










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versão impressa ISSN 1609-9117
Rev. investig. vet. Perú vol.27 no.4 Lima out./Dez. 2016
http://dx.doi.org/10.15381/rivep.v27i4.12642
ARTÍCULOS PRIMARIOS
Análisis de los Cambios Etarios según Sexo en el Borrego de Chiapas
Age changes in Chiapas sheep breed according to sex
P.M. Parés-Casanova1,2, M. Caballero-Sala1, R. Perezgrovas2
1 Departament de Ciència Animal, Universitat de Lleida, Lleida, Cataluña, España
2 Laboratorio de Calidad de Lana, Instituto de Estudios Indígenas, Universidad Autónoma de Chiapas, San Cristóbal de Las Casas, Chiapas, México
3 E-mail: peremiquelp@ca.udl.cat
RESUMEN
El objetivo del estudio fue la evaluación de los cambios que se producen en diversas medidas corporales en el Borrego Chiapas. Se utilizaron 66 machos y 79 hembras de diversas edades y se registraron las siguientes variables: peso vivo, perímetro torácico, perímetro de la rodilla, perímetro de la caña, perímetro de la corona, perímetro de la rabadilla y longitud de la oreja. El estudio reflejó que hay cambio con la edad diferencial de distintos componentes del cuerpo, en ningún caso isométricos, siendo el peso vivo y el perímetro torácico las variables más discriminativas en el cambio con la edad. Aunque el peso vivo de los machos adultos (>12 meses, n=56) fue mayor que en las hembras (n=64) (U=1320, p=0.014), pero similar en el perímetro torácico (U=1720, p=0.704), las trayectorias de las rectas de regresión de la edad no presentaron diferencias de trayectoria alométrica entre machos y hembras, ni para el peso vivo (F=1.006, p=0.317) ni para el perímetro torácico (F=1.265, p=0.262).
Palabras clave: alometría, dimorfismo sexual, ontogenia, variación intraespecífica
ABSTRACT
A survey was conducted in order to study changes in body measurements in the Chiapas sheep breed. For this purpose 66 males and 79 females of various ages were measured: body weight, chest girth, knee perimeter, cannon perimeter, coronne perimeter, hock perimeter and ear length. The study reflects a regional change according to age, which are not isometrical. Body weight and chest girth are the most discriminative variables, which presented identical age regression trajectories for both sexes (F=1.006, p=0.317). Although body weight in adult male (>12 months, n=56) was greater than those of females (n=64) (U=1320, p=0.014), and no differences appeared for chest girth (U=1720, p=0.704), regression trajectories for both traits appeared similar.
Key words: allometry, sexual dimorphism, ontogeny, intraspecific variability
INTRODUCCIÓN
El término alometría se refiere al cambio de dimensión de una parte corporal en relación con el cambio de dimensión en el tamaño total. Durante el desarrollo de un organismo, la alometría puede referirse al cambio diferencial de diferentes partes del cuerpo con la edad.
Descrita por primera vez por Snell en 1892 y D’Arcy Thompson en 1917, la alometría es conocida por la expresión de Huxley (1972): y=a xb, donde «y» es el tamaño de una de las partes, «x» el tamaño de otra tomada como referencia, y «a» y «b» son constantes. Este modelo se basa en el principio de que la velocidad de cambio relativo entre dos variables se mantiene constante con la edad. Si «b» (exponente alométrico) es 1 cuando se relacionan longitudes con longitudes, o superficies con superficies, o bien volúmenes (masas) con volúmenes (masas), los cuerpos poseen similitud geométrica (o se dice que los cuerpos son isométricos). La ecuación ha sido utilizada ampliamente en numerosas disciplinas: fisiología, morfogénesis, evolución, etc. (Gould, 1966), y en toda clase de grupos animales, y es bien conocida, particularmente en biología, debido a las aplicaciones prácticas que tiene el conocer las tasas de cambio con la edad diferencial de cada parte del cuerpo. Los parámetros de este cambio resultan, además, una parte central de todo trabajo de investigación que se dirija a la explotación racional de los animales domésticos (Véase Pelábon et al [2013] para mayor información).
