











 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
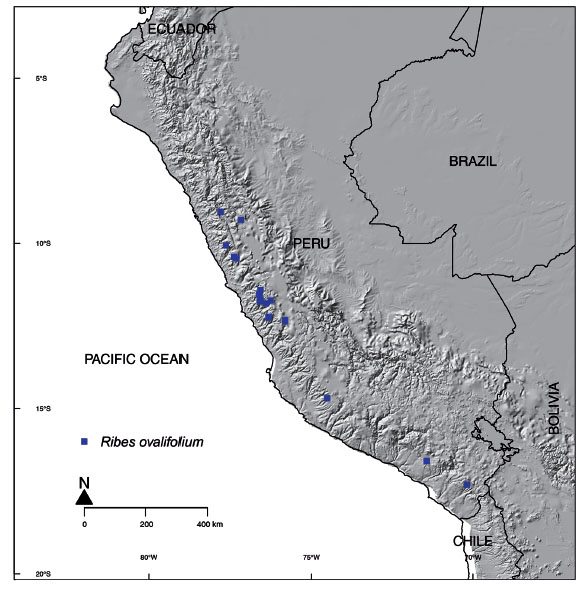
The genus Ribes is highly diverse in the northern hemisphere, especially Eastern Asia and western North America, but also has a considerable number of species in South America (Janczewski 1907). The most recent revision of the genus on a global basis was provided by Janczewski (1907) and the most recent revision for Peru was published by Macbride (1941). Recent studies have yielded a considerable number of new species from Peru, especially from northern Peru and adjacent Ecuador (Weigend & Rodriguez 2006; Weigend et al. 2005, 2010). The genus is most diverse in Andean cloud forest and sub-paramo and sub-puna habitats, but a small handful of species are also found at higher elevations or at least seasonally very dry habitats on the western slope of the Andes. These species have unusually small and - atypical for Ribes - virtually undivided leaves and are here informally assigned to the Ribes cuneifolium group. Ribes cuneifolium Ruiz & Pav. was originally described from high elevations of Junin, but has since been recorded along much of the Peruvian cordillera at elevations of 3500 to over 5000 m. A doubtfully distinct species, Ribes incertum J.F. Macbr., was described from Ancash (Macbride 1930). Additionally, field and herbarium studies have yielded material similar to, but differing from “typical” Ribes cuneifolium from both southern Peru (Ayacucho and Cuczo) and lower elevations of Pasco. Species limits therefore need to be critically reexamined. Similarly, Ribes ovalifolium Jancz. was described from the southernmost part of the Department Ancash (Janczewsky 1905) and was considered as a narrow endemic of Ancash and Lima until recently (Mendoza & Monsalve 2007), but recent field studies yielded more material of the species, further expanding its range. The present study aims at providing a critical examination of species limits and updated distribution data plus a detailed morphological characterization of the species involved.
Material and methods
Specimens of Ribes cuneifolium group from BONN, USM as well as the specimens available online from F, MO, G, MA and OXF were analyzed. Additionally, field studies were carried out in all Andean departments of Peru between 1997 and 2019, adding field observations and additional specimens for the study. For morphological analysis, bud scales, leaves, inflorescences, flowers, bracts, bracteoles and fruits of the specimens were measured. The indument and presence or absence of glands was evaluated in all available specimens. The structures such as glands and trichomes in the leaves, petioles, bark, and the details of the flowers were investigated under the stereoscopic microscope (ZEISS, Germany) in the specimens available at the BONN Herbarium.
The micromorphology of leaf surfaces and flowers was additionally studied in liquid preserved material of Ribes cuneifolium from Weigend 8805 (BONN), Weigend 8820 (BONN), and Weigend et al. 5043 (BONN) under the scanning electron microscope (SEM, CAMBRIDGE INSTRUMENTS, England). Material was dehydrated in a graded alcohol series and then critical point dried in a CPD 020 (BALZERS UNION, Switzerland). For viewing the samples were sputter coated with an ultra-thin coating of gold (Au) for 45-60 seconds in the sputtering device SCD 040 (BALZERS UNION, Liechtenstein).
All specimens seen of the Ribes cuneifolium group were mapped by georeferencing localities, unless coordinates were given on the vouchers. Collection localities for vouchers without coordinates were identified with the help of GeoNames respectively Google Earth Pro. Each specimen analyzed was plotted using the package “maps” (Brownrigg 2017) for the R software (R Core team, 2018).
The present study uses a morphological species concept as previous studies on South American Ribes (Weigend & Rodriguez 2005; Weigend et al. 2005, 2010). Species are recognized as distinct if at least two discrete morphological characters differentiate groups of specimens or populations. Infraspecific taxa are recognized where one or several populations differ consistently in individual character states or quantitative characters only.
Results
The results of the morphological investigation show some taxonomic informative characters for the Ribes cuneifolium group, mainly in leaf morphology, and trichome distribution and morphology. We found the Ribes cuneifolium group to be highly conserved in flower morphology, as previously demonstrated for Bolivian species of the genus (compare Weigend & Binder, 2001, Figs. 9, 10, 11). Both Ribes cuneifolium and Ribes ovalifolium have superficially similar leaves, atypically small for Ribes and with poorly developed - if any - lateral lobes, but the two species are easy to distinguish. Ribes ovalifolium has petioles up to 10 mm long with only long stalked trichome glands (Fig. 13), whereas petioles in R. cuneifolium are absent or very short (rarely up to 6 mm) with up to three different types of glands (sessile, minute stalked, long stalked). The lamina of R. ovalifolium (up to 26 × 23 mm) is larger than in R. cuneifolium (up to 15 × 12 mm). Moreover, R. ovalifolium has leaves with a central lobe slightly longer than the lateral lobes, distinctly serrate margins all around, and the leaf base rounded to truncate. Leaves of R. cuneifolium are obscurely three-lobed, with lateral margins entire, and the leaf base cuneate. Finally, adaxial surfaces of R. ovalifolium are eglandular and densely pubescent in contrast to R. cuneifolium with glabrous and glandular (sessile glands) adaxial surfaces. The abaxial surface of R. ovalifolium leaves is pubescent and eglandular to sparsely glandular (with a few sessile glands) while it is glabrous to subglabrous and glandular (sessile glands) in R. cuneifolium.
