Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versão impressa ISSN 1609-9117
Rev. investig. vet. Perú v.17 n.1 Lima jan./jun 2006
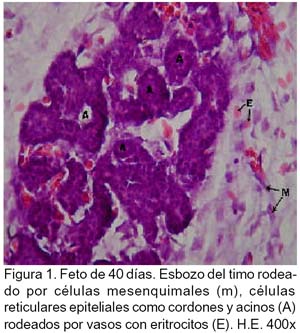
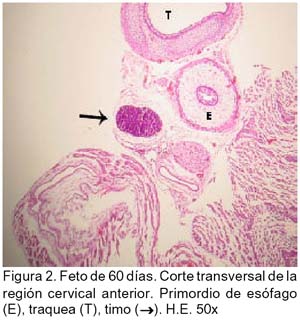
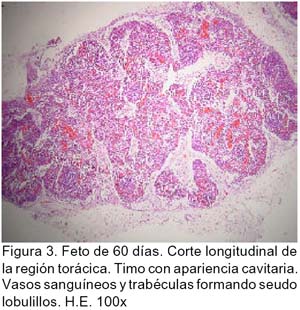
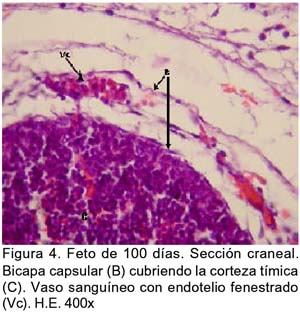
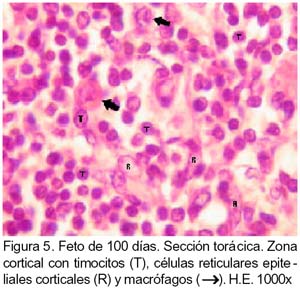
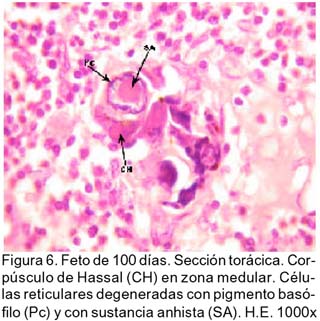
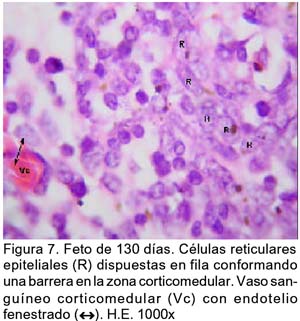
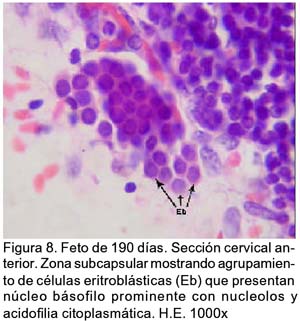
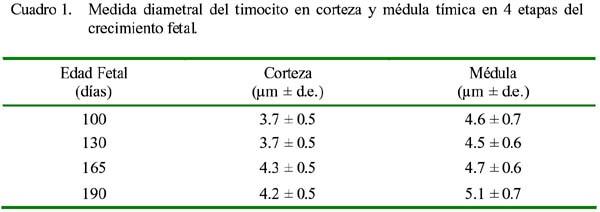
ARTÍCULO PRIMARIO Caracterización histológica del timo en fetos de alpaca Juan Montenegro B.1; Nieves Sandoval Ch.2,3; Alfonso Chavera C.2 y Alberto Manchego S.4 1 Práctica privada. E-mail: jubevet76@walla.com RESUMEN El objetivo del presente trabajo fue el estudio citoarquitectural de los segmentos cervicales y torácicos del timo en fetos de alpaca (Lama pacos). Se recolectaron 10 fetos entre 40 y 190 días de desarrollo para ser sometidos a estudio histológico. A los 40 días, el esbozo del timo tiene una morfología citoarquitectural de tipo acinar infiltrado por escasas células linfocíticas y rodeado por células mesenquimales. A los 60 días, el segmento cervical se presenta como un folículo linfoide densamente poblado por timocitos, mientras que la región torácica tiene una apariencia cavitaria con proyecciones trabeculares cortas. La diferenciación de corteza y médula se inicia en los segmentos cervicales posteriores y torácicos a los 100 días y en los segmentos cervicales anteriores a los 130 días. Todos los segmentos presentan disposición lobulillar bien definidos en corteza y médula a los 165 días. La cápsula tímica se mantiene principalmente como una doble capa celular con tejido conjuntivo laxo. Los vasos sanguíneos subcapsulares y trabeculares presentan endotelio fenestrado. Las células eritroblásticas se ubican en las zonas subcapsulares y corticales periféricas desde los 100 días. La barrera hematotímica comienza a organizarse desde los 60 días. Los corpúsculos medulares de Hassal son evidentes a los 100 días y la barrera celular reticular epitelial en la zona corticomedular se encuentra desde los 130 días. ABSTRACT The objective of the present study was to study the cytoarchitectural structure of the cervical and thoracic segments of thymus in alpaca (Lama pacos) fetuses. For this purpose, 10 fetuses between 40-190 days of gestation were collected form the local slaughterhouse. At day 40, the developing thymus has acinar cytoarchitectural morphology, infiltrated by a few lymphoid cells surrounded by mesenchymal cells. At day 60, the cervical segment looks like a «follicular» lymphoid, densely populated by thymocytes, whereas the thoracic region has a cystic appearance with short trabecular projections. Cortex and medulla differentiation starts in the posterior cervical and thoracic segments by day 100 and in the anterior cervical segments by day 130, and is well established by day 165. The thymic capsule remained as a double cellular layer of lax connective tissue. Subcapsular and trabecular blood vessels have fenestrated endothelium. Erythroblastic cells were observed by day 100 in the cortex and subcapsule zones. Medullar Hassalls corpuscles