Cuando se consideran las diferentes partes o tejidos de un organismo, no todas crecen con la misma intensidad y ritmo, lo que origina un desarrollo diferencial. Por lo tanto, otro concepto íntimamente ligado al desarrollo es el de desarrollo relativo o alométrico. El principio de la alometría es que los cambios morfogenéticos que se producen en un animal en desarrollo tienen lugar, principalmente, por desarrollo relativo; o sea, por el desarrollo que se produce en las distintas partes del organismo animal como un todo. Ejemplo: los huesos del cráneo crecen con la misma tasa específica de desarrollo que el cerebro; los músculos que forman el muslo de un animal no pueden crecer en discordancia con los huesos que forman su base ósea.
El bor rego Chi apas está muy bi en adaptado a zonas montañosas, es un animal rústico y su crianza tienecomo principal objetivo laproducción delana. La ovinocultura indígena en la región montañosa de Los Altos de Chiapas difiere con la practicada en otras partes de México, pues se aleja de los patrones establecidos debido a que es desarrollada exclusivamentepor mujeres. Ellas se encar gan del cui dado de l os borr egos, trasquilan y tejen la lana para la confección de los vestidos tradicionales del grupo étnico Tzotzil; asimismo, toman las decisiones relacionadas con esta actividad que, además deproductiva, tieneun carácter sociocultural. Actualmente las cualidades lanígeras de la raza se aprovechan en sistemas de producción extensiva bajo pastoreo, pero se requiere contar con mayor información sobre el crecimiento y desarrollo en estas condiciones. Este trabajo, pues, viene a representar esta contribución.
Un aspecto importante en este estudio es la diferenciación entre crecimiento y desarrollo, que no debe confundirse. Crecimiento y desarrollo son fenómenos separados, si bien se puede plantear alguna dificultad al definirlos. Según Hammond (1960), se entiende por ‘crecimiento’ el aumento de masa (peso) experimentado por los animales desde el nacimiento hasta su estabilización en la edad adulta, expresado por unidad de tiempo, y por desarrollo, las modificaciones que experimentan las proporciones, conformación, composición química corporal y funciones fisiológicas del animal a medida que avanza la edad. Aunque ambos fenómenos pueden producirse simultáneamente, es posible que un individuo se desarrolle (aumente su largo y alto, o sea con desarrollo) sin experimentar alteraciones en su peso (sin crecimiento) o que un individuo adulto (que ha terminado su desarrollo) aumente su peso por engorde (con crecimiento).
MATERIALES Y MÉTODOS
Se utilizaron 145 ejemplares (66 machos y 79 hembras) de un amplio espectro de edad (3 a 156 meses, siendo para machos y hembras el mismo rango), procedentes de varias comunidades indígenas de la zona de Chiapas (oeste de México). De cada animal se obtuvieron las siguientes variables: peso vivo, perímetro torácico, perímetro de la rodilla, perímetro de la caña, perímetro de la corona, perímetro de la rabadilla y longitud de la oreja, siguiendo los protocolos habituales en zoometría (Rodero y González, 2009). El peso vivo se registró mediante el pesado directo de los animales con balanza digital. Todos los parámetros fueron obtenidos y anotados siempre por la misma persona.
Para detectar alometría se utilizó, como medida de referencia (variable independiente), la edad expresada en meses. La edad fue la que declararon las propias ganaderas.
Análisis Estadístico
El test D de Kolmogorov-Smirnov evaluó la igualdad en la distribución de las muestras. Para evaluar las trayectorias alométricas de machos y hembras, se calcularon las regresiones bi y multivariantes de los logaritmos de cada una de las medidas con respecto a la edad, también transformada logarítmicamente, mediante un análisis univariado de la covarianza (ANCOVA), y entre sexos, separadamente.
La expresión alométrica en este caso fue y=b log x+log a. Se utilizó como variables dependientes para el análisis de varianza los scores de los Componentes Principales (CP) que resumiesen al menos el 80% de la variación en forma (Anzelmo et al., 2012). Los CP se obtuvieron del Análisis de Componentes Principales (ACP) a partir de la matriz de varianza/covarianza. Finalmente, la comparación de medianas del peso vivo y del perímetro torácico se realizó con el test no paramétrico U de Mann-Whitney. Todos los análisis estadísticos fueron realizados mediante la aplicación estadística PAST v. 2.17c (Hammer et al., 2010) a un valor p del 5%.