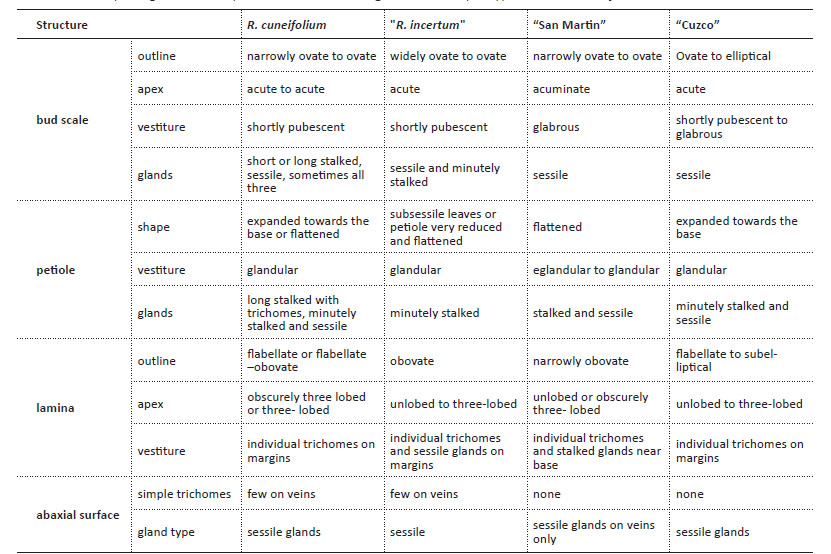
The leaves of R. cuneifolium are typically obovate with the lateral lobes - if present - in the distal half of the lamina (Figs. 4-5). The microphyllous leaves are 3-15(- 20) mm long and 3-9(-12) mm wide and range from subsessile to shortly petiolate, the lamina is flabellate, obovate, or sub-elliptical (Fig. 4). Bud scales are narrowly to widely ovate, and the apex narrowly acute or acuminate (Fig. 1). There is some degree of variability between different populations of R. cuneifolium with regards to leaf shape and indument (Fig. 4), but also flower color (Fig. 8). This continuous range is also evident in the presence of different types of glands on bud scales and petioles. Typically, sessile, minutely stalked, and distinctly stalked glands are present (Fig. 2-3), all three types or only one or two of them may be present in any given individual, with no indication of a clear geographical pattern or correlation with, e.g., leaf morphology. Some specimens differ marginally from the typical condition, e.g., those from San Martín (Tab. 1; Fig. 1E, 2D), Cuzco (Tab. 1; Fig. 1F, 2E), and what has been described as “Ribes incertum” (Tab. 1; Fig. 1C, D, 2C). Despite obvious differences in details of leaf morphology or trichome types, we failed to identify any clear dividing lines between groups of populations. Macbride (1930) described the segregate species Ribes incertum J.F. Macbr. from the Cordillera Blanca in Ancash. Our copious collections from the Cordillera Blanca showed no consistent morphological differences whatsoever to material from Junin (where the type of Ribes cuneifolium comes from), apart possibly from the prevalence of yellow versus greenish-red flowers. The southern collections of R. cuneifolium tend to have slightly longer, more elliptical leaves (Fig. 4 H), but there is no clear distinction to other populations of the species. We color-coded the - poorly differentiated - collections in Fig. 7: All over, there is neither morphological nor ecogeographical evidence (Fig. 7) to support the recognition of separate taxa in R. cuneifolium or maintaining the segregate R. incertum.The only clear exception are the low-elevation collections from Pasco, with considerably larger, flabellate leaves (Fig. 12) and larger inflorescences (see below).
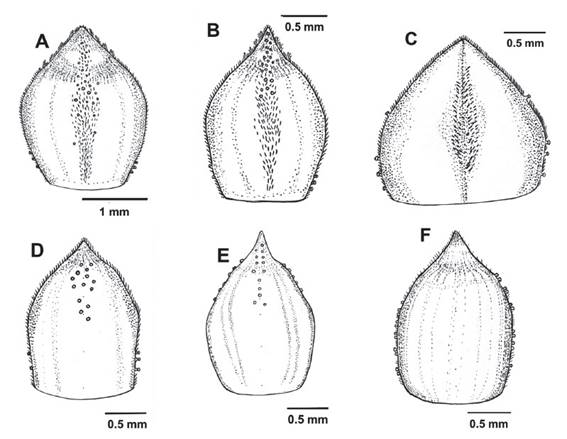
Figure 1 Morphological variation of Ribes cuneifolium bud scales. A-B. Bud scales with all the different glands (stalked with trichomes, minutely stalked and sessile) of Ribes cuneifolium (A Weigend et al. 7676; B Weigend & Schwarzer 8035). C-D. Bud scales of “Ribes incertum” with two types of glands (minutely stalked and sessile) (C Cano et al. 13715, D Cano et al. 13652). E. Bud scale of the specimens from San Martín with only one type of gland (sessile) (Quipuscoa & Vilchez 2614). F. Bud scale with only one type of gland (Cusco, Galiano et al. 6020).
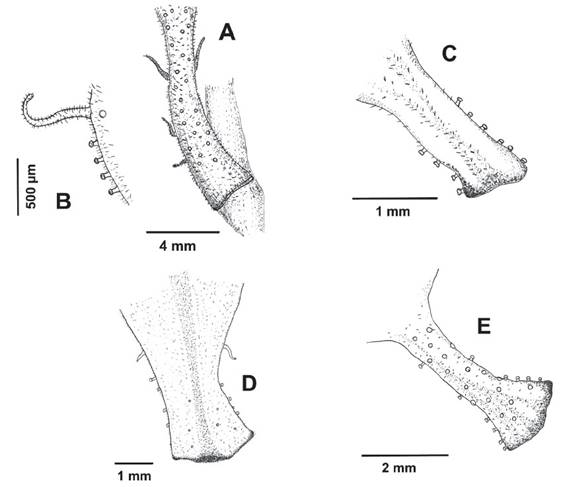
Figure 2 Morphological variation in the petiole of Ribes cuneifolium. A-B. Petiole displaying all the different types of glands (long stalked with trichomes, minutely stalked and sessile) (A Weigend et al. 7676, B Weigend & Schwarzer 8035). C. Reduced flattened petiole of “Ribes incertum” with one type gland (Cano et al. 13715, 13652). D. San Martín specimens with reduced petiole expanded towards the base and two type of glands (Quipuscoa & Vilchez 2614). E. Petioles with two type of glands (minute stalked and sessile glands, Cusco, Galiano et al. 6020).
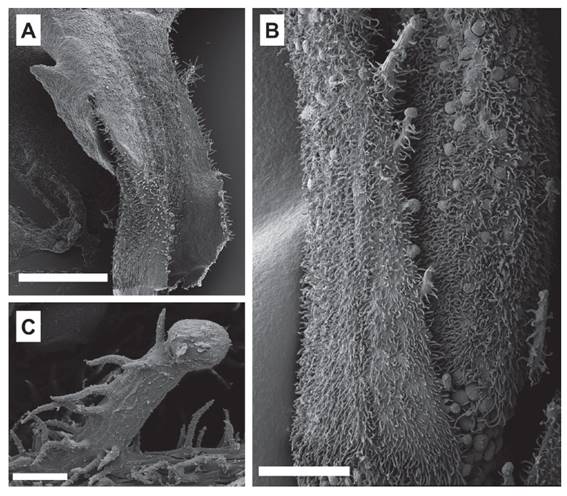
Figure 3 Different types of glands of Ribes cuneifolium petiole. A. Free petiole with long stalked with trichomes gland and minutely stalked glands (Weigend 8820). B. Petiole attached to the branchlets with all types of glands known from Ribes cuneifolium: long stalked glandular trichomes, minutely stalked glandular trichomes and sessile glands. C. Long stalked glandular trichomes on Ribes cuneifolium petiole (Weigend et al. 5043). Scale bars A, B: 500 µm; C: 50 µm.