were observed by day 100, and the epithelial reticular cell barrier in the cortical medullar zone was observed by day 130. INTRODUCCIÓN La alpaca (Lama pacos), especie nativa y de gran potencial económico en el Perú, es susceptible a múltiples agentes patógenos, especialmente en hatos con inadecuados niveles tecnológicos de crianza. La respuesta frente a un agente patógeno es dado por el sistema inmunológico, siendo el timo un órgano inmune primario que cumple un rol central en el sistema inmune. Allí, las células linfoides de la línea T puedan desarrollar, proliferar, madurar, generar su repertorio de receptores de antígenos, y dejar este órgano para poblar órganos linfoides secundarios (Romano et al., 1999). La estructura histológica del timo y la ontogenia del sistema inmune ha sido estudiada y establecida en la mayoría de especies domésticas (Osburn et al., 1982; Tizard, 2002); sin embargo, no se dispone de estudios en fetos de camélidos sudamericanos. El objetivo del presente estudio fue caracterizar histológicamente el desarrollo del timo en la etapa fetal de la alpaca, estableciendo los tipos celulares que lo conforman durante su desarrollo estructural e infiriendo la probable función inmunológica que pueda efectuar en la etapa fetal. MATERIALES Y MÉTODOS Se emplearon 10 fetos de alpacas entre 40 y 190 días de edad gestacional, provenientes de hembras de descarte que fueron sacrificadas en un camal de la ciudad de Huancayo, Junín. La zona se encuentra a 3,350 msnm. La edad gestacional fue determinada utilizando como referencia el diámetro biparietal fetal (Gazitua et al., 2001), medido a través de un caliper Pretul de 5. Los fetos fueron extraídos de las bolsas amnióticas y fijados de inmediato en formol bufferado al 10%. La obtención del tejido tímico se realizó en base a los siguientes criterios: - En fetos mayores de 4 meses el timo, se diseccionó in situ, obteniéndose secciones de tejido de la porción cervical y torácica. - En fetos menores de 4 meses, se hicieron cortes o secciones transversales a lo largo del cuello y entrada del tórax. La descripción histológica del desarrollo embriológico del timo se hizo en seis grupos etáreos: 40, 60, 100, 130, 165 y 190 días de edad fetal aproximada (los últimos 4 grupos estuvieron compuestos por 2 fetos). Las muestras se sometieron a técnicas histológicas de rutina y se emplearon 4 tinciones: Hematoxilina Eosina para la distribución de parénquima y estroma, Masson para la distribución de las fibras colágenas, Wilder para la distribución de las fibras reticulares, y Jonson para observar los gránulos de los mastocitos. RESULTADOS El timo, a los 40 días de desarrollo embriológico, se presenta como una organización de células reticulares epiteliales poliédricas de núcleos basófilos a manera de cordones y acinos, tomando una apariencia glandular (Fig. 1). Se observan células esféricas con núcleos pequeños y escaso citoplasma, compatibles con células linfoides (timocitos), dispuestas dentro y entre los cordones o rosetas a nivel del timo torácico. A la edad de 60 días, el timo se observa como una estructura folicular densamente poblada por timocitos en los segmentos cervicales (Fig. 2), mientras