











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The rivers of Andes western slopes of Peru are characterized by being independent, seasonable, short, and drain directly to the Pacific Ocean (Ortega & Hidalgo 2008). Most of the coast from Peru is a desert, except for the northern regions in Piura and Tumbes departments, which have tropical weather with marked rainy seasons. Contrary to most rivers in the Peruvian Pacific drainages, the Chira River (Piura department) has permanent flowing water. Also, the Chira River is the southernmost portion of North Andean Pacific Slopes-Rio Atrato ecoregion of the South America, according to the classifications of ecoregions for freshwater fishes (301, Abell et al. 2008, Reis et al. 2016).
The Pacific slope basins have relatively poor diversity in fishes but a high degree of endemism, making the northern slope richer than the southward (Ortega & Hidalgo 2008, Reis et al. 2016). The lack of knowledge on the distribution and taxonomy of ichthyofauna from coastal drainages from Peru is noteworthy. The Chira River is the second most diverse river of the Peruvian coast after the Tumbes River (Ortega et al. 2015). Studies on freshwater fishes from this basin are scarce; the first study was made by Fowler (1945) registering three species for the basin, followed by Lopez et al. (1982) who registered eight species. Furthermore, other research recorded 13 species (Bonastre 2008) and 20 species (Marchena 2013, Ortega et al. 2015). However, recent collections revealed the presence of new records for the basin.
Moreover, anthropogenic activities, like agriculture, artisanal mining, pollution, and domestic wastewater discharge, have affected the preservation of the aquatic systems and biodiversity of the basin (More 2019, Ortega 2012). The aim of this study is to (1) update the composition of freshwater fishes from the Chira basin and (2) provide the altitudinal distribution of the species along the basin to better understand the ichthyofauna.
Material and methods
Study area. The Chira River is in Piura department in north western Peru. The Chira is considered part of the bi-national basin Catamayo-Chira between Ecuador and Peru. The headwaters of the Catamayo-Chira basin are in the complex lakes in the Pacific slope of Parque Nacional Podocarpus, Loja Province in Ecuador; and in the complex lakes Las Huaringas and Arrabiatadas, Ayabaca Province in Peru (SOFIA, 2018). The bi-national basin has a total surface drainage area of 19095 km2, where 11933 km2 (62%) are in Peru (MINAM 2012) which drain to the Pacific Ocean.
The area of the Chira River and its tributaries goes from 0 - 3600 m above sea level. and crosses the provinces of Ayabaca and Sullana, flowing from east to west. Their main tributaries in Peru are Macara, Quiroz, Chipillico and Alamor rivers, and Jaguay de Poechos, Venados and Saman streams (Fig. 1).
The distribution of species is strongly related to elevation and vegetation; therefore, we show the distribution of species following ecosystem classification in Peru. Along the Peruvian coast are eleven ecosystems and the Chira basin passes through seven of them; Coastal desert (Dc, 0 - 2500 m a.s.l.), Andean scrub (Ma, 1500 - 3900 m a.s.l.), Paramo (Pa, more than 3000 m a.s.l.), Seasonally dry hill and mountain forest (Bes-cm, 400 - 2000 m a.s.l.), Seasonally dry plain forest (Bes-ll, 0 - 500 m a.s.l), Seasonally dry riparian forest (Besr, 100 - 700 m a.s.l.), Western relic montane forest (Br-mvoc, 1400 - 3000 m a.s.l.) according to MINAM (2019).
Sampling. Fish samples from the Chira River belong to scientific collections and correspond to extensive sampling in October 2015 of 47 sites along the Chira basin and tributaries (Fig. 1, Appendix 1). Fishes were collected using seine nets, gill nets, cast nets and hand nets using the standardized collecting according to Birindelli et al. (2016).
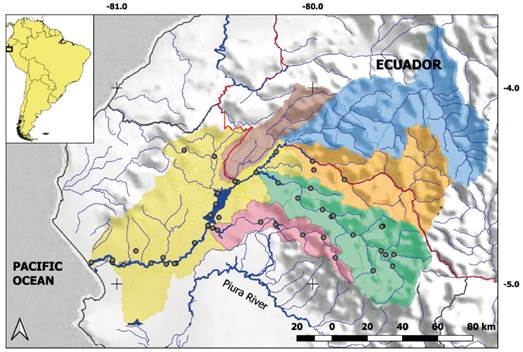
Figure 1 Location of sampling in Chira River basin, northwest Peru. Lower Chira (yellow), Alamor (brown), Chipillico (pink), Quiroz (green), Macará (orange), Catamayo (blue), International border (red line).
