











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Las lagartijas del género Liolaemus es uno de los grupos de vertebrados más diversos con más de 280 especies (Uetz et al. 2022). Este género usualmente se ha dividido en dos subgéneros (Eulaemus y Liolaemus), pero estudios recientes han demostrado que la posición del clado walkeri es basal a ambos (Aguilar et al. 2013, Aguilar et al. 2015, Esquerré et al. 2019) sugiriendo un nuevo subgénero. Las especies del grupo montanus del subgénero Eulaemus se distribuyen en Argentina, Bolivia, Chile y Perú desde el nivel de mar hasta más de 5 mil metros (Olivera & Aguilar 2020). Las especies de este grupo que habitan el territorio peruano se encuentran principalmente en la región costera y andina hasta los 3000 m de altitud, la mayoría de ellas con un fenotipo distintivo (“phrynosauroide”); sin embargo, en la región altoandina están la mayoría de las especies (Aguilar-Puntriano et al. 2018, 2019).
Una de las especies recientemente descritas del grupo montanus es Liolaemus warjantay Ubalde-Mamani et al. 2021, que habita las partes altas de los Andes a 4519 m de altitud. Esta especie fue descrita en base a muestras procedentes del departamento de Arequipa (sur de Perú), y todas ellas cerca de la localidad tipo (Ubalde-Mamani et al. 2021). A pesar de tener secuencias de cyt-b de trabajos previos, Ubalde-Mamani et al. no realizaron una búsqueda exhaustiva de secuencias homologas depositadas en GenBank, por lo que no incluyeron la secuencia de cyt-b con el nombre de Liolaemus sp. “Parinacochas” (Aguilar-Puntriano et al. 2018). En este estudio se analiza esta secuencia con las de L. warjantay y otras especies del grupo montanus de Perú, y se detalla la variación morfológica y patrón de coloración de la nueva población de L. warjantay procedente del departamento de Ayacucho.
Material y métodos
Las secuencias seleccionadas de especies del grupo montanus y del marcador citocromo oxidasa b (cyt-b) fueron obtenidas de GenBank (Tabla 1). Las secuencias fueron alineadas usando el plugin MUSCLE alignment en GENEIOUS PRIME 2023.0.4 (https://www.geneious.com). El alineamiento se analizó usando el método del vecino más próximo (“neighbor joining” o NJ) con el modelo de distancia de Jukes Cantor. Se utilizó a Liolaemus cf. ornatus (grupo darwini, subgénero Eulaemus) como grupo externo. Los valores de soporte se obtuvieron llevando a cabo un remuestreo de bootstrap con mil replicaciones. El análisis filogenético como el de bootstrap se hicieron en GENEIOUS PRIME 2023.0.4. Se estimaron distancias genéticas desde el árbol de NJ a partir del plugin Species Delimitation 1.4.5 en GENEIOUS PRIME 2023.0.4.
Tabla 1 Secuencias de cyt-b utilizadas por taxón en este estudio con su respectivo número de acceso de Genbank y de Museos de Historia Natural. BYU = Brigham Young University, MUBI = Museo de la Biodiversidad, MUSA = Museo Universidad San Agustín, MUSM = Museo Universidad Nacional Mayor de San Marcos.