que se muestra una organización seudolobulillar indiferenciada, aparentando un órgano cavitario en el segmento torácico (Fig. 3). Dentro de éstas cavidades tímicas no existe organización celular definida, encontrándose los timocitos y las células reticulares epiteliales dispuestos aleatoriamente, así como múltiples tipos celulares inmaduros de la línea mieloblástica, además de abundantes vasos sanguíneos tortuosos y pletorizados. A la edad de 100 días, la estructura folicular de los segmentos tímicos cervicales muestra una tenue diferenciación medular. Están cubiertos por una cápsula compuesta por doble capa de tejido conjuntivo laxo y dos capas celulares de fibroblastos (Fig. 4). La estructura seudolobulillar del segmento torácico se encuentra diferenciada en corteza y médula. La corteza está tapizada por células reticulares epiteliales limitantes y en la zona medular se aprecian macrófagos (Fig. 5) y corpúsculos de Hassal (Fig. 6). Estos son cuerpos acidófilos que constituyen células reticulares epiteliales aplanadas y frecuentemente hialinizadas, dispuestas alrededor de una zona central constituída por una o varias células queratinizadas (pigmentos basófilos) y/o degeneradas. En las zonas trabeculares y subcapsulares se encontraron algunos grupos de células eritroblásticas. Así mismo, la barrera hemato-tímica se encuentra definida. A la edad fetal de 130 días, tanto la estructura seudolobulillar de la sección cervical como la estructura lobulillar de la sección torácica están diferenciadas en corteza y médula. Las células reticulares epiteliales de la zona corticomedular se encuentran estrechamente relacionadas conformando una barrera celular que divide corteza y médula (Fig. 7). Los corpúsculos de Hassal de la médula son evidentes. En las zonas subcapsulares y trabeculares se observa un tipo celular granular (mastocito). Las secciones cervical y torácica presentan como una estructura lobulillar diferenciada en corteza y médula a los 165 días de desarrollo fetal. En las zonas subcapsulares y trabeculares se encuentran abundantes y variados tipos celulares de la línea mieloide y eritroblástica. La estructura de ambas secciones a los 190 días es similar a la observada a los 165 días, con la salvedad de que en algunas zonas de la sección torácica, la cápsula se hace más fina presentando una sola capa celular. En las áreas subcapsulares y trabeculares se encuentran, dentro los tipos celulares de la línea mieloide, grupos de células eritroblásticas (Fig. 8) y otras compatibles con células plasmáticas. La medida diametral de los timocitos medulares es mayor que de los corticales (Cuadro 1). Los timocitos medulares presentan un núcleo más grande y de mayor contenido citoplasmático. DISCUSIÓN El esbozo del timo en los fetos de alpacas se hizo visible a los 40 días de edad, similar a lo descrito en el feto bovino (Schultz et al., 1973), a pesar que las alpacas tienen un periodo de gestación más largo (11 meses). Las formaciones celulares a manera de acinos glandulares, cordones o rosetas fue similar a lo descrito para animales domésticos (Jubb et al., 1993), donde el componente epitelial reticular es desplazado caudalmente en el cuello e invade el