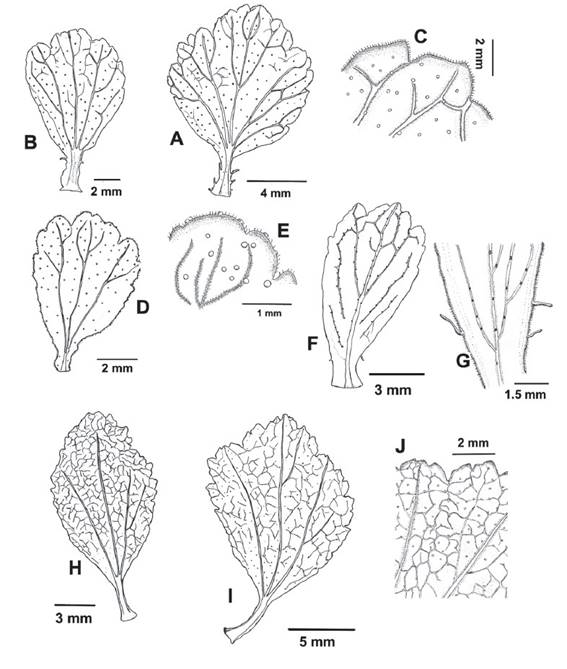
Figure 4 Morphological variation of Ribes cuneifolium leaves. A-B. Flabellate and obovate leaves both with cuneate leaf base (A Weigend et al. 7676, B Weigend & Schwarzer 8035). C. Subglabrous abaxial surface with sessile glands (Weigend et al. 7676). D. Obovate leaf observed in the formerly “R. incertum” with cuneate leaf base (Cano et al. 13715). E. Subglabrous abaxial surface with sessile glands which are also present in the margin (Cano et al. 13715, 13652). F-G. Specimens from San Martin with cuneate leaf bases and abaxial surfaces with sessile glands restricted to the veins (Quipuscoa & Vilchez 2614). H-I. Flabellate and subelliptical leaves with cuneate leaf bases, from Cusco. J. Abaxial surface with sessile glands (Cusco, Galiano et al. 6020).
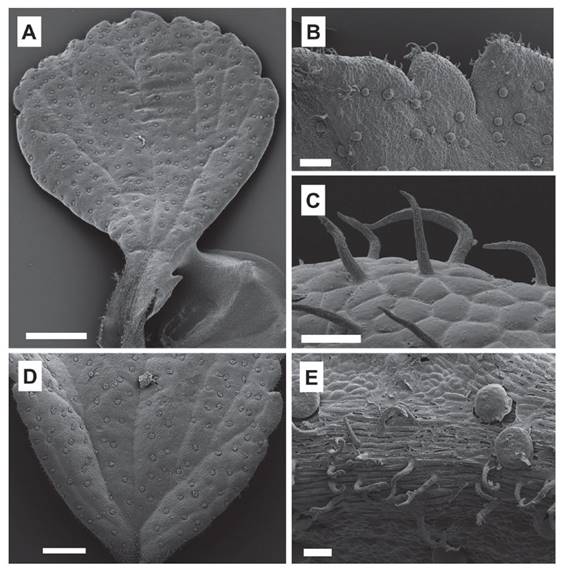
Figure 5. Microstructures of the leaf surfaces of Ribes cuneifolium. A. Adaxial surface with sessile glands (Weigend 8805). B-C. Leaf margin with unicellular trichomes. D. Abaxial surface with many sessile glands (Weigend et al. 5043). E. Abaxial surface with the presence of unicellular trichomes mainly on the vein (Weigend et al. 5043). Scale bars A, D: 700 µm, B, C: 200 µm, E: 50 µm.
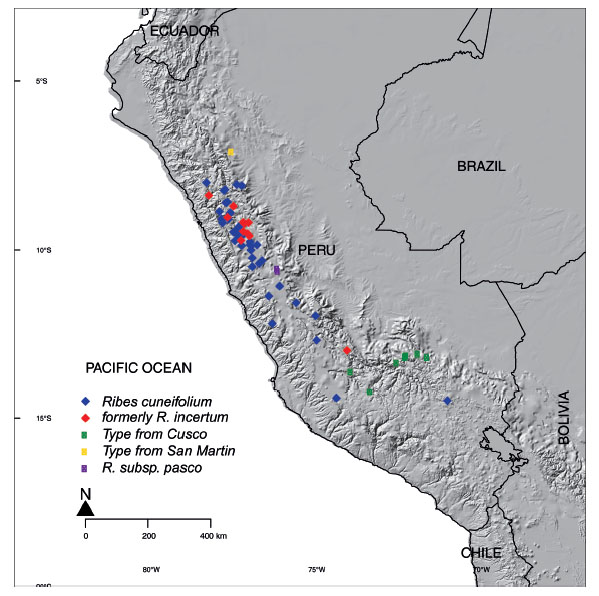
Figure 7 Distribution map of Ribes cuneifolium. Morphotypes and the new subspecies are color-coded, based on herbarium specimens analyzed.

Figure 8 Inflorescences and flowers of Ribes cuneifolium A-B. Flowering branch with and individual inflorescences with pale yellow flower, west side of Abra Carpish, Dep. Ancash (Weigend et al. 8891). C-D. Typical red-flowering form, note the five large sepals and five tiny petals, Huallanca, Prov. Bolognesi, Dep. Ancash (Weigend 8805).
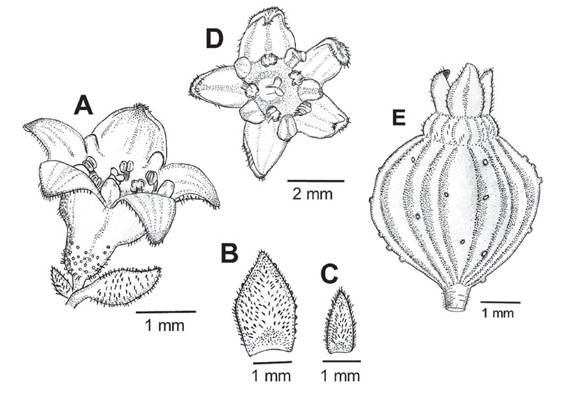
Figure 9 Flower and fruit of Ribes cuneifolium. A. Flower with sessile glands toward the ovarian portion. B. Bract with sessile glands restricted to the margin. C. Bracteole. D. Pentamerous flower. E. Fruit with few sessile glands. (Huallanca, Prov. Bolognesi, Dep. Ancash (Weigend 8805).