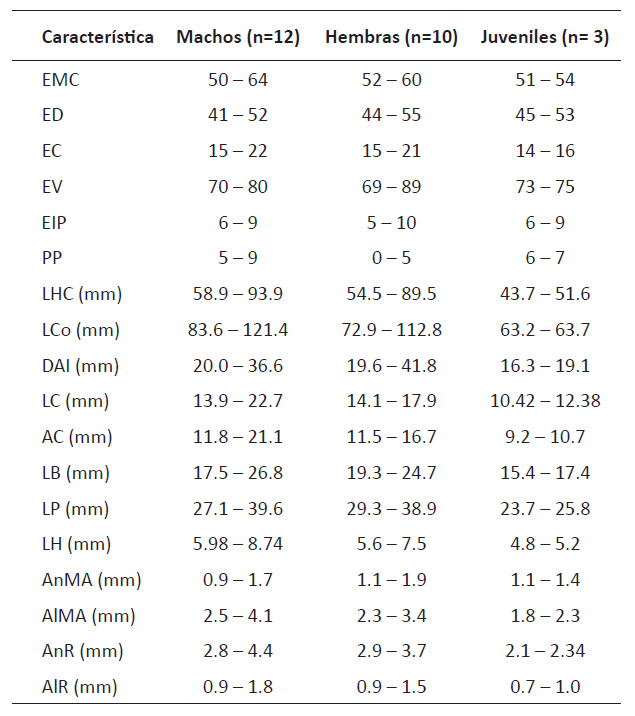
Se revisaron 25 especímenes de Liolaemus warjantay que se encuentran depositados en el Departamento de Herpetología del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MUSM). Se sigue a Smith (1946) y a Frost (1992) para la definición de las características evaluadas y la terminología de los pliegues del cuello respectivamente. Se tomaron las siguientes mediciones y conteos: 1) (LHC) longitud hocico-cloaca; 2) (LCo) longitud de la cola; 3) (DAI) distancia de la axila a la ingle (entre la inserción posterior del miembro anterior y la inserción anterior del miembro posterior); 4) (LC) longitud de la cabeza (entre la punta de la cabeza al borde anterior del oído) ; 5) (AC) ancho de la cabeza (en su ancho máximo); 6) (LB) longitud del brazo (distancia desde la inserción del miembro anterior al cuerpo hasta el término del cuarto dígito); 7) (LP) longitud de la pierna (distancia desde la inserción del miembro posterior al cuerpo hasta el término del cuarto dígito); 7) (LH) longitud del hocico (desde la comisura de la boca hasta el extremo de la cabeza); 8) (AnMA) ancho del meato auditivo; 9) (AlMA) altura del meato auditivo; 10) (AnR) ancho del rostral; 11) (AlR) altura del rostral; 12) (EMC) escamas alrededor del medio cuerpo (contadas transversalmente en la mitad del cuerpo); 13) (ED) escamas dorsales (desde el nivel del borde anterior del oído hasta el nivel del borde anterior de los miembros posteriores); 14) (EC) escamas de la cabeza (desde la escama rostral hasta el nivel del borde interno del oído); 15) (EIP) escamas alrededor de la interparietal; 16) (EV) escamas ventrales (desde la escama mental hasta la cloaca); 14) (PP) poros precloacales en machos y hembras. La comparación del patrón de coloración se basó en fotografías de las lagartijas tomadas en vida y la descripción original de L. warjantay (Ubalde-Mamani et al. 2021). Las mediciones fueron tomadas con un caliper de precisión digital hasta el 0.01 mm usando el lado derecho de los especímenes. Para el conteo de escamas se usó un estereoscopio (10x-40x).
Resultados y discusión
La identificación molecular (Fig. 1) en base a secuencias de cyt-b de especies del grupo montanus de Perú muestra que el individuo MUSM 26393 se agrupa con los individuos de L. warjantay formando un clado con un soporte de 100. Por lo tanto, el individuo identificado como Liolaemus “Parinacochas” en Aguilar-Puntriano et al. (2018) corresponde a L. warjantay, pero no tomado en consideración por Ubalde-Mamani et al. (2021).
