











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El género Hylephila Billberg, 1820 (Hesperiidae: Hesperiinae, Hesperiini, Hesperiina), contiene actualmente 21 especies válidas, de pequeño tamaño, que se distribuyen a lo largo de la Cordillera de los Andes en América del Sur (MacNeill & Herrera 1998, MacNeill 2002, Mielke 2005), con la única excepción de la especie H. phyleus (Drury, 1773), cuya distribución abarca casi todo el continente americano, desde Canadá hasta el sur de Chile y Argentina, incluyendo la costa atlántica de Brasil, las islas del Caribe y -artificialmente introducida- el archipiélago de Hawaii (Hayward 1973, Tashiro & Mitchell 1985, Cock 2007, Clements 2012, Giovenardi et al. 2013), siendo reconocidas ocho subespecies válidas: ancaEvans, 1955, andina (Staudinger, 1894), basistrigata (Eaton, 1932), eureka Austin & Emmel, 1998, muertovalle Scott, 1981, monticola Hayward, 1939, phyleus, y taxus Evans, 1955 (Mielke 2005).
MacNeill & Herrera (1998) separaron el género Hylephila en cuatro grupos con base en caracteres morfológicos: boulleti, ignorans, venusta, y phyleus; en ese mismo trabajo revisaron los grupos ignorans y venusta. Posteriormente, MacNeill (2002) revisó el grupo boulleti, describiendo varias nuevas especies. Lamentablemente, durante el estudio del último grupo, phyleus, MacNeill falleció a la edad de 80 años debido a una insuficiencia cardiaca (Brown et al. 2006), por lo que la publicación de la tercera y última parte de la revisión de Hylephila quedó trunca.
Hylephila phyleus basistrigata fue originalmente descrita dentro del género Talides Hübner, [1819] por Eaton (1932), con base en seis machos y una hembra, material recolectado por W. T. M. Forbes en la ciudad de Lima (Perú) y sus alrededores, durante la expedición entomológica efectuada por la Universidad de Cornell (E. U. A.) en 1919-1920 (Lindsey 1925). Bridges (1988), en su catálogo de Hesperiidae, retuvo esta especie dentro del género Talides, a pesar de que Evans (1955) ya la había reducido a una subespecie de Hylephila phyleus, argumentando que basistrigata ocurre en ambientes más secos que aquellos habitados por phyleus, sin mayor elaboración. Aunque Pallister (1956), Pérez-D’Angello (1965) y Hayward (1973) citaron -sin mayor explicación- a basistrigata como una especie de Hylephila, Mielke (2005) mantuvo el estatus taxonómico propuesto por Evans, utilizado históricamente para designar a las poblaciones afines a phyleus que habitan en la vertiente occidental de los Andes desde Ecuador hasta Chile, a elevaciones entre el nivel del mar y 3,000 m (Evans 1955, Ureta 1963, Lamas 1977, Ruiz 1977, Peña & Ugarte 1997, Benyamini et al. 2014, Farfán 2018).
Recientemente, durante el desarrollo de un proyecto de investigación cuyo objetivo era evaluar lepidópteros en el suroeste de Perú, examinamos ejemplares de Hylephila pertenecientes al grupo phyleus (MacNeill 2002), recolectados en el Departamento de Arequipa, y descubrimos diferencias notorias en las estructuras internas (genitalia masculina) entre ejemplares inicialmente identificados como una sola especie, H. phyleus basistrigata, con base en su patrón alar y lugares de recolección, presumiéndose así la existencia de dos entidades taxonómicas crípticas con similar apariencia externa. Luego de revisar minuciosamente las imágenes de la genitalia masculina de basistrigata y phyleus en Eaton (1932), se llegó a la conclusión que un grupo de los individuos examinados correspondería a H. phyleus y otro a H. basistrigata. Por lo tanto, el objetivo del presente trabajo es restituir como especie válida a H. basistrigata con base en un análisis integrativo con aspectos morfológicos, ecológicos y genéticos.
Material y métodos
Material examinado. Ejemplares de Hylephila phyleus (sensu lato) depositados en las colecciones entomológicas del Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Perú (MUSM) y Museo de Historia Natural, Universidad Nacional de San Agustín de Arequipa, Arequipa, Perú (MUSA), así como la serie típica de Talides basistrigata, depositada en la Cornell University Insect Collection, Ithaca, New York, USA (CUIC).
Análisis morfológico. Para el examen de la genitalia se procedió a remover los abdómenes de los ejemplares y colocarlos en una solución de hidróxido de potasio (KOH) al 10% por 10 min en baño maría a 60 °C. Posteriormente, cada abdomen fue limpiado en agua y se removió los tejidos blandos para exponer la genitalia utilizando pinzas finas, luego esta última fue sumergida por unos minutos en solución de Negro de Clorazol para identificar las partes blandas. El agua fue extraída de la genitalia disectada utilizando soluciones graduadas de etanol al 90 y 95%, y finalmente la genitalia fue almacenada en microviales con glicerina. La terminología utilizada sigue a MacNeill (1993, 2002). Las medidas están en milimetros y fueron realizadas con base en las fotografías de los ejemplares cerca de una escala y aumentadas en una pantalla de computadora. Las fotografías de adultos fueron tomadas con una cámara digital Nikon D5600; las de la genitalia fueron tomadas con una cámara Nikon DS-Ri2 a través de un estereoscopio Nikon SMZ25. Las imágenes fueron ensambladas y editadas en Photoshop CS5.1.