Figure 10 Details of the inflorescence of Ribes cuneifolium, SEM. A. Inflorescence displaying one flower as well as bracts and bracteoles. B. Pentamerous flower of Ribes cuneifolium with its petals and stamens inserted ca.1 ml from the base of the hypanthium. C. Glandular hypanthium (sessile glands). D. Ovate bract pubescent from unicellular trichomes and glandular with sessile glands mainly on the margin. E. Rachis with ovate bracts and lance-ovate bracteoles (Weigend 8805). Scale bars A, E: 700 µm; B, C: 500 µm; D: 300 µm.
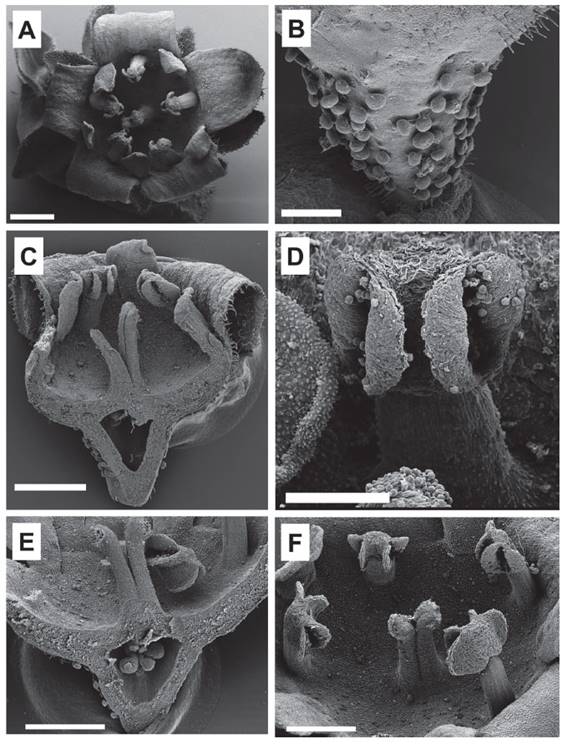
Figure 11 Details of the flower of Ribes cuneifolium. A. Pentamerous flower (Weigend 8805). B. Many sessile glands present in the hypanthium of R. cuneifolium (Weigend 8820). C. Longitudinal section of Ribes cuneifolium flower showing the inferior ovary. D. Stamen. E. Longitudinal section of Ribes cuneifolium showing the presence of ovules. F. Deeply bifid style surrounded by stamens with basifixed anthers (Weigend 8805). Scale bars A: 700 µm; B: 300 µm; C, E, F: 500 µm; D: 200 µm.
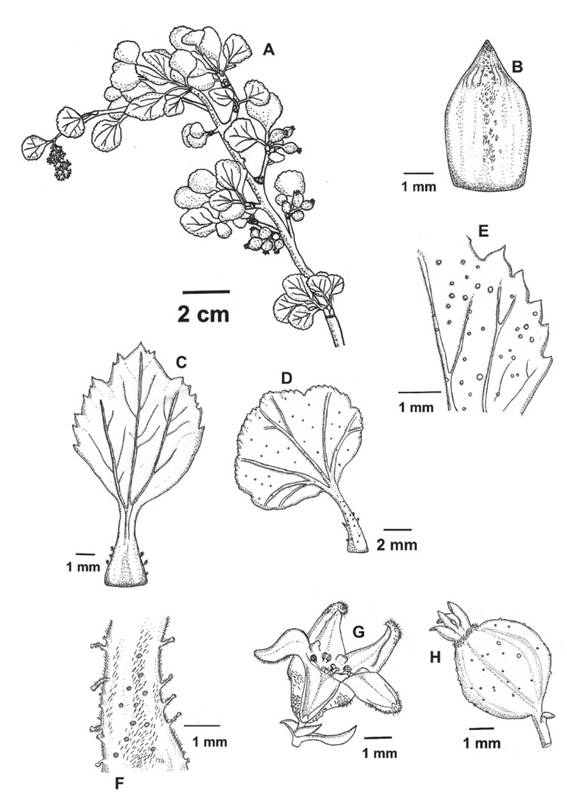
Figure 12 Ribes cuneifolium subsp. pascoense. A. Flowering branch. B. Bud scale with few and scattered minutely stalked glands. C. Ovate leaf D. Flabellate leaf. E. glabrous abaxial surface with sessile glands. F. Petiole with three type of glands (sessile, minutely stalked and stalked with trichomes). G. Pentamerous flower. H. Fruit. (Type specimen Rivas et al. s.n.)
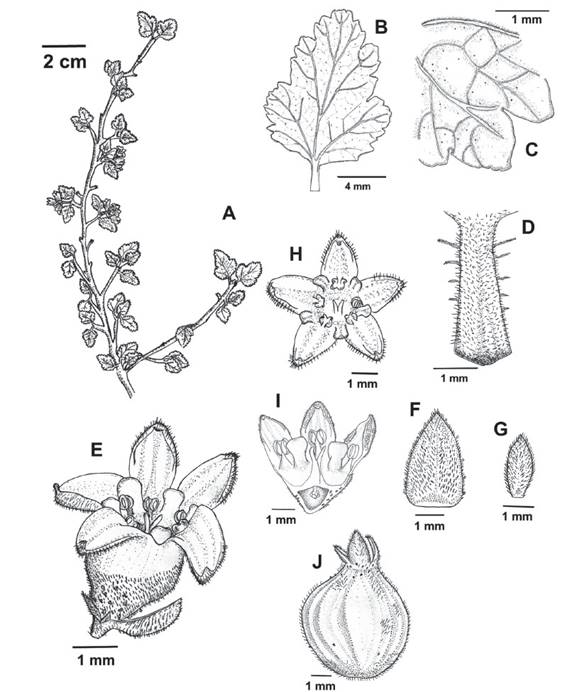
Figure 13 Ribes ovalifolium. A. Flowering branch B. leaf. C. Abaxial surface with simple individual hairs mainly located on veins, and sessile glands. D. Petiole with long stalked glandular trichomes. E. Flower with minutely stalked glands towards the ovarian portion. F. Bract with minutely stalked glands. G. Bracteole H. Pentamerous flower I. Flower longitudinal section displaying the oblong petals and the ovary fused with the hypanthium. J. Fruit with few minutely stalked glands (Weigend et al. 7756).
Key to the species of the Ribes cuneifolium group:
1. Leaves obovate, elliptical or subcircular, unlobed and only deeply, irregularly serrate and sometimes obscurely lobed in the distal part (1/3rd or 2/3rds) of the lamina; leaf margin serrate only distally, lamina densely covered with sessile glands, sometimes with very few scattered simple trichomes. 1. Ribes cuneifolium
1´. Leaves widely ovate, unlobed or with one lateral lobe on each side in the proximal half of the lamina; leaf margin serrate along the entire margin, lamina densely pubescent adaxially and abaxially and with scattered sessile glands. 2. Ribes ovalifolium
Taxonomic treatment
Ribes cuneifolium Ruiz & Pav.
Flora Peruviana, et Chilensis III (1802:13, tab. 233 c).