The captured fishes were anesthetized in clove oil (Lucena et al. 2013), then were fixed in 10% formalin and after 48 hours were preserved in 70% ethanol. Species were identified by primarily literature and deposited at the fish collection from Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima (MUSM) and Royal Ontario Museum, Toronto (ROM).
List of fishes. The complete list of species is arranged by the classification of order and family following Eschmeyer Catalog (Fricke et al. 2020). Additionally, information of the habitats (freshwater, brackish, and marine), the origin of species (native or introduced), the categorization of IUCN, and references are provided.
Results
The compilation of data recorded 27 species for the Chira River basin, distributed in 10 orders, 19 families (Table 1, Appendix 2). The ichthyofauna was predominantly Characiformes and Siluriformes representing 44% of all species (Fig. 2) with six species in each order. The most diverse families were Characidae and Cichlidae with four species in each family (Fig. 3).
Five species are considered new records for the Chira River: Astroblepus rosei, Basilichthys semotilus, Trichomycterus piurae, Oncorhynchus mykiss and Poecilia reticulata. Trichomycterus sp. is probably a new species to science. Furthermore, six marine species were recorded, collected approximately 90 m from the mouth of the river. Among the species recorded, 20 species are native; one catfish species (Chinchaysuyoa ortegai) is endemic to the Chira River; and seven species are introduced (Table 1).
Related to the abundance, a total of 3896 specimens were captured during fieldwork (October 2015). However, Poeciliidae were the most abundant with 39% of all specimens represented by Poecilia reticulata and Gambusia affinis which are non-native species; followed by Characidae with 27% of total represented by Eretmobrycon peruanus, Landonia latidens and Rhoadsia altipinna; Bryconidae with 13% of total represented by Brycon atrocaudatus; and Cichlidae with 15% represented by the native species Andinoacara rivulatus and A. stalsbergi, and the introduced species Coptodon rendalli and Oreochromis niloticus. Remaining families have less than 10% abundance (Fig. 3).
Regarding the ecosystems present in the basin, 15 species were exclusively found in the seasonally dry forest of the plain, eight species were found in both seasonally dry forest of the plain and seasonally dry forest of hill and mountain; Astroblepus rosei, Trichomycterus sp. and Basilichthys semotilus are exclusive from the latest. In Andean scrub ecoregion, Oncorhynchus mykiss is exclusive and Astroblepus rosei was found in both, Andean scrub and seasonally dry forest of hill and mountain (Fig. 4, Fig. 5).
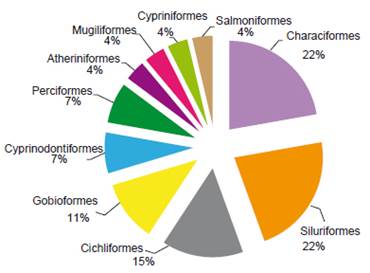
Figure 2 Species richness (percentages of total species number) according to fish orders sampled in Chira River basin.
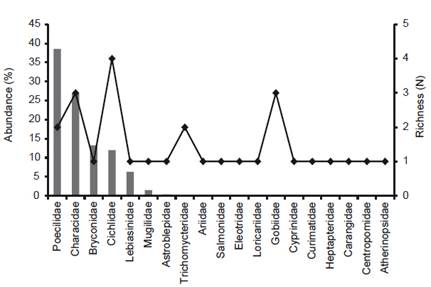
Figure 3 Species richness (N = expressed in species number) and abundance (percentages of total individuals), according to fish families sampled in Chira River basin.