Análisis genético. El ADN genómico fue extraído de tejido abdominal de dos ejemplares (una hembra de H. phyleus y una hembra de H. basistrigata) siguiendo los procedimientos descritos en Huanca-Mamani et al. (2015). Un fragmento del gen COI fue amplificado por Reacción en Cadena de Polimerasa (PCR) utilizando los cebadores LEP-F1 y LEP-R1 (Hebert et al. 2004). Las reacciones PCR fueron efectuadas en un volumen final de 20 μL.; cada reacción contuvo 1 μL de ADN extraído, 10 ρmoles de cada cebador, 2.5 mM de cada dNTP, 2 mM MgCl2, 1X PCR buffer (KCl), 1 unidad de Taq DNA polimerasa (ThermoScientific) y agua destilada estéril. La amplificación se realizó con el siguiente programa: 5 min a 94 °C, 35 ciclos de 30 s a 94 °C, 30 s a 47 °C, 1 min a 72 °C, y un paso final de elongación de 10 min a 72 °C. Luego, 3 μL de cada producto PCR fue visualizado sobre 1.5% de gel agarosa teñido con gel-red (Biotium). Las reacciones que contuvieron fragmentos del tamaño esperado fueron purificadas y secuenciadas directamente por una empresa comercial (Macrogen, Corea del Sur). Para explorar las relaciones filogenéticas de Hylephila y su clasificación combinamos nuestros datos obtenidos del gen COI con una base de datos de secuencias depositadas en GenBank y Bold Systems (Tabla 1). Las secuencias se alinearon mediante el algoritmo Clustal W (Thompson et al. 1994) implementado en el software MEGA X (Kumar et al. 2018). Un análisis filogenético de máxima verosimilitud (ML) fue realizado con MEGA X. La búsqueda de árboles fue efectuada con el modelo de sustitución GTR + G + I (determinado sobre la base de criterio de información Akaike), y 1000 replicaciones bootstrap.
Tabla 1 Ejemplares usados para el análisis genético de Hylephila en el presente estudio.
| Especie | País / Localidad | Voucher | Código de acceso Genbank / Bold Systems | Referencia |
|---|---|---|---|---|
| Hylephila basistrigata | Perú / Arequipa | E1786_Sg313 | No asignado | Este estudio |
| Hylephila phyleus | Perú / Arequipa | E1787_Sg313 | No asignado | Este estudio |
| Hylephila phyleus | Colombia | BMC19987 | MN306972.1 | Marín et al. 2021 |
| Hylephila phyleus | Colombia | BMC19510 | MN306952.1 | Marín et al. 2021 |
| Hylephila phyleus | Argentina / Buenos Aires | MACN-Bar-Lep-ct 03062 | MF546989.1 | Lavinia et al. 2017 |
| Hylephila phyleus | Argentina / Jujuy | MACN-Bar-Lep-ct 06736 | MZ335625.1 | Attiná et al. 2021 |
| Hylephila phyleus | Argentina / Corrientes | MACN-Bar-Lep-ct 03593 | MF547053.1 | Lavinia et al. 2017 |
| Hylephila phyleus | Argentina / Salta | MACN-Bar-Lep-ct 07709 | MZ335587.1 | Attiná et al. 2021 |
| Hylephila phyleus | Canada / Ontario | BIOUG-10890-C06 | CNCBF315-14 | ‒ |
| Hylephila phyleus | USA / Texas | ‒ | HQ583472.1/ USLEP1367-10 | ‒ |
| Hylephila phyleus | USA / Florida | ‒ | HQ583475.1 / USLEP1370-10 | ‒ |
| Hylephila phyleus | USA / Florida | ‒ | LOFLC160-06 | ‒ |
| Hylephila phyleus | USA / Florida | ‒ | LOFLC162-06 | ‒ |
| Hylephila phyleus | USA / Georgia | ‒ | MOBIL1441-16 | ‒ |
| Hylephila phyleus | Chile / Malleco | 630-AVZB | EU364467.1 | Warren et al. 2008 |
| Hylephila ancora | Argentina / Tucumán | MACN-Bar-Lep-ct 07518 | MZ335849.1 | Attiná et al. 2021 |
| Hylephila ancora | Argentina / Tucumán | MACN-Bar-Lep-ct 07516 | MZ335941.1 | Attiná et al. 2021 |
| Hylephila ancora | Argentina / Tucumán | MACN-Bar-Lep-ct 07520 | MZ335381.1 | Attiná et al. 2021 |
Muestreo en el campo. Para determinar el comportamiento de oviposición y confirmar la planta hospedera utilizada por las poblaciones de las dos especies H. phyleus e H. basistrigata en el departamento de Arequipa, se seleccionó la localidad de Yumina (16°26’S, 71°28ʹW), ubicada a 15 km al Este de la ciudad de Arequipa, a una altitud de 2500 m. Debido a que se encontraron depositados en la colección del MUSA ejemplares de ambas especies con datos de recolección de dicho lugar y con las mismas fechas, se pudo inferir que utilizaban el mismo hábitat u ocurrían en dicha localidad en la misma época de vuelo. Por lo tanto, se realizaron visitas a la localidad de Yumina entre los meses de abril y junio de 2022. En cada visita se efectuaron recorridos en el lugar, entre las 08:00 y 13:00 h, abarcando todos los ambientes posibles para ubicar individuos de phyleus y basistrigata. Para confirmar su identificación respecto al resto de especies presentes se utilizaron binoculares o los individuos fueron capturados temporalmente con una red entomológica para examinar su patrón alar y posteriormente ser liberados. Previamente se había determinado con los ejemplares depositados en la colección de MUSA características de su patrón alar, principalmente de la cara ventral del ala posterior, y que diferenciaban a los individuos de phyleus y basistrigata que habitan en los alrededores de la ciudad de Arequipa (16°23’S, 71°31’W). Luego de ubicar los individuos e identificar las zonas por donde se desplazaban, se efectuó un seguimiento de las hembras, las cuales podían ser distinguidas de los machos por el tipo característico de vuelo errático al ras del suelo, en aparente búsqueda de lugares de oviposición, comportamiento descrito por Porter (1992) y con la ayuda de binoculares se observó directamente la oviposición, confirmando de esta manera la planta hospedera de los estadios inmaduros.