RESULTADOS
La distribución etaria para ambos sexos resultó sin diferencias significativas (D=0.105, p=0.794). Los resultados de las regresiones bivariantes (Cuadro 1) indicaron la existencia de una regresión significativa y positiva en casi todas las dimensiones, excepto para la longitud de la oreja (p=0.056) en el caso de los machos. Resulta llamativa la escasa pendiente de la recta de regresión que presentaron las variables (valor a), lo que indica que dichos parámetros alcanzan muy lentamente el desarrollo definitivo.
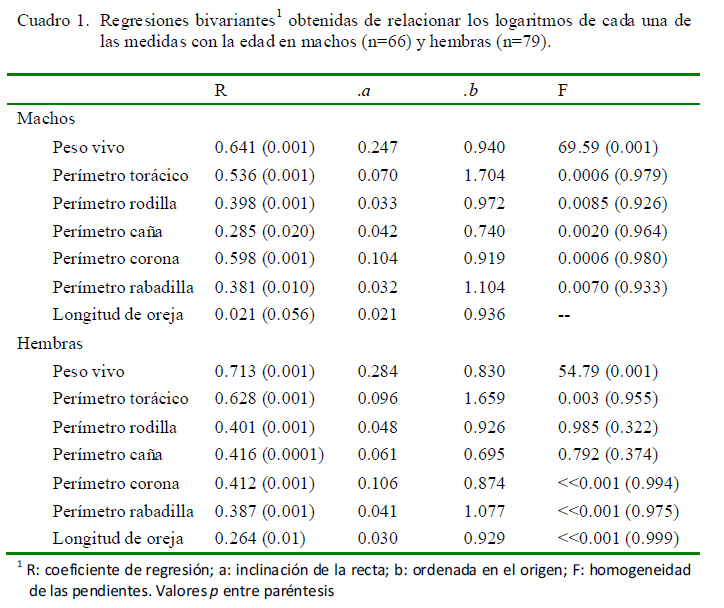
En el ACP, el CP1 fue el único que presentó un vector propio (eingenvalue) superior al corte de Jolliffe (12.62), y por ende significativo, explicó más del 90% de la variación, tanto en machos como en hembras (Cuadro 2). Para ambos sexos, el análisis de regresión multivariado demostró que el CP1 representa cambios, sobre todo en tamaño (peso vivo y perímetro torácico), asociados a los cambios corporales relacionados con la edad (Cuadro 3).
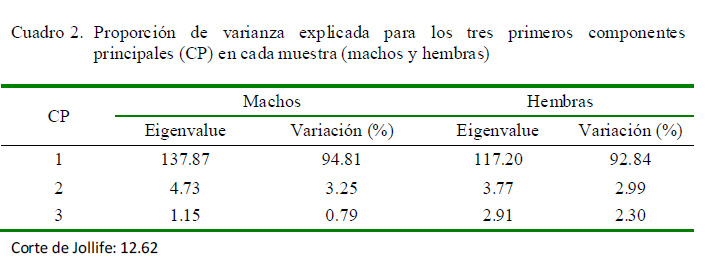

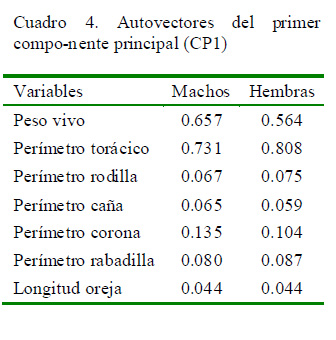
El Cuadro 4 muestra la contribución de cada una de las variables para el CP1, siendo todas positivas. Para este CP, las variables con mayor contribución para ambos sexos fueron el peso vivo y el perímetro torácico (0.657 y 0.564, y 0.731 y 0.808 en machos y hembras, respectivamente).
