supratropicales de localidades con inviernos fríos, F) orotropicales, G) mesotropicales que no superan hacia el sur la Cordillera Blanca, H) mesotropicales que superan hacia el sur la Cordillera Blanca, I) mesotropicales de ambientes semiáridos del sur del Perú, J) termotropicales.
Cambio climático y conservación. - La distribución de especies de Senecio en diferentes pisos bioclimáticos (Fig. 4B), sobre todo en el supra- y orotropical, sugiere la posibilidad de cambios ante el calentamiento global. En la Figura 6 se muestra el reparto de especies en el piso orotropical en mapas con la correspondencia entre el modelo de Köppen-Geiger y el de los pisos bioclimáticos (Fig. 6A), en el 2020 (Fig. 6B), dentro de 50 años en el 2070 (Fig. 6C), y en el 2099 (Fig. 6D). Se puede observar que dentro de 50 años los hábitats que ocupaban las especies orotropicales y, por tanto, las criorotropicales presentes en la vegetación periglaciar, comenzarían a desaparecer, y a los 100 años prácticamente habrían desaparecido con la excepción de áreas de la Cordillera Blanca y del Coropuna. Esto además supone el ascenso de especies termo-, meso- y supratropicales hacia altitudes superiores, y el peligro de extinción de un buen número de endemismos, como S. chachaniensis, S. pascoensis, S. qosqoensis, S. ticsanicus, S. tubicapillosus, y en general de todas las especies localizadas en los pisos orotropical y criorotropical.
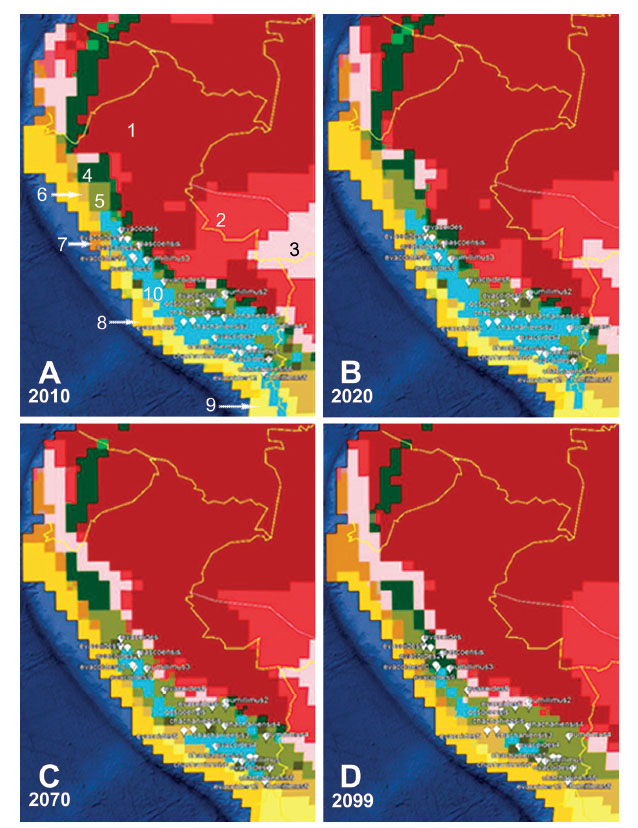
Figura 6: Alteración de los hábitats de las especies orotropicales de 0Senecio ante cambios climáticos. A) Simbología de Koppen-Geiger relacionada con los pisos bioclimáticos [1. Bosques ecuatoriales muy húmedos (infra- y termotropical hiperhúmedo y ultrahiperhúmedo), 2. Clima ecuatorial monzónico (infra- y termotropical húmedo-hiperhúmedo), 3. Sabana ecuatorial con inviernos secos (infra- y termotropical seco-subhúmedo), 4. Clima templado y cálido muy húmedo con verano fresco (mesotropical hiperhúmedo-ultrahiperhúmdo), 5. Clima templado y cálido con invierno seco y verano cálido (supratropical de seco a húmedo), 6. Clima de estepa frío y árido (supratropical de semiárido a seco), 7. Clima de estepa cálido y árido (termo- y mesotropical de árido a semiárido), 8. Clima desértico cálido y árido (infra- y termotropical hiperárido), 9. Clima desértico frío y árido (termotropical de ultrahiperárido a hiperárido), 10. Clima de tundra (oro- y criorotropical de seco a subhúmedo] y distribución de especies en los pisos bioclimáticos oro- y criorotropical en 2010. B) Distribución de especies con relación a los bioclimas oro- y criorotropical en la actualidad. C) Distribución de especies en los pisos bioclimáticos oro- y criorotropical en 50 años. D) Distribución de especies en los pisos bioclimáticos oro- y criorotropical en el año 2099 (Mapas obtenidos a partir del archivo kmz existente en http://koeppen-geiger.vu-wien.ac.at/shifts.htm visible con el portal Google-Earth, basado en cambios climáticos observados y previstos, según Rubel & Kottek 2010).