Resultados
El análisis filogenético realizado con las secuencias del gen COI obtenidas de los ejemplares de Hylephila de Arequipa (Fig. 1) revela que las dos poblaciones (phyleus y basistrigata) que vuelan en la misma localidad presentan una distancia genética mayor al 3% entre ellas (Material Suplementario 1), lo que confirmaría que son especies distintas. La especie denominada aquí como H. phyleus de Arequipa se agrupa con otras entidades también identificadas como H. phyleus provenientes de Norteamérica y Sudamérica con distancias genéticas menores al 1%, con excepción de una secuencia proveniente de Chile (Tabla 1), la cual se separa tanto de H. phyleus como de H. basistrigata (Fig. 1), con distancia genética mayor al 3% (Material Suplementario 1), esto debido probablemente a un error en su identificación como H. phyleus. Sin embargo, los resultados obtenidos apoyan la hipótesis que H. basistrigata es una especie válida y la población de H. phyleus que habita la vertiente occidental de los Andes de Perú y Chile corresponde a una probable subespecie inédita que no será descrita aquí por no ser el objetivo del presente trabajo, pero por razones de diferenciación será citada aquí como H. phyleus ssp. A continuación, se brinda una redescripción de H. basistrigata con base en los resultados obtenidos en el análisis morfológico.

Figura 1 Árbol filogenético de máxima verosimilitud en base a secuencias del gen citocromo c oxidasa I (COI) de ejemplares de Hylephila, mostrando el estado monofilético de H. basistrigata.
Hylephila basistrigata (Eaton, 1932) stat. rest.
Material examinado (todos con preparación de genitalia): Lectotipo ♂ (designado aquí, depositado en CIUC). Material adicional (18♂, 8♀): 3♂, 1♀, PERU, AR [Arequipa], Prov. Arequipa, Yura, S 16°14´50” / W 71°42´01”, 2480 m, 18.vi.2014, Leg. Erick Huamaní (MUSA); 2♂, 1♀, PERU, AR [Arequipa], Prov. Arequipa, Yumina, S 16°26´25” / W 71°28´48”, 2500 m, 12.vii.2014, Leg. G. Apaza (MUSA); 1♂, PERU, AR [Arequipa], Prov. Caravelí, Jaqui río Yauca, S 15°28´43” / W 74°26´53”, 200 m, 21.iii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 2♂, PERU, AR [Arequipa], Aplao, Valle Majes, S 15°53´46” / W 72°28´03”, 800 m, 02.vi.2014, Leg. Marco Delgado (MUSA); 1♂, PERU, AR [Arequipa], Prov. Arequipa, 7 Km SE Pueblo Yumina, S 16°26´31” / W 71°28´53”, 2500 m, 26.vii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 1♂, 1♀, PERU, AR [Arequipa], Prov. Camaná, Characta río Camaná, S 16°30´48” / W 72°38´27”, 100 m, 16.iii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 1♀, PERU, TA [Tacna], Ite, ruta Ilo-Tacna, S 17°54´29” / W 70°57´25”, 30 m, 29.x.2015, Leg. Jose Cerdeña (MUSA); 2♀, PERU, AR [Arequipa], Prov. Castilla, Uraca-Corire, S 16°21´11” / W 72°29´19”, 307 m, 24.viii.2014, Leg. G. Apaza (MUSA); 1♀, PERU, AR [Arequipa], Prov. Arequipa, Yarabamba, S 16°33´01” / W 71°28´18”, 2477 m, 17.v.2014, Leg. G. Apaza (MUSA); 1♀, PERU, AR [Arequipa], Prov. Arequipa, Yarabamba, S 16°33´07” / W 71°27´56”, 2503 m, 01.vii.2014, Leg. G. Apaza (MUSA); 1♂, PERU, LI [Lima], Végueta, 10-15 m, 1101/7739 [S 11°01´ / W 77°39´], 27.ii.1995, J. Grados (MUSM); 1♂, PERU, La Molina [Lima], 250 m, 01.v. 1974, G. Lamas (MUSM); 1♂, PERU, IC [Ica], Ica, 400 m, 02.x.1984, G. Lamas & E. Pérez (MUSM); 1♂, PERU, AR [Arequipa], Yura, S 16°14´48” / W 71°41´56”, 2500 m, 11.vii.2014, Leg. G. Apaza (MUSM); 1♂, PERU, Pi [Piura], Península Illescas, 0 m, v.84[1984], T. Torres (MUSM); 1♂, PERU, AR [Arequipa], Characato, xii.82 [1982], G. Lamas (MUSM); 1♂, PERU, LI [Lima], Pantanos de Villa, 30 m, 1212/7659 [S 12°12´ / W 76°59´], 22.ii.1997, J. Grados (MUSM); 1♂, IQUIQUE, prov. Tarapacá, col. Etcheverry, Cent.Ent.U., CHILE (MUSM).