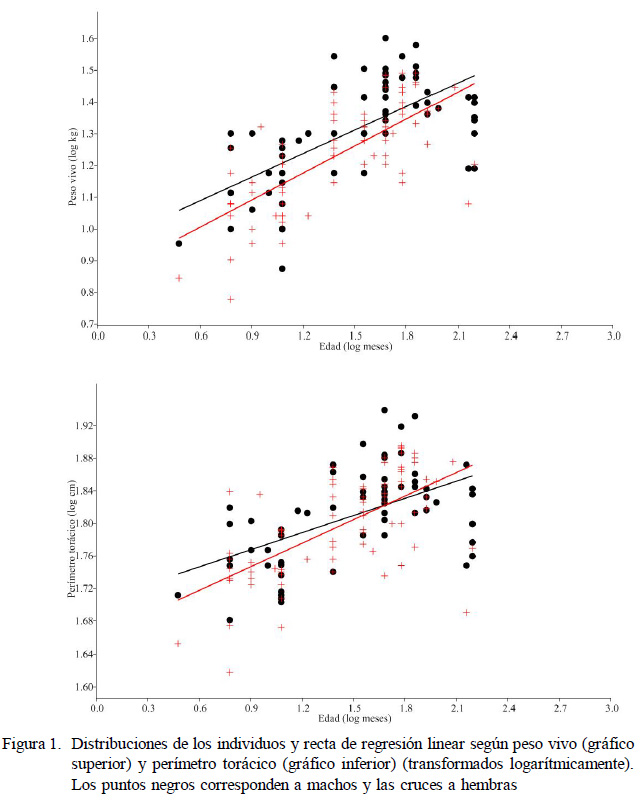
El peso vivo de los machos adultos (>12 meses, n=56) fue mayor que en las hembras (n=64) (U=1320, p=0.014); sin embargo, el perímetro torácico resultó similar para ambos sexos (U=1720, p=0.704). El peso vivo inmaduro (<12 meses) de los machos, en comparación con las hembras, no apareció significativamente diferente (U=49.5, p=0.163). Considerando las rectas de regresión de la edad, se observó que las trayectorias no presentaban diferencias de trayectoria alométrica entre machos y hembras ni para el peso vivo (F=1.006, p=0.317) ni para el perímetro torácico (F=1.265, p=0.262) (Figura 1). Si los pesos iniciales (entre 3 y 12 meses) son iguales para ambos sexos y la trayectoria de crecimiento es similar, puede deducirse que los machos presentan una mayor velocidad en la ganancia de peso con la edad.
DISCUSIÓN
La modificación de las proporciones de las diversas regiones morfológicas, órganos y sistemas, hasta que se accede al estado adulto o estable, es el resultado del crecimiento desigual o relativo de las diferentes partes del organismo y, para su cuantificación, el modelo alométrico es el más idóneo. Este modelo se basa en el principio de que la velocidad de crecimiento relativa entre dos variables que crecen se mantiene constante. Ello permite relacionar ambas variables mediante una ecuación sencilla de tipo potencial, que mediante una transformación logarítmica se convierte en una recta cuya pendiente es el coeficiente de alometría, y que ha sido utilizada ampliamente en numerosas disciplinas: fisiología, morfogénesis, evolución, etc. (Pélabon et al., 2014).
En el presente trabajo se pretende describir las características ontogénicas del peso de determinados elementos óseos de los corderos, desde los 3 meses de edad hasta que alcanzan la plena madurez. Así, el análisis de regresión simple indica que existe una alometría ontogénica, debido al cambio con la edad diferencial de distintos componentes del cuerpo, donde machos y hembras presentan un patrón de cambio morfológico similar, aunque no idéntico, sobre todo en lo que se refiere al peso vivo final. Con la edad se produce más crecimiento (especial incremento en lo ponderal y en el perímetro torácico) que desarrollo (cambio de conformación), aunque en ningún caso lineal; o dicho de otra forma, la relación de todas las variables corporales con la edad no mantiene una proporcionalidad estricta. Tanto el crecimiento como el desarrollo no son isométricos.