Discusión
La mayor parte de las especies de Senecio en el Perú se encuentran en los departamentos de la puna húmeda (Fig. 3), y guardan relación con las áreas del mapa de las provincias biogeográficas del Perú, que basadas en el mapa biogeográfico de América del Sur de Rivas-Martínez et al. (2011), establece tres áreas (Fig. 7): especies dentro de la provincia biogeográfica Guayaquileño-Ecuatoriana, que quedan al norte y no alcanzan la Cordillera Blanca, que se configura como una barrera biogeográfica hacia el sur para muchas especies (Weigend 2002, Galán de Mera et al. 2015); especies que definen la provincia de la Puna Xerofítica, como S. behnii, S. tacorensis y S. ticsanicus (Fig. 2A); un grueso de especies repartidas por la provincia de la Puna Mesofítica; y las especies del desierto en la provincia biogeográfica Norperuana Hiperdesértica. En la provincia biogeográfica de la Puna Mesofítica contrastan especies con un reparto muy amplio, como es el caso de S. melanandrus, con distribuciones muy restringidas, como es el caso de S. qosqoensis o S. roseoandinus (Montesinos-Tubée et al. 2018), lo que está de acuerdo con los postulados de las islas andinas (Sklenář et al. 2014), pues las montañas andinas originaron rutas migratorias durante los periodos fríos del Plio-Pleistoceno, cuando surgieron islas en las alturas que favorecieron la especiación en numerosos géneros, como Draba, Gentianella, Senecio y Valeriana (Sklenář & Balslev 2005); el enfriamiento pliocénico y los niveles de altitud alcanzados por los Andes dio origen a la vegetación de la puna, que con sucesivas expansiones y reducciones permitió la radiación de algunos géneros hacia latitudes medias, como Lupinus (Graham 2009), y posiblemente Senecio.
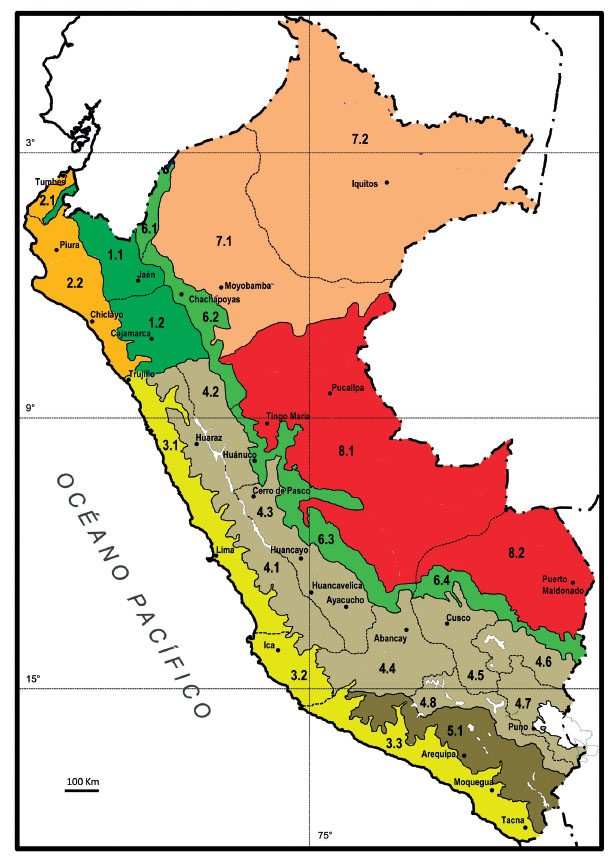
Figura 7: Mapa con las provincias y sectores biogeográficos del Perú, basado en Rivas-Martínez et al. (2011). Dentro del Reino Neotropical-Austroamericano y del Subreino Neotropical existen 4 regiones biogeográficas de las que participa el Perú: A) Región Neogranadina: 1. Provincia Guayaquileña-Ecuatoriana (1.1. Sector de Loja-Cutervo, 1.2. Sector de Chota-Contumazá), 2. Provincia Pacífica (2.1. Sector de Tumbes, 2.2. Sector de Sechura). B) Región Pacífica Tropical Hiperdesértica: 3. Provincia Norperuana Hiperdesértica (3.1. Sector de Lima, 3.2. Sector de Ica, 3.3. Sector de Moquegua-Tacna). C) Región Surandina Tropical: 4. Provincia Puneña Mesofítica (4.1. Sector de Huaraz, 4.2. Sector de Chavín, 4.3. Sector de Xauxa, 4.4. Sector de Apurimac, 4.5. Sector de Cusco, 4.6. Sector del Allincapac, 4.7. Sector de la Cuenca del Titicaca, 4.8. Sector del Colca), 