Otro material examinado (todos con preparación de genitalia): Hylephila phyleus ssp. (30♂, 2♀): 5♂, PERU, AR [Arequipa], Aplao, Valle Majes, S 15°53´46” / W 72°28´03”, 800 m, 02.vi.2014, Leg. Marco Delgado (MUSA); 1♂, PERU, AR [Arequipa], Prov. Arequipa, Yura viejo, S 16°13´02” / W 71°42´17”, 2600 m, 30.iii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 3♂, PERU, AR [Arequipa], Prov. Arequipa, Yumina, S 16°26´25” / W 71°28´48”, 2500 m, 03.v.2014, Leg. G. Apaza / Percy (MUSA); 1♂, PERU, AR [Arequipa], Prov. Caravelí, Yauca río Yauca, S 15°40´13” / W 74°31´27”, 50 m, 19.iii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 1♂, 1♀ PERU, AR [Arequipa], Prov. Caravelí, Atiquipa, lomas costeras, S 15°47´33” / W 74°21´58”, 300 m, 18.iii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 1♂, PERU, AR [Arequipa], Prov. Camaná, ca. puente río Camaná, S 16°34´57” / W 72°43´47”, 50 m, 15.iii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 1♂, PERU, AR [Arequipa], Prov. Arequipa, Vítor, campos de cultivo, S 16°27´52” / W 71°55´37”, 1150 m, 28.viii.2008, Leg. J. Cerdeña / J. Farfán (MUSA); 1♂, 1♀, PERU, AR [Arequipa], Aplao, Valle Majes, S 15°53´46” / W 72°28´03”, 800 m, 02.vi.2013, Leg. J. Cerdeña / Marco Delgado (MUSA); 1♂, PERU, AR [Arequipa], Arequipa, río Chili 5 Km N Chilina, S 16°21´41” / W 71°32´14”, 2500 m, 24.v.2013, Leg. Jackie Farfán (MUSA); 1♂, PERU, La Molina [Lima], 250 m, 01.v.1974, G. Lamas (MUSM); 1♂, PERU, Tumbes, 10 Km W El Huásimo, 500 m, 14.ii.1976, G. Lamas (MUSM); 1♂, PERU, Lima, Surco, 2000 m, 22.iv.1974, G. Lamas (MUSM); 1♂, PERU, Lima, Chosica, 800 m, 04.ix.1963, P. Hocking (MUSM); 1♂, PERU, CA [Cajamarca], Baños del Inca, 2700 m, 05.iii.1985, G. Lamas (MUSM); 1♂, PERU, Lima, Antioquia, 1450 m, 14.ix.1969, G. Lamas (MUSM); 1♂, PERU, AN [Ancash], SE de Caraz, 0904/7747 [S 09°04´/ W 77°47´], 2265 m, 23.vii.2006, C. Peña (MUSM); 1♂, PERU, AN [Ancash], Puente Pacosbamba, 0854/7722 [S 08°54´/ W 77°22´], 2500 m, 20.vii.2006, G. Lamas (MUSM); 1♂, PERU, AN [Ancash], 12 Km N San Luis (puente), 2550 m, 06.x.1982, G. Lamas & E. Pérez (MUSM); 1♂, PERU, LI [Lima], La Molina, Universidad Agraria La Molina, S 12°05´12” / W 76°56´54”, 239 m,11.iii.2016, E. Gamboa (MUSM); 1♂, PERU, MO [Moquegua], Moquegua, 1350 m, 17.x.1986, G. Lamas (MUSM); 1♂, PERU, LI [Lima], Quintay, 7 Km E Sayán, 850 m, 04-06.v.1982, G. Lamas & E. Pérez (MUSM); 1♂, PERU, CA [Cajamarca], 1 Km puente Papayo, ca. Hda. Monteseco, 550 m, 17.v.1982, G, Lamas & E. Pérez (MUSM); 1♂, PERU, CA [Cajamarca], Chamaya, 500 m, 13.x.1985, G, Lamas (MUSM); 1♂, Valle Sta. Eulalia (cerca Lima) [PERU, Lima], 2180 m, vi.1953, Coll. F. Blancas (MUSM). Hylephila phyleus phyleus: 1♀, GUADELOUPE, Basse-Terre, Saint-François, Anse a l´Eau, 16.xii.2000, C. Brévignon (MUSM). Hylephila phyleus andina: 1♂, PERU, CU [Cuzco], Calca, Quebrada Quellouno, 2950 m, 1321-2/7157-8 [S 13°21-22´ / W 71°57-58´], 10.ii.2001, J. L. Curo (MUSM).