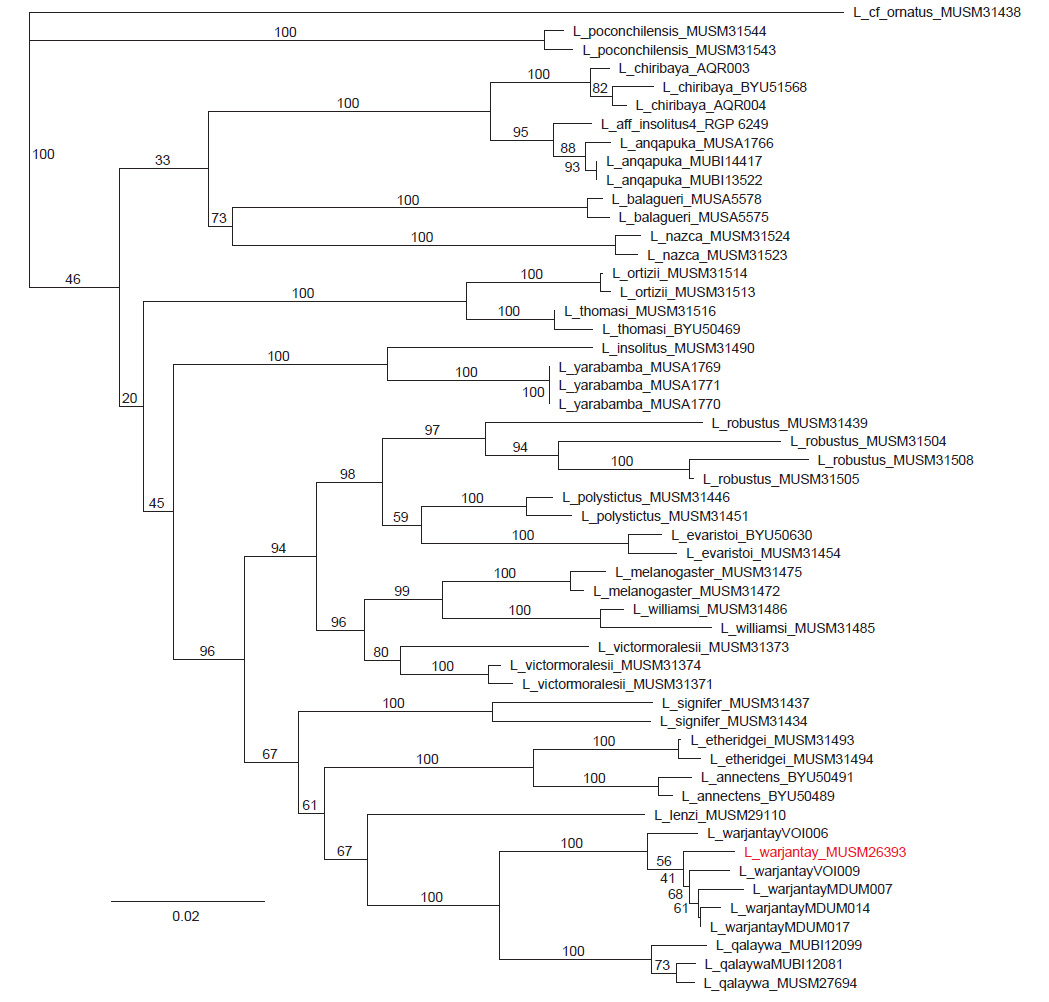
Figura 1 Arbol de Neighbor Joining en base a secuencias de cytb mostrando (en rojo) al individuo de Liolaemus warjantay procedente del distrito de Chumpi, provincia de Parinacochas, departamento de Ayacucho. Los valores de soporte de bootstrap ≥ 70 se muestran en las ramas.
El nuevo registro de Liolaemus warjantay se encuentra en dos localidades cercanas, ambas ubicadas en el distrito de Chumpi, provincia de Parinacochas, departamento de Ayacucho, con coordenadas -15.096 S, -73.664 W, y -15.142 S, -73.73 W. Este nuevo registro extiende la distribución de L. warjantay 76 km al noroeste (Fig. 2). La distribución altitudinal en la nueva localidad es desde 3484 hasta 4088 m, extendiendo su rango altitudinal entre los 3484 - 4529 m. El ecosistema de la nueva localidad es Pajonal de Puna Seca (Fig. 4). Todos los especímenes fueron encontrados debajo de piedras.