5. Provincia Puneña Xerofítica (5.1. Sector de Arequipa), 6. Provincia Yungueña (6.1. Sector del Norte del Perú, 6.2. Sector de Chachapoyas, 6.3. Sector de Huánuco-Junín, 6.4. Sector del Urubamba). D) Región Amazónica: 7. Provincia Amazónica Occidental (7.1. Sector del Pastaza-Marañón, 7.2. Sector de Iquitos), 8. Provincia Amazónica Suroccidental (8.1. Sector de Contamana-Pucallpa, 8.2. Sector de Madre de Dios).
Las especies de la Puna Xerofítica se localizan en torno al arco volcánico que se inicia en Ayacucho y se adentra en Chile y Bolivia, pero algunas de ellas, como S. beltranii, S. trifurcifolius (Fig. 2A), S. chachaniensis o S. humillimus también aparecen hacia oriente. En estos departamentos volcánicos entre Ayacucho y Tacna encontramos una menor diversidad respecto a los de la Puna Mesofítica del centro (Fig. 3B); la localización muy lejana oriental de algunas especies tal vez sea debida a las formas anemócora y ectozoocora (Stuessy et al. 1996) de dispersión del fruto y a un cambio en la composición química del suelo a causa de la actividad volcánica (Schwarzer et al. 2010).
Los elementos termotropicales, como S. calcicola, S. fernandinus y S. smithianus se circunscriben a áreas precisas de la provincia biogeográfica Norperuana Hiperdesértica, en el sector de Ica, en el caso de S. fernandinus, y en el sector de Moquegua-Tacna, en el de S. calcicola y S. smithianus, como ya se señaló en una sectorización previa del desierto en atención a la flora y vegetación de las lomas (Galán de Mera et al. 1997).
Numerosos autores han estudiado en distintas partes del mundo cómo el calentamiento global afecta a la distribución y conservación de especies y ecosistemas hasta provocar su extinción (Halloy & Mark 2003, Thuiller et al. 2005, Dullinger et al. 2012). En los Andes, los resultados de otros autores muestran, al igual que con Senecio (Figura 6A-6D), una migración altitudinal de las especies con el riesgo de extinción por pérdida de los hábitats (Thomas et al. 2004, Feeley et al. 2011, Peyre et al. 2020), con problemas de conservación por su endemicidad (Funk et al. 2005). En este sentido, nuestros resultados con Senecio están de acuerdo con los de Gonzáles et al. (2018) en los Andes de Carabaya (Puno), donde señalan cómo el cambio climático asociado a influencias antrópicas conllevan la fragmentación de hábitats y a posibles extinciones en las cimas de las montañas. Asimismo, son acordes con Sklenář et al. (2021) en el páramo ecuatoriano, donde concluyeron que S. nivalis ascendió en el nevado Antisana de los 4600 m en 1880 a 4850 m en la actualidad, y de unos 4800 a 5300 m en el Chimborazo.