Descripción. Macho (Figs. 2A-F). Cabeza. Dorsalmente con abundante pilosidad naranja, un penacho de pelos negros cubriendo los ojos. Antenas marrones con el ápice naranja principalmente en la cara ventral. Palpos con pilosidad naranja, siendo más claro en la parte basal en vista ventral, pero con pequeños pelos negros en la base. Tórax. Marrón obscuro con collar (patagia) naranja similar al de la cabeza y pilosidad lateral abundante naranja en vista dorsal, pilosidad más clara en vista ventral. Alas. Longitud del ala anterior (10-14 mm, n=18). Cara dorsal con estigma presente, masa micro androconial negra, mancha post-estigmal negra y ancha, coloración naranja con base alar negra alcanzado hasta el estigma, banda marginal negra con prolongaciones internas triangulares entre las venas, pequeña mancha negra postdiscal cerca del ápice de la celda discal. Cara ventral con base negra hasta la celda discal conectándose con pequeña mancha negra ubicada a la altura de la mancha androconial de la cara dorsal pero menos extensa, margen anal negro paralelo a la vena anal, ápice de la celda discal con una línea negra sobre el margen, las manchas menos notorias que en la cara dorsal y variando de extensión e intensidad entre los ejemplares examinados. Ala posterior naranja similar al ala anterior, cara dorsal con banda marginal negra y prolongaciones triangulares menos extensas que las del ala anterior, área basal negra y una notoria línea negra partiendo de la base cerca del margen costal, bifurcándose a la altura del ápice de la celda discal y alcanzando el inicio de la vena M1; margen anal con franja negra abarcando casi todo el espacio entre la vena 2V y el margen anal con pilosidad de color naranja; cara ventral naranja con una mancha marcada negra iniciando en la base alar y atravesando longitudinalmente la celda discal como una línea continua hasta el margen discal, en algunos ejemplares dividida casi al finalizar su recorrido en dos franjas una más larga y otro de pequeño tamaño como un punto negro, en algunos ejemplares se observa un patrón de manchas negras en el área postdiscal ubicadas entre los espacios R-M1, M2-M3, M3-Cu1 y en el área submarginal en los mismos espacios, siendo variables en número e intensidad entre los ejemplares examinados. Abdomen. Marrón obscuro en vista dorsal, y blanco-crema en vista ventral, con escasa pilosidad naranja en ambos lados. Genitalia (Figs. 3A-B). Valva ancha en vista lateral, con margen ventral ligeramente convexo, con pliegue interno con dos invaginaciones formando una pequeña elevación ligeramente pronunciada hacia la cara dorsal, margen caudal dividido en dos ápices anchos dentados. Uncus con dos espinas dorsolaterales cerca a la base, margen dorsal ligeramente pronunciado hacia arriba en vista lateral, margen ventral con pronunciada hendidura hacia arriba cerca de la base en vista lateral y las pectinas (para una definición de “pectinas” ver MacNeill 1993) con proyección recta. Gnathos ligeramente esclerotizado, delgado, en vista lateral no sobrepasa las pectinas. Tegumen ligeramente redondeado. Edeago ligeramente curvado hacia la cara dorsal, con una hendidura distal en el centro en vista dorsal, cuatro cornuti en forma de espina: dos cortos laterales y dos largos. Hembra (Figs. 2G‒H).Similar al macho pero con manchas más pronunciadas y con las siguientes diferencias: Longitud del ala anterior (14‒20 mm, n=8); coloración general más tenue, ala anterior en la cara dorsal con una mancha negra en la celda discal extendiéndose hasta el espacio M3-Cu1 en el área postdiscal; ala posterior en la cara ventral manchas negras en la celda discal y área postdiscal ubicadas entre los espacios R-M1, M2-M3, M3-Cu1 mucho más pronunciadas y presentes en todos los ejemplares examinados. Genitalia (Figs. 3C-D). Octavo esternito esclerotizado, en vista ventral con una placa esclerotizada distalmente más ancha y en el extremo proximal terminando en punta ancha alcanzado el antrum, apófisis anteriores cortos y posteriores el doble del tamaño de los palpos anales. Lamella postvaginalis dividida en vista ventral. Antrum dorsalmente cóncavo y ligeramente esclerotizado, ventralmente membranoso. Ductus bursae esclerotizado, ancho, en vista ventral con constricción ductal ligera. Corpus bursae membranoso. Ductus seminalis naciendo dorsolateralmente cerca de la base del corpus bursae.

Figura 2 Adultos de Hylephila basistrigata. (A-F) machos; (G-H) hembras. Caracteres diagnósticos apuntados con flechas rojas en figura H. Izquierda: vista dorsal; derecha: vista ventral. Barra: 10 mm. (A) Perú, Arequipa, Yumina, 2500 m; (B, H) Perú, Arequipa, Aplao, 800 m; (C) Perú, Arequipa, Yura, 2480 m; (D) Perú, Ica, 400 m; (E, G) Perú, Lima, La Molina, 250 m; (F) Perú, Lima, Pantanos de Villa, 30 m.