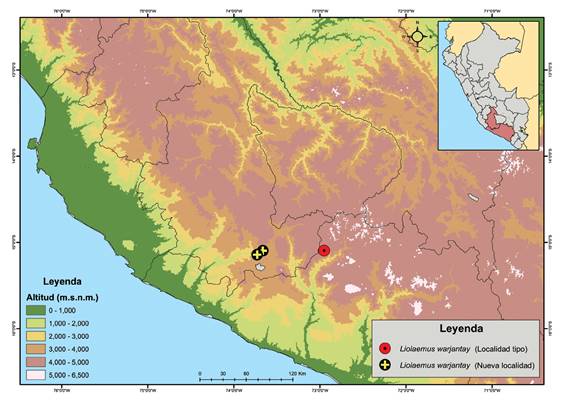
Figura 2 Mapa mostrando el nuevo registro de Liolaemus warjantay procedente del distrito de Chumpi, provincia de Parinacochas, departamento de Ayacucho.
La Tabla 2 muestra la variación merística y morfométrica encontrada en las muestras del departamento de Ayacucho. Aunque Ubalde-Mamani et al. (2021) solo distingue variación entre machos y hembras para unas pocas características en las localidades de Arequipa, las principales diferencias merísticas y morfométricas se presentan con relación al número máximo de escamas dorsales y ventrales del cuerpo (63 y 85 respectivamente en la muestra de Arequipa), escamas de la cabeza (hasta 18 en Arequipa), y en la longitud de la cola (longitud máxima menor en Arequipa tanto en machos como en hembras). La diferencia entre la distancia máxima de la axila a la ingle entre las muestras de Arequipa (71.9 mm) reportado por Ubalde-Mamani et al. (2021) y las de Ayacucho (41.8 mm) es considerable por lo que estimamos que existe un error de medición en las muestras de Arequipa.
Tabla 2 Variación morfométrica y merística de la nueva población de Liolaemus warjantay procedente de Ayacucho. Ver Material y Métodos para las abreviaturas de las características.
El patrón de coloración de los especímenes de la nueva localidad es bastante similar a los ejemplares de la localidad típica (Ubalde-Mamani et al. 2021). Los individuos de la nueva localidad presentan manchas oscuras paravertebrales bordeadas con una línea clara en la parte anterior que da la impresión de ocelos, y manchas laterales parecidas, pero menos diferenciadas. Las manchas son visibles y claras en los ejemplares juveniles, pero desaparecen con la edad y ejemplares de mayor tamaño pueden ser casi uniformes sin huellas de manchas (Figs. 3A - C). La coloración de la población en la localidad típica es en general más contrastante y colorida. El color base es gris amarillento en contraste con la población nueva que tiene base gris o gris-amarillento muy leve. También las escamas palpebrales son amarillentas en la población de la localidad típica, pero grises en la nueva localidad. La parte oscura de la cabeza es más intensa y tiene mayor extensión en los individuos en la localidad típica que en la nueva localidad. De la misma forma, el vientre de los especímenes de la localidad típica es más oscura que en la nueva localidad.

Figura 3 (A) Macho (MUSM 31470), (B) hembra (MUSM 31469) y (C) juvenil (MUSM 31471) de Liolaemus warjantay procedente del distrito de Chumpi, provincia de Parinacochas, departamento de Ayacucho. Fotografía de M. Lundberg.

Figura 4 Hábitat de Liolaemus warjantay en el distrito de Chumpi, provincia de Parinacochas, departamento de Ayacucho. Fotografía de M. Lundberg.