Table 1 List of fishes in the Chira River basin. F = Freshwater; B = Brackish; M = Marine; N = Native; E = Endemic; I = Introduced; IUCN = Conservation status (IUCN 2020). * = New record
| ORDER / Family / Species | Habitat | Origin | IUCN | MUSM / References |
|---|---|---|---|---|
| CHARACIFORMES | ||||
| Bryconidae | ||||
| Brycon atrocaudatus (Kner 1863) | F | N | - | 48207, 48197, 48198, 52777 |
| Characidae | ||||
| Eretmobrycon peruanus (Müller & Troschel 1845) | F | N | - | 48187, 48195, 48199, 48219 |
| Landonia latidens Eigenmann & Henn 1914 | F | N | - | Meza-Vargas et al., 2019 |
| Rhoadsia altipinna Fowler 1911 | F | N | - | 48191, 48215, 48218, 52753 |
| Curimatidae | ||||
| Pseudocurimata troschelii (Günther 1860) | F | N | - | 48229 |
| Lebiasinidae | ||||
| Lebiasina bimaculata Valenciennes 1847 | F | N | - | 48194, 52754, 52758, 52762 |
| SILURIFORMES | ||||
| Trichomycteridae | ||||
| Trichomycterus piurae (Eigenmann 1922) * | F | N | - | 52816, 52817 |
| Trichomycterus sp. | F | N | - | 52759 |
| Astroblepidae | ||||
| Astroblepus roseiEigenmann 1922 * | F | N | DD | 52747, 52755, 52756, 52760 |
| Heptapteridae | ||||
| Pimelodella yuncensis Steindachner 1902 | F | N | - | 52780, 52797, 52808, 52810 |
| Loricariidae | ||||
| Hypostomus sp. | F | I | - | 48194 |
| Ariidae | ||||
| Chinchaysuyoa ortegai Marceniuk, Marchena, Oliveira & Betancur 2019 | F | E | - | Marceniuk et al., 2019 |
| SALMONIFORMES | ||||
| Salmonidae | ||||
| Oncorhynchus mykiss (Walbaum 1792) | F | I | - | |
| CYPRINODONTIFORMES | ||||
| Poeciliidae | ||||
| Gambusia affinis (Baird & Girard 1853) | F | I | LC | 48232,48243, 48246, 48250 |
| Poecilia reticulata Peters 1859 * | I | - | 23125 | |
| CICHLIFORMES | ||||
| Cichlidae | ||||
| Andinoacara rivulatus (Günther 1860) | F | N | 23127, 48210 | |
| Andinoacara stalsbergi Musilová, Schindler & Staeck 2009 | F | N | 48190, 48217, 48226, 48234 | |
| Coptodon rendalli (Boulenger 1897) * | I | LC | 48245 | |
| Oreochromis niloticus (Linnaeus 1758) | F | I | LC | 23124, 48189, 48192, 48204 |
| PERCIFORMES | ||||
| Carangidae | ||||
| Caranx caninus Günther 1867 | F, B, M | N | LC | 48225 |
| Centropomidae | ||||
| Centropomus unionensis Bocourt 1868 | B, M | N | LC | 48255 |
| GOBIIFORMES | ||||
| Eleotridae | ||||
| Gobiomorus maculatus (Günther 1859) | F, B, M | N | LC | 48239, 48248, 48331, 52812 |
| Gobiidae | ||||
| Awaous transandeanus(Günther 1861) | F, B, M | N | - | 48249 |
| Ctenogobius sagittula (Günther 1862) | F, B, M | N | LC | 48256 |
| ATHERINIFORMES | ||||
| Atherinopsidae | ||||
| Basilichthys semotilus (Cope 1874) * | F | N | LC | 52750 |
| MUGILIFORMES | ||||
| Mugilidae | ||||
| Mugil cephalus Linnaeus 1758 | F, B, M | N | LC | 48216, 48223, 48238, 48244 |
| CYPRINIFORMES | ||||
| Cyprinidae | ||||
| Cyprinus carpio Linnaeus 1758 | F, B | I | VU | Ortega et al., 2015 |
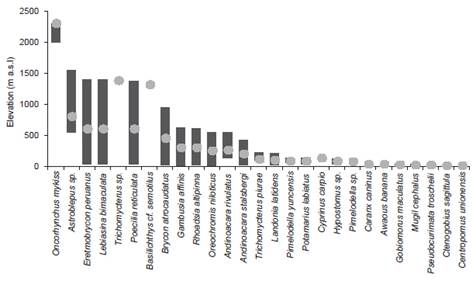

Figure 5 Photos of some species from the Chira River basin. (A) Brycon atrocaudatus, ROM 109159. (B) Eretmobrycon peruanus, ROM 109172. (C) Lebiasina bimaculata, ROM 109170. (D) Landonia latidens, MUSM 52785. (E) Rhoadsia altipinna, ROM 109160. (F) Trichomycterus piurae, MUSM 52816. (G) Trichomycterus sp., MUSM 52759. (H) Astroblepus rosei, MUSM 52747. (I) Pimelodella yuncensis, MUSM 52780. (J) Gambusia affinis, MUSM 52781, male. (K) Andinoacara stalsbergi, MUSM 52782. (L) Gobiomorus maculatus, MUSM 52812. (M) Basilichthys semotilus, MUSM 52750. Scale = 1 cm.
In the Chira River 11 species have been categorized according to IUCN (2020) conservation status, most of them euryhaline (occurs in freshwater, brackish, and marine habitats) and four introduced species (Table 1). Among native species, six are Least Concern (LC) and one species is in the Deficient Data (DD) category.