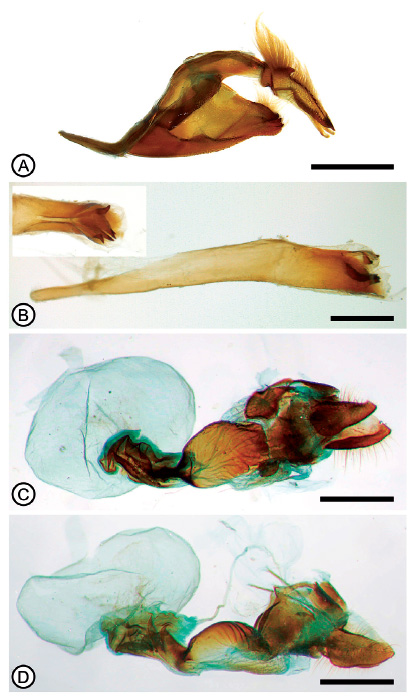
Figura 3 Genitalia de Hylephila basistrigata. (A) armadura genital, macho, vista lateral; (B) edeago, vista lateral, recuadro superior izquierdo mostrando detalle de los cornuti en vista dorsal; (C) hembra, vista ventral; (D) hembra, vista lateral. Barras: 1 mm (A, C, D), 0.5 mm (B).
Distribución. Hylephila basistrigata está restringida a la vertiente occidental de los Andes, siendo confirmada su presencia desde el norte de Perú (Tumbes) hasta el norte de Chile (Iquique), desde el nivel del mar hasta los 2,500 m de elevación, en base a los ejemplares examinados aquí.
Comentarios. El patrón alar de los ejemplares examinados, tanto de basistrigata como phyleus, presenta variaciones (Figs. 2, 4), inclusive entre ejemplares de la misma localidad, por lo que se hace difícil su identificación en el campo; sin embargo, las hembras de basistrigata mostraron una mayor consistencia en el patrón alar, especialmente en la cara ventral del ala posterior, la disposición de manchas negras ubicadas entre las áreas basales y postdiscales pueden ser consideradas como diagnósticas para diferenciarlas de otras especies afines (Fig. 2H), sobre todo dentro del área de distribución de basistrigata, donde se registran otras dos especies del grupo “phyleus”: H. phyleus ssp. e H. isonira Dyar, incluso pueden hallarse las tres especies al mismo tiempo en una localidad. Al exhibir un patrón alar variable en los individuos de phyleus e basistrigata, principalmente entre los machos, resulta necesario el examen de la genitalia para una correcta identificación de los mismos, pues hay diferencias notorias en las estructuras de la genitalia de phyleus (Fig. 5) y basistrigata (Fig. 3), principalmente en los procesos caudales de las valvas, la disposición de las pectinas y la forma del uncus en la genitalia masculina, y en la esclerotización del octavo esternito y forma del antrum en la femenina. Con tal motivo, se ofrece aquí una breve clave de identificación para separar las especies del grupo phyleus en el área de distribución de H. basistrigata (desde el norte de Perú hasta el norte de Chile):
- Macho sin espinas dorsolaterales en el uncus. Hembra con lamella postvaginalis entera H. isonira
- Macho con espinas dorsolaterales en el uncus. Hembra con lamella postvaginalis dividida 2
- Macho con pectinas del uncus proyectadas hacia abajo en vista lateral, edeago con cuatro cornuti largos. Hembra con placa esclerotizada del octavo esternito rectangular, sin alcanzar el antrum en vista ventral (Fig. 5) H. phyleus
- Macho con pectinas del uncus proyectadas rectas en vista lateral, edeago con cuatro cornuti, dos cortos y dos largos. Hembra con placa esclerotizada del octavo esternito triangular alcanzando el antrum en vista ventral (Fig. 3) H. basistrigata

Figura 4 Adultos de Hylephila phyleus. (A-D) machos; (E-H) hembras. Barra: 10 mm. (A) Perú, Arequipa, Aplao, 800 m; (B) Perú, Tumbes, Huásimo, 500 m; (C) Perú, Lima, La Molina, 239 m; (D) Perú, Arequipa, Yumina, 2500 m; (E) Perú, Lima, Jesús María, 120 m; (F) Perú, Arequipa, Atiquipa, 800 m; (G) Perú, Cusco, Calca, 2950 m; (H) Guadeloupe, Basse-Terre, 10 m.
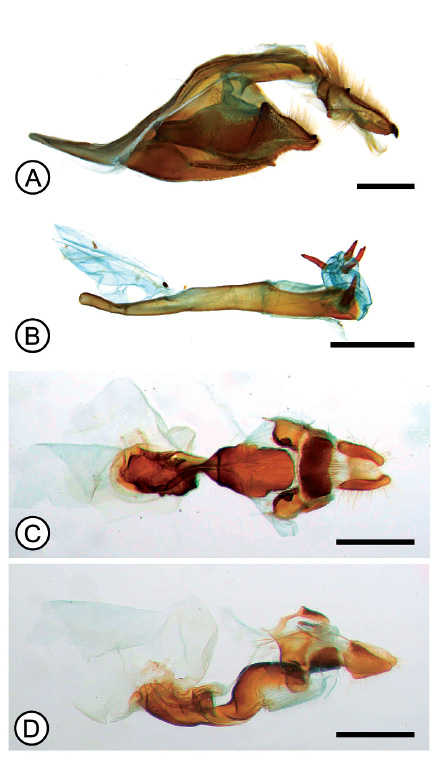
Figura 5 Genitalia de Hylephila phyleus. (A) armadura genital, macho, vista lateral; (B) edeago, macho, vista lateral; (C) hembra, vista ventral; (D) hembra, vista lateral. Barras: 0.5 mm (A), 1 mm (B-D).