Shrub 0.3-4 m high, very densely branched with numerous short shoots; branches narrowly spaced, lateral branches typically 3-5 mm long. Bark matt brown to gray, not or weakly exfoliating later; on young branches shortly pubescent with simple individual hairs, with minutely stalked or subsessile glands. Bud scales ovate, apex acute, rarely acuminate, 1-5 × 1-3 mm; abaxially finely pubescent with a few simple hairs, minutely stalked glands on margin, sometimes also with sessile glands, rarely also with subsessile glands. Leaves evergreen or semi-deciduous, coriaceous; subsessile to shortly petiolate, petiole 1-3.5(-7) mm long, flattened and expanded especially towards the base, shortly pubescent with unicellular hairs and, sessile, minutely stalked and/or stalked glands. Lamina flabellate, obovate or sub-elliptical, 3-15(-20) × 3-9(-13) mm; three-veined; entire, obscurely or distinctly three-lobed, incision between lobes less than ¼ of leaf length, free portion of the central lobe up to 3 × 6 mm; lateral margin entire, distal margin serrate with (2-)3-6(-10) serrations per lobe; margin with few small unicellular trichomes and in some cases sessile or stalked glands restricted to the base of the leaf margin; leaf base cuneate; adaxial surface glabrous with sessile glands; abaxial surface glabrous to subglabrous with very few simple trichomes and sessile glands. Inflorescence terminal racemes on brachyblasts, usually deflexed, (5-)8-13(-26) mm long; with 3-14 flowers; peduncle 1.5-5 mm long, variously pubescent with simple unicellular trichomes, eglandular or with few sessile glands; pedicels 0-2 mm long, ca. 1 mm apart in open flowers, pubescent from simple trichomes; bracts ovate-acuminate, 1-3 mm long, pubescent from simple small trichomes and with scattered sessile glands; bracteoles narrowly ovate-acuminate, ca. 1 mm long, with scattered unicellular trichomes and some sessile glands. Flowers rotate, 2-5 mm long, 2.5-5 wide, pale yellow, cream-coloured, green or dark red; hypanthium 1-2 mm long, largely glabrous, externally pilose especially at base and with many sessile to subsessile glands on the inferior ovary; calyx lobes ovate-acuminate, 1-2,5 × 1-1.5 mm, (sub-)glabrous, eglandular; petals narrowly oblong, apex rounded to truncate, margin entire, ca. 0.6-0.8 × 0.3-0.5 mm, glabrous; five stamens, filaments ca. 0.6-0.9 mm long, anthers basifixed, ca. 0.2-0.4 mm, nectarial glands absent; style deeply bifid; ovary inferior, fused with hypanthium, bicarpellate, unilocular, with several anatropous ovules. Fruit a spherical berry, 1-5 per raceme, pale red or green, ca. 5-8 mm in diameter, glabrous, with a few sessile glands; perianth remnants ca. 1-2 mm.
Notes: Ribes cuneifolium is a very widespread and often common high-elevation species. We propose the recognition of two subspecies - the very common and widespread typical subspecies cuneifolium and the newly described subspecies pascoense.
Key to the subspecies of Ribes cuneifolium:
1. Lamina flabellate to elliptical, 5-15(-20) × 3-9(-12) mm, base cuneate; inflorescence with 3-8 flowers; peduncle 1.5-4.5 mm long.
1.1 Ribes cuneifolium subsp. cuneifolium
1´. Lamina (flabellate to) subcircular, 6-16 × 7-13 mm, base cuneate to obtuse; inflorescence with 8-14 flowers; peduncle 4-5 mm long.
1.2. Ribes ovalifolium subsp. pascoense
1. Ribes cuneifolium Ruiz & Pav. subsp. cuneifolium
Type: PERU. [Junin, Diezmo], s.d., H. Ruiz & J. Pavón, s.n. (holotype MA MA811943!; paratypes : MA MA811942 [photo!], G barcode G00388802 [photo!], F V0066659F [frag. MA811942], F! Neg. No. 27343, OXF 00059252[photo!]). = Ribes incertum J.F. Macbr., Publ. Field Mus. Nat. Hist., Bot. Ser. 8: 116 (1930: 116-117) Type: PERU. [Ancash], Pomopamba, Estación 30 miles east of Huaraz, 4 October 1922, F. Macbride & [W.] Featherstone 2493 (holotype F V0066662F [photo!] F!/Neg. No. 64979; isotypes: G: G00388804 [photo!], G: G00388803[frag.!]).
Bark on young branches shortly pubescent with simple individual hairs, and glandular with sessile, minute stalked (rarely present), or subsessile glands. Bud scales ovate, apex acuminate, 1-3 mm long, 1-2.5 mm wide. Petiole 1-3.5(-6) mm long. Lamina flabellate, obovate, oblance-ovate or elliptical, 5-15(-20) mm long, 3-9(-12) mm wide; three veined; unlobed, obscurely or distinctly three-lobed, incision between lobes less than ¼ of leaf length, free portion of the central lobe up to 3 × 6 mm; base cuneate; abaxial surface glabrous to subglabrous with very few simple individual small trichomes (especially on veins) and glandular with sessile glands sometimes restricted to the veins. Inflorescence (5-)8-13(-26) mm long in flower; with 3-8 flowers; peduncle 1.5-4.5 mm long, pedicels 0-1-2 mm long. Flowers 2-3.5 mm long, 2.5-5 wide, hypanthium 1-1.5 mm long.
Distribution and Habitat: This subspecies is widely distributed in Peru. There are collections of this species from Ancash, Apurimac, Ayacucho, Cuzco, Huancavelica, Huánuco, Junín, La Libertad, Lima, Pasco, and San Martín. It is mainly found in isolated patches at the base of rocks and in small valleys in páramo and puna vegetation. Thespecies is found at elevations from 2900 m to 5100 m. At the upper limit of its distribution it is often the only sizeable woody plant and associated with superparamo vegetation (Lysipomia, Phyllactis, Chuquiragua) and reaches sizes of barely 50 cm. At lower elevations it may be the dominant species of dwarf forests ca. 4 m tall or it may be associated with rock-falls and tall stands of Polylepis spp. Where heavily browsed, it may form a dense, cushion-shaped topiary, especially in drier formations (Fig. 6).

Figure 6 Habit and Habitat of Ribes cuneifolium Ruiz & Pav. A-B. Tall shrubs in Polylepis thicket, Huallanca, Dep. Ancash (Weigend 8805). C-D. Isolated patches at the base of the rocks in Páramo, Pamparomas, Dep Dep.. Ancash (Weigend s.n.). E-F. Free standing shrubs in Puna vegetation, Prov. Huaylas, Dept. Ancash (Weigend, s.n.).
Phenology: Flowering appears to be concentrated in May and June, but there are also flowering specimens from January, February, March, August, October, and November, indicating that the species flowers more or less opportunistically throughout the year. Unlike all other species of Ribes, this species may also pass into flower a second time with individual plants bearing flowers and fruits at the same time.