intersticio mediastínico con yemas tipo roseta irregulares y sólidas que crecen para formar los lóbulos primordiales. Los linfocitos (timocitos) que se observan a los 40 días en la alpaca son igualmente observados en el timo fetal bovino (Osburn et al., 1982) y ovino (Jubb et al., 1993) en edades similares. Se presume que éstas células son las primeras células pro timocitos (linfoblastos) que están siendo reclutadas desde órganos hematopoyéticos fetales hacia el timo primitivo para poblarlo, comenzar el proceso de maduración tímico y dar origen a los primeros linfocitos T. El desarrollo temprano del sistema lobular primitivo, donde las células reticulares epiteliales que forman las yemas tipo rosetas se estratifican tomando formas sólidas inicialmente y luego se hacen císticas como, al parecer, sucede en el timo fetal de alpaca, ha sido descrito por Jubb et al. (1993). La invasión vascular se presentó previo a la formación de lóbulos con componentes medulares y corticales, tal como ha sido reportado en la literatura (Banks, 1992). La cápsula tímica, a partir de los 100 días, está conformada por una capa externa que cubre la periferia del lóbulo tímico y una capa interna que delimita el contorno periférico de la corteza del lóbulo (Sainte-Marie et al., 2005), similar a lo encontrado en el presente estudio. Además, en el timo fetal de la alpaca se encuentran abundantes vasos sanguíneos subcapsulares pletorizados de endotelio plano fenestrado, así como células de la línea blanca y roja. En la rata neonatal ocurre un patrón similar, donde los leucocitos se encuentran próximos a los capilares subcapsulares y participan en acontecimientos tímicos peculiares, tales como el reclutamiento de células precursoras sanguíneas (Sainte-Marie et al., 2005). Diferencias morfológicas entre timocitos, donde los corticales fueron más pequeños que los medulares, se describen en otras especies como los humanos (Junquera et al., 1988) y la brema del mar (Romano et al., 1999). Los macrófagos que se observan desde los 60 días en los fetos de alpaca cumplirían la función de remoción de los cuerpos apoptósicos, aunque no se pudo determinar estos cuerpos. En el timo fetal se pudo apreciar la unión corticomedular como una barrera de células reticulares epiteliales dispuestas secuencialmente próximas entre sí. Allí, según Abbas et al. (2002), ocurrirían interacciones físicas con las células reticulares epiteliales durante la migración de los timocitos a través de la unión corticomedular. La porción torácica del timo se organiza a partir de la edad fetal de 100 días en lobulillos con corteza definida que comparten una médula común, donde la presencia del corpúsculo medular de Hassal es evidente. Estos corpúsculos aparecen más tardíamente que en el feto bovino donde ocurre a los 65 días (Schultz et al., 1973). Tales corpúsculos se observaron de manera similar a lo descrito por Banks (1992) y Dellmann y Brown (1980). En el timo fetal de alpaca se aprecian grupos celulares hematopoyéticos indiferen-ciados, formando nidos compatibles con células eritroblásticas. Tienen núcleo esférico basófilo intenso, con presencia de nucleolos y citoplasma acidófilo, y una proporción de núcleo a