Planta hospedera y comportamiento de oviposición. Durante las evaluaciones de campo en la localidad de Yumina (Fig. 6), entre los meses de abril y junio, se logró confirmar como planta hospedera de basistrigata al pasto Distichlis spicata (L.) Greene (Poaceae), conocido como “grama salada” (Figs. 6B-D). Se observó hembras ovipositando únicamente en áreas del suelo cubiertas por este pasto. Las hembras realizaban vuelos cortos y erráticos por encima del pasto, luego posaban sobre la planta y efectuaban un movimiento del abdomen típico entre las mariposas previo a la colocación de huevos (Porter 1992). Sin embargo, las hembras no colocaban los huevos sobre sustrato alguno de la planta hospedera, sino que eyectaban los huevos individualmente con dirección hacia el suelo (Material suplementario 2). Luego de eyectar un huevo, nuevamente llevaban a cabo vuelos erráticos por unos segundos o minutos, para volver a posarse y lanzar otro huevo. En un lapso de 30 min se observó que una hembra distribuyó individualmente 20 huevos en distintos puntos en un área de aproximadamente 4 m2 de pasto de D. spicata. El horario de oviposición iniciaba aproximadamente a las 10:00 h y podía durar hasta las 12:00 h en días soleados.

Figura 6 Valle de Yumina, Arequipa, Perú. (A) vista panorámica, pasto Distichlis spicata señalado con flecha roja, pasto Cynodon dactylon señalado con flecha azul; (B) Distichlis spicata; (C-D) hembra de H. basistrigata al momento de ovipositar sobre Distichlis spicata (flecha roja señalando el huevo en D); (E) Cynodon dactylon; (F-G) huevo de H. phyleus ssp. sobre Cynodon dactylon (flecha roja señalando el huevo en G).
Por otro lado, en la misma localidad se observó hembras de H. phyleus ssp. ovipositando sobre el pasto Cynodon dactylon (L.) Pers. (Poaceae). Incluso en áreas donde crecían ambos pastos (Fig. 6A), las hembras de H. phyleus ssp. colocaban los huevos individualmente en las hojas de dicha especie (Figs. 6E-G), principalmente en el envés de las hojas.
Designación de Lectotipo
Para la descripción original de basistrigataEaton (1932) empleó una serie típica de siete ejemplares: una hembra y seis machos, depositados en CUIC; al no haber sido designado un holotipo en dicho artículo, todos los ejemplares utilizados son considerados sintipos, y sólo un macho de dicha serie fue utilizado para remover el abdomen y examinar la genitalia, ilustrada en el trabajo original. Logramos examinar fotografías de los sintipos depositados en CUIC (Fig. 7), dispuestos en dos cajas entomológicas (Figs. 7A-B). La primera contiene ejemplares diversos del género Hylephila, con varios separadores (Fig. 7A) y uno de ellos contiene cinco machos sintipos de basistrigata con un rótulo en la pared del separador indicando “Talides basistrigata Eaton / PARATYPE C.U. TYPE No 1504” (Fig. 7C); otro separador anterior a éste lleva una etiqueta roja en el medio, atravesada por un alfiler, indicando “See TYPE colecction No 1504”, con un rótulo en la pared del separador indicando “Talides basistrigata Eaton / HOLOTYPE & ALLOTYPE C.U. TYPE No 1504” (Fig. 7A), dando a entender que alguna persona, en un tiempo posterior a la publicación de la descripción original, segregó los sintipos en dos grupos, cinco supuestos paratipos en una caja y en otra caja un supuesto holotipo y un supuesto alotipo depositados en la colección de tipos con el número 1504. Por lo tanto, la segunda caja fue hallada en la sección de holotipos (colección tipo número 1504), donde se encontraban separadores con ejemplares de distintos grupos de Lepidoptera albergando holotipos de distintas especies (Fig. 7B), y ahí se halló los dos restantes sintipos de basistrigata en uno de los separadores, con el rótulo “Talides basistrigata Eaton / HOLOTYPE & ALLOTYPE / C.U. TYPE No 1504”, correspondiendo a un macho y una hembra (Fig. 7 D). Ambos ejemplares coinciden plenamente con los fotografiados e ilustrados en el artículo de Eaton (1932) (Fig. 7E). Por lo tanto, dicha designación de paratipos, holotipo y alotipo es inválida, puesto que en ningún lugar de la descripción original se seleccionó explícitamente un ejemplar como holotipo. Así, es lógico suponer que alguien que tuvo acceso a la colección ordenó los ejemplares y los separó bajo un criterio nomenclaturalmente errado. Tomando en cuenta que los machos de basistrigata pueden ser confundidos fácilmente con machos de phyleus ssp. con base en su patrón alar y que ambas especies pueden ser recolectadas en el mismo lugar, designamos como lectotipo macho al ejemplar cuyo abdomen fue removido para la ilustración de la genitalia en la descripción original, que concuerda plenamente con la genitalia descrita e ilustrada en el presente trabajo. La única hembra de la serie típica presenta caracteres en el patrón alar diagnósticos de basistrigata, tal como detallado líneas arriba. La identidad taxonómica de los cinco sintipos machos restantes se considera incierta hasta que sus caracteres genitálicos y/o moleculares puedan ser examinados en detalle.