Discussion
The Chira River basin has characteristics of ecoregion and ecosystem that make it unique and because that is considered an endemic area (Albert et al. 2011, Reis et al. 2016). Although the Chira river basin is unique in comparison to other Peruvian river basins, little research has been done to demonstrate the diversity of the area. This survey represents the most thorough sampling of the Chira basin and most complete sampling on the Peruvian coast.
Although the river is considered an ecosystem (MINAM, 2019), it is not uniform from headwater to mouth in terms of fishes. There has been observed differential distribution of fishes along the Chira basin, which are related to ecosystems it crosses. As ecosystems are established by considering biodiversity and characteristics of habitats as elevation, vegetation, ecosystem services, weather, etc. (MINAM, 2019), future monitoring of biodiversity can be integrated with ecosystem assessments as well (Llambi et al. 2020).
Diversity of fishes in the basin was increased to 27 species, compared to previous ichthyofauna diversity studies in the basin and its tributaries (Bonastre 2008, Marchena 2013, Ortega et al. 2015). Most prolific species are the orders Characiformes and Siluriformes which belong to the superorder of Ostariophysi, predominant in most freshwater rivers of the Neotropical region (Roberts 1972, Lowe-McConnell 1987, Reis et al. 2003, 2016) and in coastal rivers from the northeast of Peru (Ortega et al. 2015). Besides, species show restricted distribution for altitude. For instance, most of them are distributed less than 500 m a.s.l, Astroblepus, Basilichthys and Trichomycterus are present in a range between 500-1500 m a.s.l. and the introduced species Oncorhynchus is found more than 2000 m a.s.l.
Among new additions there are three genera (Chinchaysuyoa, Astroblepus and Basilichthys), of which the catfish Chinchaysuyoa ortegai is a new genus, and species, endemic from the Chira River (Marceniuk et al. 2019). Additionally, the presence of Astroblepus rosei Eigenmann (1922), originally described from Jequetepeque River, 300 km south of Chira River, represents an extension in its distribution. The current distribution of Basilichthys semotilus is from Reque River, Lambayeque department to Sama River, Tacna department (Dyer 2003); however, the species was recorded in the Chira basin in the present study and by Bonastre (2008) in her unpublished undergraduate project (no voucher deposited in any fish collection), representing the northernmost distribution of B. semotilus.
Non-native fish species were introduced to several rivers in Peru as biological control vectors (Gambusia affinis and Poecilia reticulata), farming and sport fishing (Oncorhynchus mykiss), fish culture (Oreochromis niloticus), and living food to big fishes (Coptodon rendalli) (Ortega et al. 2007). Ortega et al. (2015) recorded Gambusia affinis and Oreochromis niloticus from Tumbes and Chira River which is the northernmost distribution for these species; in this study we added Poecilia reticulata, Oncorhynchus mykiss, and Coptodon rendalli (previously introduced as Tilapia rendalli) which represent the northernmost distribution for these species in the Pacific drainages. As non-native species have increased along the Chira River, the future impact that these exotic species could have on native species is of high importance. Some impacts on native species include competition, predation, and habitat alteration (Primack 1993, Simberloff 2000), which has already happened in some native species in the Amazonian drainages and Pacific coastal basins in Peru (Ortega et al. 2007). Additionally, Hypostomus sp. should be treated carefully, as the genus is well distributed along the Amazonian basin and not found in the Peruvian coast. This implies the species was transferred from the Peruvian amazon, or is a species related to Hypostomus species of the north coastal rivers from Ecuador; however, more taxonomic studies are needed. Also, Arapaima gigas (paiche) is the most recent species transferred to San Lorenzo reservoir for farming purposes to improve the economy of the residents of Las Lomas municipality (Produce 2018), but future studies could show us the impact of the introduction of this non-native species.
During fieldwork, anthropogenic activities were identified which have modified aquatic bodies, such as agriculture, mining, dams, pollution and even proliferation of exotic species which threaten native species (Ortega et al. 2007). Changes in environmental conditions can affect biodiversity and its distribution (Lujan et al. 2013, Albert et al. 2020); hence those anthropogenic activities are contributing to the modification of habitats. For instance, the lower Chira River, headwater of Chipillico and Queiroz River are highly impacted by agriculture (MINAM, 2019). Also, as was pointed out in Meza-Vargas et al. (2019), water diversion between Chira and Piura basins can explain the presence of Landonia latidens in the latest basin. We recommend increasing efforts for regulation of these activities for the sake of keeping in good conditions and conservation of the basin and the region. Finally, most of the species in the Chira basin lack categorization according to IUCN; therefore, we recommend categorizing mainly native species from the Chira River to avoid loss of fish diversity along the coastal rivers of Peru.