citoplasma de 6:1. Estas células se ubican en las cavidades del timo inmaduro de 60 días, pero a partir de que el timo se organiza, sus cavidades se pueblan de timocitos, definiéndose la corteza y médula. Las células se ubican principalmente en las áreas trabeculares, subcapsulares, y ocasionalmente en la corteza periférica. Vicente et al. (1998) reportan que el parénquima cortical del timo de rata hospeda una eritropoyesis transitoria alrededor del nacimiento. Esto indicaría que el timo fetal de alpaca también estaría cumpliendo alguna función eritropoyética aislada. CONCLUSIONES El desarrollo citoarquitectural del timo fetal de alpaca sigue un patrón similar al de otras especies de rumiantes domésticos. El rudimento tímico está presente a la edad de 40 días, definiéndose en corteza y médula a la edad de 100 días en los segmentos cervicales posteriores y torácicos y a los 130 días de edad en los demás segmentos. Los corpúsculos de Hassal se encuentran en la médula tímica a partir de los 100 días de edad. El timo fetal de alpaca alberga algunos nidos celulares eritroblásticos. LITERATURA CITADA 1. Abbas, A.K.; A.H. Lichtman; J.S. Pober. 2002. Inmunología celular y molecular. 4ta ed. p 129-165. McGraw-Hill Interamericana. Madrid. [ Links ] 2. Banks, J. 1992. Histología veterinaria aplicada. p 399-415. Ed. Manual Moderno. México DF. [ Links ] 3. Dellmann, H.D.; E.M. Brown. 1980. Histología veterinaria. p 177-198. Ed. Acribia. Zaragoza. [ Links ] 4. Gazitua, F.J.; P. Corradini; G. Ferrando; L.A. Raggi; V.H. Parraguez. 2001. Prediction of gestational age by ultrasonic fetometry in llamas (Lama glama) and alpacas (Lama pacos). Anim. Reprod. Sci. 66: 81-92. [ Links ] 5. Jubb, K.V.F.; P.C. Kennedy; N. Palmer. 1993. Pathology of domestic animals. 4th ed. Vol. 3. p 209-211. Academic Press. California, USA. [ Links ] 6. Junqueira, L.C.; J. Carneiro. 1988. Histología básica. 3ra ed. p 285-306. Ed. Salvat. Barcelona. [ Links ] 7. Osburn, B.I.; N.J. MacLachlan; T.G. Terrel. 1982. Ontogeny of the immune system. J.A.V.M.A. 181: 1049-1051. [ Links ] 8. Romano, N.; M. Fanelli; G. Maria del Papa; G. Scapigliati; L. Mastrolia. 1999. Histological and cytological studies on the developing thymus of sharpsnout seabream, Diplodus puntazzo. J. Anat. 194: 39-50. [ Links ] 9. Sainte-Marie, G.; F.S. Peng; D. Marcoux. 2005. The stroma of the thymus of the rat: morphology and antigen diffusion, a reconsideration. Am. J. Anat. 177: 333-352. [ Links ] 10. Schultz, R.D.; H.W. Dunne; C.E. Heist. 1973. Ontogeny of the bovine immune response. Infect. Immun. 7: 981-991. [ Links ] 11. Tizard, I.R. 2002. Inmunología veterinaria. 6ta ed. p 74-89, 227-239, 470-484. McGraw-Hill Interamericana. México DF. [ Links ] 12. Vicente, A.; A. Varas; R. Sacedón; A.G. Zapata. 1998. Histogenesis of the epithelial component of rat thymus: an ultrastructural and immunohistological analysis. Anat. Rec. 244: 506-519. [ Links ]
2 Laboratorio de Histología, Embriología y Patología, FMV–UNMSM
3 E-mail: nieves_1@mixmail.com
4 Laboratorio de Microbiología y Parasitologìa Veterinaria, FMV-UNMSM
Palabras clave: alpaca, feto, timo, timocito, corpúsculo de Hassal
Key words: alpaca, fetus, thymus, Hassal corpuscle