Notes: Ribes cuneifolium subsp. cuneifolium as here redefined ranges from San Martín in the North to Cuzco in the South. Across the geographical range there is some degree of clinal variation in morphology which can be roughly grouped into four variants (compare Table 1). These include “Ribes incertum”, originally differentiated from R. cuneifolium based on putatively elliptical leaves and yellow flowers, two characters that turn up independently from each other in different populations of the species across most of its range (see Fig. 14). The northern populations and most of the southern collections (esp. from Cuzco) are marginally different in habit and leaf shape, but there is no clear dividing line nor complete ecogeographical segregation from typical R. cuneifolium. We therefore advocate defining the subspecies broadly.
Table 1 Morphological variability across the different regional and morphotypes of Ribes cuneifolium.
Additional specimens examined: PERU. Ancash: Villcamanín, near Huaraz, 3640 m, 24 March 1983, O. Tovar et al. 9931 (USM); Prov. Antonio Raimondi, Distict of Chingas, Yanachancha Lagoon and surroundings, 4200-4300 m, 9° 10' 50'' S, 77° 4' 31, 1'' W, 22 - VIII - 2003 , A. Cano et al. 13715 (BSB, USM); Prov. Bolognesi, road from Abra Janashalla down to Huallanca, bellow Huansalá, 3390 m, 9° 52,083' S, 76° 59,443' 08 W, X - 2007, M. Weigend et al. 8805 (BONN); Road from Huallanca to San Marcos via Lago Canrash, 4230 m, 09 - X - 2007, M. Weigend et al. 8820 (BONN); 3900 m, VIII - 1972, A. Cerrate 4 (USM); Jahuacocha, surroundings of the lagoon sustained by the Jerupaja, 5100 m, 27 - V - 1954, E. Cerrate 2274 (USM); 3800 m, 24 - X - 1981, E. Cerrate et al. 8358 (USM); Prov. Corongo, road Sihuas to Corongo/Yuramarca, on the north-west-facing slopes above Río Huallcallanca before reaching Mamapamba, 3912 m, 8° 34,193' '' S, 77° 42,741' '' W, 26 - VI - 04, M. Weigend & Ch. Schwarzer 8035 (USM, HUSA, HUT, BSB, BONN); road from Yanac to Sihuas. Near Mamapamba, ca. 3500 m, 08 - VIII - 2001, M. Weigend et al. 5043 (HUT, M, USM, BONN); Prov. Huaraz, Nevado Huascarán, below Glaciar Kinzl, 4400 m, 17 September 04, M. Richter s.n. (BONN); Alpamayo valley, 4250 m, 12 - VIII - 2004, M. Richter s.n. (BONN); Prov. Huari, San Marcos, Yanacancha, above the Antamina mining camp, 9° 34' 34,7'' S, 77° 2' 57,6'' W, 06 - VII - 2003, A. Cano et al. 13652 (BSB, USM, BONN); Road from San Luis to Huari, after pass, 4000 m, 13 - III - 2001, M. Weigend et al. 5139 (BSB, M, USM, BONN); Chavin de Huantar, road from Chauvin de Huantar to Catac via Abra Carpish, 4180 m, 10 - X - 2007, M. Weigend et al. 8891; (Prov. Huaylas?) PN Huascarán, Parón valley, 3760 m, 9° 01'S, 77° 43' W, 1 - III - 1985, Smith & Goodwin 8932 (F: V0280546F!, USM!, MO); Prov. Huaylas, 47 km after Caraz on road to Casma, pass height, 4391 m, 14 - III - 2003, M. Weigend et al. 7676 (HUT, B, USM, M, BONN); Ocshapamba (Oxapamba), 4350-4630 m, 9° 11' 29,7'' S, 77° 51' 50,7'' W, 11 - III - 1999, A. Cano 9743 (USM); Prov. Pallasca, Pampas, Cordillera Pelagatos, 4500-4794 m, 8° 12' 3.636" S, 77° 47' 3.9114" W, 20 - III - 2011, A. Cano et al. 20284 (USM); District of Conchucos, midle part of the ravine Toldobamba, 4154 m, 8° 13' 35.148" S, 77° 49' 12.2154" W, 15 - IX - 2015, P. Gonzáles & N. Tenorio 3842 (USM); Prov. Recuay, road Catac to Chavin de Huantar via Laguna Querococha; between Laguna Querococha and Abra Cahuish/ (FO 4), 4170 m, 17 - V -2003, M. Weigend et al. 7729 (HUT, B, USM, M, BONN); Road to Aija, 3890 m, 9° 43' 18.9'' S, 77° 29' 19.2'' W, 29 - V -2001, B. León et al. 4931 (USM); Prov. San Luis, road from San Luis to Huari before Laguna Huachococha and near Laguna Huachococha, 4000-4500 m, 13 - III - 2001, M. Weigend et al. 5116 (BSB, M, USM, BONN); Prov. Sihuas, Road from Sihuas to Pomabamba, pass after Sicsibamba, before Palo Seco, 3700-3900 m, 11 - III - 2001, M. Weigend et al. 5095 (BSB,M, USM, BONN); Ayacucho: Pedregal (Approximately 2 km of road to Minas Canarias), 4150-4200 m, 14° 24' 30.4'' S, 74° 24' 23.5''23 W, II - 2002, A. Cano et al. 11896 (USM); Prov. Huamanga, Quinua, in the highlands of Pallcca, 4000 m, V - 2014, J. Hurtado JAHH-135 (USM); Quinua, in the highlands of Pallcca, 4000 m, V - 2014, J. Hurtado JAHH-134 (USM); Prov. Vilcashuaman, Intiwatana-Pizará, 3500 m, 09 - I - 2007, J. Barrientos 34 (USM); Cusco: Prov. Calca, District Calca, rise to the Peñas, 2900 m, 13° 11' 58'' S, 72° 17' 50'' W, IV - 2004, W. Galiano et al. 6020 (MO); Roads from Calca to Amparaes, after pass above village of Aymaraes, 3600-3800 m, 6 - II - 2000, M. & K. Weigend 2000/192 (fr., F, HUSA, NY, USM, M); Prov. Calca (?): Above Sañaica, 3800 m, XI - 1915, Weberbauer 7176 (F!, HUH); Prov. Calca, Salcantay (?), 4400 m, 29 - V - 1954, Rauh-Hirsch P1428 (F). Prov. Urubamba, road from Cusco to Quillabamba, Abra Málaga, 3600-3700 m, 25 - II - 2000, M. & K. Weigend 2000/463 (F, HUSA, NY, USM!, M); Road from Cuzco to Quillabamba, before Abra Málaga, 3400 m, 23 - II - 2000, M. & K. Weigend 2000/406 (fr., F, HUSA, NY, USM!, M); Prov. Paucartambo, road from Pucartambo to turnoff Tres Cruces, 4000 