En los últimos cuatro años se ha incrementado considerablemente el conocimiento taxonómico del género Liolaemus en Perú, describiéndose 10 especies tanto en ecosistemas de costa y andinos (Aguilar-Puntriano et al. 2019, Arapa-Aquino et al. 2021, Chaparro et al. 2020, Huamaní-Valderrama et al. 2020, Quiroz et al. 2021, Ubalde-mamani et al. 2021, Valladares-Faúndez et al. 2021, Villegas Paredes et al. 2020); sin embargo, algunas de las hipótesis taxonómicas realizadas en estos trabajos necesitan revisión. Por ejemplo, para proponer a L. yarabamba Arapa-Aquino et al. 2021 y L. basadrei Valladares-Faúndez et al. 2021 como linajes independientes solo se incluyó evidencia morfológica y del patrón de coloración (Arapa-Aquino et al. 2021, Valladares-Faúndez et al. 2021). Aunque una sola línea de evidencia puede ser necesaria para proponer un nuevo linaje, en taxa como Liolaemus, con casos de fuerte polimorfismo intraespecífico y de convergencia, una sola evidencia puede ser insuficiente y se podrían estar repetitiendo los errores cometidos en la historia taxonómica de Liolaemus en Perú (Aguilar et al. 2017, Aguilar-Puntriano et al. 2018, CA observación personal). Las descripciones de L. polystictus Laurent 1992 y L. melanogaster Laurent 1998, las cuales también se basaron solamente en una fuente de evidencia, ilustran muy bien estos errores. Los individuos usados en la descripción de estas dos especies provenían de dos localidades, pero posteriormente los individuos de una de las localidades terminaron correspondiendo a un taxón diferente (Laurent 1992, 1998; Aguilar et al. 2017). En el caso de L. polysticus, seis paratipos tienen como localidad Castrovirreyna, pero después fueron separadas por Aguilar et al. (2017) y luego descritas como L. evaristoi Gutierrez et al. 2018. Mientras que en la descripción de L. melanogaster, dos paratipos provenían de Abra Apacheta. Sin embargo, Aguilar et al. (2017) demostraron que los individuos de Liolaemus de esta localidad estaban más relacionados a L. polystictus y L. evaristoi, pero no a L. melanogaster.
Aunque, en otras hipótesis taxonómicas como las de L. ankapuka Huamaní-Valderrama et al. 2020 y L. yarabamba Quiroz et al. 2021, éstas se basaron en más de una fuente de evidencia, un reanálisis de los datos moleculares podría resultar en la sinonimia de L. yarabamba y L. insolitus, y de L. ankapuka y L. chiribaya. Si bien, no es objetivo de este trabajo realizar el mencionado reanálisis, una estimación de las distancias interespecíficas a partir del árbol de Neighbor Joining obtuvo que las distancias entre L. yarabamba y L. insolitus es de 4.9% y entre L. ankapuka (incluyendo a “L. aff. insolitus 4”) y L. chiribaya es de 3.2%. En comparación, las distancias entre dos taxones hermanos como L. balagueri y L. nazca es de 10%, a pesar de que son especies fenotípicamente muy similares (Aguilar-Puntriano & Salazar 2021) Quiroz et al. (2021) muestran una tabla con distancias genéticas (el procedimiento para estimarlas no se menciona en sus métodos), pero la única comparación interespecífica que proporcionan es entre L. insolitus y L. yarabamba, con valores similares al encontrado, estimando distancias desde el árbol. En el caso de L. ankapuka, Huamaní-Valderrama et al. (2020) no llevan a cabo una estimación de distancias genéticas ni ningún otro análisis molecular adicional, pero concluyen que como los terminales de la nueva especie forman un grupo monofilético esto da soporte a la evidencia fenotípica de la existencia de un linaje independiente. Como se mencionó en Aguilar-Puntriano & Salazar (2021) la presencia de un grupo monofilético por sí mismo no es evidencia suficiente para considerar a una población aislada de Liolaemus como un linaje independiente, cuando también puede inferirse que es conespecífica con un taxón ya descrito como lo sugiere la distancia genética entre L. chiribaya y L. ankapuka. Las diferencias fenotípicas entre L. ankapuka y L. chiribaya como entre L. insolitus y L. yarabamba también podrían inferirse como diferencias entre poblaciones y no interespecíficas, pero esto amerita un estudio más amplio que está más allá de los objetivos de este trabajo.