Que no haya diferencias en las rectas de regresión entre sexos sugiere, además, que el escalamiento ontogénico no es en el Borrego de Chiapas un mecanismo importante de diferenciación sexual, aunque comparativamente los machos crecen más rápido que las hembras debido a la mayor potencia de los andrógenos con respecto a los estrógenos. No obstante, en lo que se refiere al desarrollo no hay diferencias de ningún tipo. O sea, que a la misma edad, tanto machos como hembras tendrán tamaño corporal similar y, por ende, un dimorfismo sexual poco marcado, una forma similar.
El grado de dimorfismo sexual depende de muchos factores (Frayer y Wolpoff, 1985), pero uno de los principales se relaciona a la malnutrición proteico-calórica, que actúa reduciendo las diferencias en tamaño entre los sexos (Pucciarelli, 1980). En la especie humana, en caso de estrés nutricional, los hombres son más susceptibles a este estrés, por lo que reducen su tamaño, mientras que las mujeres son más estables (Stini, 1969). En el ovino, al ser el macho más inmaduro al momento de nacer, puede ser afectado por una deficiencia nutricional; en cambio, la hembra resulta más resistente a penurias alimenticias. Entonces, condiciones nutricionales subóptimas en el caso del Borrego de Chiapas podrían ser las que explicasen las escasas diferencias en el desarrollo de su dimorfismo sexual. Cabe preguntarse, finalmente, sobre la relevancia de la «pedomorfia» o
«pedomorfosis», proceso de retraso del desarrollo ontogenético que podría conducir a una morfología adulta con retención de aspectos llamativos de la morfología infantil; en este caso, las relacionadas con las de desarrollo. Análisis histológicos y osteológicos podrían ser concluyentes para confirmar o rechazar estas hipótesis.
LITERATURA CITADA
1. Anzelmo M, Sardi ML, Barbeito-Andrés J, Pucciarelli HM. 2012. Alometrías ontogénicas y dimorfismo sexual facial en dos poblaciones humanas modernas. Rev Arg Antropol Biol 14(1): 89-100. [ Links ]
2. Frayer DW, Wolpoff MH. 1985. Sexual dimorphism. Annu Rev Anthropol 14: 429-473. doi: 10.1146/annurev.an.14.100185.002241 [ Links ]
3. Gould SJ. 1966. Allometry and size in ontogeny and phylogeny. Biol Revs 41: 587-640. doi: 10.1111/j.1469-185X.1966-tb01624.x [ Links ]
4. Hammer Ø, Harper DAT, Ryan RD. 2010. PAST. Palaeontol Stat 4(1). v 2.17c. [Internet]. Disponible en: http://folk.uio.no/ohammer/past/ [ Links ]
5. Hammond J. 1960. Farm animals. London: Edward Arnold Publ Ltd. 296 p. [ Links ]
6. Huxley JS. 1972. Problems of relative growth. New York: Dover Inc. 312 p. [ Links ]
7. Pelábon C, Bolstad GH, Egset CK, Cheverud JM, Pavlicev M, Rosenqvist G. 2013. On the relationship between ontogenetic and static allometry. Am Nat 181: 195-212. doi: 10.1086/668820 [ Links ]
8. Pelábon C, Firmat C, Bolstad GH, Voje KL, Houle D, Cassara J, Rouzic AL, Hansen TF. 2014. Evolution of morphological allometry. Ann NY Acad Sci 1320: 58-75. doi: 10.1111/nyas.12470 [ Links ]
9. Pucciarelli HM. 1980. The effect of race, sex and nutrition on craniofacial differentiation in rats. A multivariate analysis. Am J Phys Anthropol 53: 359- 368. doi: 10.1002/ajpa.1330530307 [ Links ]
10. Rodero E, González A. 2009. Las regiones corporales de los animales domésticos. En: Sañudo C (ed). Valoración morfológica de los animales domésticos. Madrid: Ministerio de Medio Ambiente y Medio Rural y Marino. p 107-144. [ Links ]
11. Stini WA. 1969. Nutritional stress and growth: sex difference in adaptive response. Am J Phys Anthropol 31: 417- 426. doi: 10.1002/ajpa.1330310316 [ Links ]
Recibido: 9 de junio de 2016
Aceptado para publicación: 22 de octubre de 2016