m, 9 - II - 2000, M. & K. Weigend 2000/227 (st., F, HUSA, NY, USM!, M); unknown locality: Piñasniocj, Panticalla pass, 3600 m, 16 - VII - 15, Cook & Gilbert 1872 (HUH). Huancavelica: Tansiri, Conaica, 4500 m, 29 March 1967, O. Tovar 5858 (USM); Prov. Castrovirreyna, Choclococha, 4500-4600 m, V - 1958, O. Tovar 2844 (USM); Prov. Tayacaja, District of Huando, Tinllacaya, 4683 m, 12° 40' 58.1514" S, 75° 0' 25.704" W, 29 - IV - 2007, H. Beltrán 6357 (USM); Huánuco: Prov. Dos de Mayo, near Huallanca, 3700 m, 23 - III - 1983, O. Tovar et al. 9878 (USM); Prov. Lauricocha, District of Cauri, sorroundings of the lagoons Tinkicocha, Lauricocha, Tauricocha y Patarcocha, 4000-4200 m, 10° 24' 4.3194" S, 76° 45' 8.8194" W, 08 - IX - 2012, P. Gonzáles & N. Valencia 1916 (USM); Cordillera de Raura. Between Tinquicocha and Patarcocha lagoons, 4200-4500 m, 24 - VI - 2000, A. Cano & N. Valencia 10721 (USM); District of San Miguel de Cauri, Lauricocha lagoon, south west side, 3845 m, 13° 10'S, 72° 17' W, 08 - VII - 2002, F. Salvador et al. 410 (USM); Junin: Prov. Huancayo, Ascent to the snow Huaytapallana, 4200 m, 19 - IX - 1987, O. Tovar, et al. s.n. (USM); District of Huancayo, Acopalca between the abra Huaytapallana and the Carhuacocha lagoon, 4491-4521 m, 11° 57' 21.4554" S, 75° 2' 30.264" W, 10 - III - 2011, D. Rodríguez 537 (USM); Hauaytapallana snowy, east of Huancayo, 4200-4600 m, 7 - XI - 1984, O. Tovar 9991 (USM); Prov. Junin, Ondores, 4100 m , 29 - IX - 1976, U. Pettersson 353 (USM); Prov. Tarma, 20 km SE of Tarma on road to Jauja, 4100 m, 28 - XI - 1962, H. Iltis 166 (USM!, UC, WISC); La Libertad: Prov. Pataz, road (Humachuco to) ChagualBuldibuyo-Tayabamba, between Parcoy and Buldibuyo, 3739 m, 8° 2,969'22" S, 77° 26,174' W, 22 - IV - 2004, M. Weigend & Ch. Schwarzer 7937 (B, USM, HUT, BONN); Road (path) Buldibuyo to Zona Maderas, pass height towards Punta Maderas, 3775 m, 8° 5,106'23" S, 23 - IV - 2004, M. Weigend & Ch. Schwarzer 8005 (B,USM, HUSA, HUT, BONN); Road (Huamachuco to) Chagual-Buldibuyo-Tayabamba, between Chagual and Parcoy, slopes in side-valley of Río San Miguel, 5km S of Parcoy, 22 - IV - 2004, M. Weigend & Ch. Schwarzer 7934 (USM!, B, HUSA, HUT); Prov. Santiago de Chuco, Santuario Nacional Calipuy, 3686-4361 m, 8° 22' 21.0714" S, 78° 16' 50.124" W, VI - 2012, M. Morales et al. 3676 (USM); Lima: Prov. Cajatambo, Raura, 4600 m, 15 - VI - 1988, S. Rivas et al. s.n. (USM); Road from Cajatambo to Ocros, pass height after Rajan towards Ocros, 4145 m, 10,37185° S, 77,31637° W, 04 - III - 2018, M. Weigend & K. Peñas 9802 (USM); Prov. Chuncal, 4050 m, 11° 22' 10'' S, 76° 27' 36'' W, 25 - III - 2005, C. Aedo & A. Galán de Mera 10856 (USM!, MA); Prov. Huarochiri, Cerro Condorcoto, 4030 m, 8 - III - 1968, E. Cerrate et al. 4628 (USM); San Martín: Prov. Huallaga, District Saposoa, Jalca de Rangrapata, between rocks, 3420 m, 23 - VIII - 2001, V. Quipuscoa & M. Vilchez 2614 (BSB, HUT, USM, M).
1.1. Ribes cuneifolium Ruiz & Pav. (1802:13, tab. 233 c) subsp. pascoense Weigend, subsp. nov. Fig. 12
Type: PERU. Pasco, Prov. Pasco, La Quinua, 3540 m, 01 - XII - 1986, S. Rivas, O. Tovar, J. Loidi, P. Canto, s.n. (holotype USM, USM 295711).
Ribes cuneifolium subsp. pascoense differs from subsp. cuneifolium in widely flabellate to subcircular leaves (vs. obovate to elliptical) and inflorescences 12-17 mm long with 8-14 flowers (vs. 8-13 mm with 3-8 flowers).
Bark brown and with sessile glands only. Bud scales ovate-acuminate, 3-5 × 2-3 mm. Petiole 3-7 mm long. Lamina (flabellate to) subcircular, 6-16 × 7-13 mm; three veined; entire or very obscurely three-lobed, incision between lobes less than 1/8 of leaf length, free portion of the central lobe up to 1.5-5 × 2-3.5 mm; base cuneate to obtuse; abaxial surface glabrous and glandular with sessile glands. Inflorescence 12-17 mm long in flower; with 8-14 flowers; peduncle 4-5 mm long, pedicels up to1 mm. Flowers subsessile, 4-5 mm long, 3-4 wide; hypanthium 1.5-2 mm long.
Distribution: This subspecies is only known from Dept. Pasco, Prov. Pasco. It was found at elevations of 3540-3780 m in cloud-forest remnants.
Phenology: The type specimen was collected flowering in December.
Etymology: The epithet “pascoense” is derived from Pasco, the name of the Peruvian department where the type collection came from.
Notes: Ribes cuneifolium subsp. pascoense is only known from a handful of collections from a small area in Pasco and may well be more widespread on the eastern Andean slope in Junin, Pasco and San Martín. Additional collections would be clearly desirable to improve our understanding of this species.
Additional specimens examined: PERU. Pasco: Prov. Pasco, Cerro de Pasco-Huariaca, 3780 m, 8 - II - 1984, S. Rivas, O. Tovar & P. Canto s.n. (USM), Prov. Pasco, Cerro de Pasco, road from Huánuco to Cerro de Pasco, km 79, 3740 m, 10° 38' S, 76° 10' W, 03 - IV - 2001, M. Weigend et al. 5468 (HUT, BSB, M, USM, BONN).
2. Ribes ovalifolium Jancz.
Bull. Acad. Cracovie 1905, 761; et in Mem. Soc. Phys. Hist. Nat. Geneve xxxv. 426 (1907: 761).
Type: PERU. [Ancash], 5 August 1903, A. Weberbauer 2771 (B† F!: Neg. No. 4159).