Figura 7 Colección de Insectos de la Universidad de Cornell (CUIC). (A) Caja entomológica conteniendo ejemplares de Hylephila, recuadro verde indicando el separador con sintipos de Hylephila basistrigata, recuadro azul esquina derecha inferior detalle de etiqueta roja del primer separador indicando ubicación de sintipos de H. basistrigata; (B) Caja entomológica de la colección de tipos N° 1504, recuadro rojo indicando el separador con sintipos de H. basistrigata; (C) vista en detalle del recuadro verde en la figura A, cinco sintipos machos de H. basistrigata, en circulo verde ejemplar macho con abdomen removido designado aquí como el lectotipo; (D) vista en detalle del recuadro rojo en la figura B, sintipos macho y hembra de H. basistrigata, etiquetados en la colección como holotipo y alotipo, respectivamente; (E) ejemplares ilustrados por Eaton (1932) en la descripción original de H. basistrigata; (F) ejemplar macho etiquetado en CUIC como “holotipo” de H. basistrigata, vista dorsal, vista ventral y etiquetas del ejemplar, respectivamente.
Discusión
La decisión de elevar a H. basistrigata al rango de especie está basada en los datos mostrados en este estudio, considerando en primer lugar la distancia genética, mayor del 3%, hallada entre poblaciones de basistrigata y phyleus, utilizando el gen COI, y considerando que Hebert et al. (2004) propusieron una distancia genética mínima del 3% como un criterio razonable para discriminar entidades biológicas en el rango de especie en Lepidoptera. En segundo lugar, los caracteres morfológicos observados en las estructuras genitálicas presentan diferencias notorias tanto en machos como en hembras lo cual refuerza la propuesta de restituir el estatus taxonómico H. basistrigata. Por otro lado, el presente trabajo es el primero que ilustra la genitalia femenina de basistrigata, puesto que Eaton (1932) sólo ilustró la genitalia masculina en la descripción original.
La planta hospedera de basistrigata, Distichlis spicata, es un pasto que se distribuye desde Norteamérica hasta el extremo sur de América del Sur (León et al. 1998, Lonard et al. 2013), siendo utilizada en algunos lugares como planta decorativa (Oakes 1990) y se caracteriza por crecer en zonas extremadamente salinas, donde otras especies halófilas no pueden desarrollarse (Pessarakli & Marcum 2013). Despland (2014) menciona que las larvas de Hylephila isonira se alimentan de Distichlis sp. en el norte de Chile, por lo que basistrigata no sería la única especie que utiliza este tipo de pasto. Por otro lado, H. phyleus en su rango de distribución utiliza distintas especies como plantas hospederas (Beccaloni et al. 2008, Robinson et al. 2010), siendo el pasto Cynodon dactylon el más comúnmente registrado en Arequipa como su planta hospedera.
Otro aporte importante en este trabajo es el registro por primera vez del comportamiento de oviposición de basistrigata, donde la hembra expulsa el huevo directamente a la base del pasto sin adherirlo a ningún sustrato, comportamiento no reportado antes en otro género de Hesperiidae; sin embargo, éste ha sido observado en mariposas paleárticas, principalmente de la subfamilia Satyrinae (Nymphalidae) (Wiklund 1984). Una hipótesis que explica este comportamiento en mariposas aduce la abundancia de la planta hospedera, pues si ésta es superabundante, cubriendo en algunos casos por completo el suelo, la hembra tiende a ser menos selectiva en el lugar donde oviposita (García-Barros & Fartmann 2009). Esto parece cumplirse para basistrigata, pues la planta hospedera D. spicata es muy abundante en la localidad de Yumina (Arequipa), siendo dominate en grandes extensiones de suelo, lo cual también se observa en otras zonas de la costa peruana conocidas en general como “gramadales”.
Hylephila basistrigata ha sido registrada como subespecie de phyleus en diferentes localidades desde Ecuador hasta Chile (Evans 1955, Ureta 1963, Lamas 1977, Benyamini et al. 2014, Farfán 2018), por lo que es posible que exista una superposición con la verdadera identidad de estos registros, ya que la forma más efectiva de identificación es a través de la revisión de la genitalia, y teniendo en cuenta que ambas especies utilizan las mismas áreas de vuelo en las mismas temporadas, no hay certeza de que todos esos registros realmente correspondan a basistrigata. Consideramos que los ejemplares de dichos registros deben ser examinados críticamente, para confirmar la presencia de esta especie en tales localidades y particularmente en Ecuador, donde Evans (1955) menciona un ejemplar macho proveniente del río Napo, o Argentina, donde cita un ejemplar de Santiago del Estero (erróneamente atribuido a Chile). De manera similar, tanto Pallister (1956) como Hayward (1973) citan localidades poco fidedignas para “Hylephila basistrigata”, tanto en Perú al este de los Andes, como en Argentina (“extremo norte de Salta”).
Con los resultados obtenidos en el presente trabajo el grupo phyleus, dentro del género Hylephila, estaría compuesto por 15 taxones hasta el momento: H. ancora (Plötz), H. basistrigata, H. fasciolata (Blanchard), H. isonira isonira, H. isonira mima, H. signata (Blanchard), H. zapala Evans, H. phyleus anca, H. phyleus andina, H. phyleus eureka, H. phyleus muertovalle, H. phyleus monticola, H. phyleus phyleus, H. phyleus taxus, y H. phyleus ssp. Sin embargo, se recomienda realizar una revisión taxonómica exhaustiva de este grupo para conocer realmente cuántas especies alberga en su rango de distribución y cuáles serían sus relaciones evolutivas dentro del género, continuando de esta manera el trabajo de Don MacNeill (MacNeill & Herrera 1998, MacNeill 2002, Brown et al. 2006).