Shrub 0.9-2 m tall; bark matt brown to bright gray, not or weakly exfoliating, moderately pubescent with simple trichomes, eglandular. Bud scales ovate-acuminate, 1.5-3 × 1.5-2.5 mm, densely pubescent with simple trichomes, eglandular or glandular with minutely stalked and stalked glands esp. along the margins. Leaves deciduous; petiole 2 -5-(10) mm, densely pubescent, eglandular to glandular with long stalked glands which along the margins. Lamina ovate to subelliptical, 7 -19-( 26) × 5 -15-(23) mm; three-veined; obscurely to distinctly three-lobed, central lobe slightly longer (1.5 ×) than the two lateral lobes, incisions between lobes ¼ of leaf length; free portion of central lobe 3-11 × 4-9 mm; leaf margins all serrate, distal margins with 3-7(-13) serrations per lobe; leaf base rounded to truncate; adaxial and abaxial surface densely pubescent with simple trichomes, adaxially eglandular, abaxially eglandular or with numerous sessile glands. Inflorescence terminal racemes on brachyblasts, pendulous, 8 mm long in flower; with 2-3 flowers; peduncle 2.5-3 mm long, pubescent with simple unicellular hairs; bracts ovate-acuminate, 2.5-3 × 1.5-2 mm, pubescent, glandular with minute stalked glands along margins; bracteoles lanceolate, apex narrowly acute, 1-1.5 × 1 mm, pubescent with unicellular trichomes. Flowers subsessile, rotate, 5.5-6 × 4-5 mm, reddish brown; hypanthium 2.5-3 mm, densely pubescent with simple unicellular hairs, glandular with minutely stalked glands towards the ovary; calyx lobes ovate, apex acute, 1.5-2.5 × 1-2 mm, abaxial pubescent with simple unicellular hairs; petals oblong, apex rounded, entire, 1 × 0.5-1 mm; five stamens, filaments 0.7 mm, anthers basifixed, 0.5 mm; style bifid; ovary inferior, fused with hypanthium, bicarpellate, unilocular, with several anatropous ovules. Fruit spherical berries, ca. 5-7 mm in diameter, sparsely pubescent with simple individual hairs, glandular with minutely stalked glands; perianth remnants ca. 1 mm long.
Distribution and Habitat: The species is present in Ancash, Arequipa, Ayacucho, Lima, Moquegua, Tacna where it is restricted to the arid western slopes of the Andes. It is mainly found in rocky slopes, on rock surfaces and in stabilized scree slopes at elevations of 2400-4000 m.
Phenology: Based on the rare flowering specimens in herbaria, the plant appears to flower in November.
Notes: Ribes ovalifolium is a superficially similar to Ribes cuneifolium, but is easily distinguished by its densely pubescent leaves (vs. glabrous to subglabrous in R. cuneifolium) with a rounded or truncate bases (vs. cuneate), leaf base and serrate leaf margins all around (vs. only distally). The species is heavily browsed by sheep, cattle and presumably both wild and domestic camelids. Therefore, across most of its range, only individual plants are found, typically on inaccessible rockfaces and between boulders. The only larger population with fully developed plants observed in the field by one of the authors was found in northern part of the Cordillera Negra on a rockslide (M. Weigend & Ch. Schwarzer 8065). This is, at the same time, the northernmost population of the species so far reported.
Additional specimens examined: PERU. Ancash: Cordillera Negra, road (Caraz to) Huaylas to Jimbe across the Cordillera Negra, after passing through Huamanyaco, 4095 m, 8° 51,3' S, 77° 57,5' W, 29 - IV - 2004, M. Weigend & Ch. Schwarzer 8065 (USM, HUSA, HUT, BONN); Prov. Bolognesi, Near Acas, 3600 m, 16 June 1979, E. Cerrate 7628 (USM); Road to Cochpun, 3579 m, 19 - VIII - 1981, E. Cerrate 8130 (USM); Cunuc ravine, between Ocros and Chonta, 3600 m, 10 - VII - 1974, E. Cerrate 6245 (USM); Prov. Huaylas, Caraz, 2418 m, 9° 2' 48.1194" S, 77° 48' 41.04" W, 11 - VIII - 2010, Xue-Jun Ge et al. 206 (USM); Ayacucho: Above the Port Toro Muerto, Km 77-78, highway Nazca-Puquio, 3500-3520 m, 14° 41' 23,8'' S, 74° 30' 36,1'' W, 23 - II - 2002, A. Cano et al. 11867 (USM); Arequipa: Surroundings of La Union above Puyca, 3800 m, VI - 2000, T. Hofreiter s.n. (M, USM, BONN); Lima: Prov. Cajatambo, Road from Cajatambo a Ocros, pass height after Rajan towards Ocros, 4145 m, S 10.37185, W 77.31637, 04 - III - 2018, M. Weigend & K. A. Peña Ramos 9802 (BONN, USM, HUT). 10 km NE of Suchi, ca. 61 road km NE of Chosica on road to Huanza, 3900-4000 m, 06 - V - 1978, A. Gentry 21637 (USM); Above Huarochiri, 4000 m, June 1953, E. Cerrate 2052 (USM); Prov. Canta, Lachaqui, Pedregales de Arquircancha, 3650 m, 29 - III - 1991, G. Vilcapoma 831 (USM); Lachaqui, roquedales de Parianhuasi, between rocks, 22 - V - 2004, G. Vilcapoma 7121 (USM); Distict of Huaraz, Huaraz, 3500 m, 11° 24' 48.62'' S, 76° 34' 40.92'' W, 07 - VI - 2008, P. Gonzáles 230 (USM); Prov. Huarochiri, in front of the cave Mortero, District of Mariatana, 3850 m, 14 - IV - 1968, E. Cerrate et al. 4799 (USM); Pacomanta, Km 120 highway Lima-Huarochirí, in the ravine between big stones, 3800 m, 01 - IXII - 1987, S. Baldeón 705 (USM); Caigola, District of Mariatana, 3250 m, 10 - IV - 1968, E. Cerrate et al. 4664 (USM); San Pedro de Casta, citadel of Marcahuasi, 3850 m, 10 - VI - 1993, G. Yarupaitán & J. Albán 731 (USM); Prov. Yauyos, District of Laraos, Peshcohayco, between Peshco and Trespuquios. 4096 m, 12° 21,61' S, 75° 48,93' W, 03 - II - 2000, H. Beltrán 3375 (USM, B, BONN); Cochapunco, above Tupinacocha, 3600 m, 18 - I - 1952, E. Cerrate 1209 (USM); Moquegua: Prov. General Sanchez Cerro, between Arequipa and Pocsi (after highest point in road), dry scrub formation, 3300-3400 m, 16° 35' 23'' S, 71° 25' 52'' W, 21 - V - 2003, M. Weigend et al. 7756 (BSB, USM, BONN); Tacna: Candarave, Yucamani volcano, 3100-3400 m, 09 - XII - 1997, M.I. La Torre 1962 (USM).

