











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Los sistemas ganaderos tropicales en el Perú se caracterizan por ser silvopastoriles, combinando pasto, árboles y animales. La presencia de árboles dentro de los potreros, cumplen un rol importante en el sistema, brindando sombra y forraje para los animales, además de ser un componente importante al estar en simbiosis con el pasto. El pasto guinea (Panicum maximum Jacq) es la especie naturalizada predominante en ecosistemas de selva alta (Nuñez, 2017). Así mismo, en dichas áreas es frecuente la deforestación, tala y quema indiscriminada de los bosques para el establecimiento de pasturas, lo cual trae una disminución de la calidad, productividad y diversidad de la vegetación (Oliva, 2016).
El uso se especies arbóreas en los sistemas silvopastoriles (SSP) ofrece ventajas para el desarrollo de ganaderías con menor impacto ambiental. Una de las bondades de SSP es la obtención de una mejor calidad y cantidad de pasto (Obispo et al., 2008), permitiendo un mayor rendimiento en los sistemas ganaderos (FAO, 2012). Los SSP presentan también un gran potencial para recuperación de áreas de pasturas degradadas (Alegre et al., 2012); sin embargo, se requiere determinar las interacciones entre sus componentes para asegurar el correcto funcionamiento del sistema.
Panicum maximum es una especie tolerante al pisoteo y la sequía, y es un alto productor de forraje de buena calidad, palatabilidad y digestibilidad; desarrollándose muy bien en SSP con una clara ventaja sobre otras especies de pasto, porque tiene la capacidad de soportar ciertos niveles de sombra sin interferir en su producción (Ledesma, 2006). No obstante, su rendimiento se ve afectado por el régimen de lluvias, mostrando niveles inferiores en épocas secas (Núñez, 2017). Por otro lado, se tiene evidencia de especies de pasto que crecen adecuadamen-te bajo la sombra de otras plantas (Abraham et al., 2014).
El valor nutritivo de la planta declina a medida que avanza en edad debido a un incremento en su lignificación, aumento de la fibra detergente neutro y menor contenido de proteína, así como una disminución del número de hojas (García, 2002). Así mismo, la sombra también puede provocar cambios morfológicos y fenológicos en la especie forrajera, los cuales funcionan como mecanismo de adaptación a la baja incidencia de energía lumínica y la consiguiente reducción en el potencial fotosintético de la planta. Para compensar esto, la especie forrajera tiende a desarrollar hojas largas, pero menos gruesas para una mejor captación de luz y una menor tasa de respiración (Wilson y Ludlow, 1991).
No existen estudios sobre los beneficios de los árboles como el faique (Acacia macracantha) dentro de los potreros en praderas tropicales, de allí que el objetivo del estudio fue determinar las tasas de crecimiento y el valor nutricional del pasto bajo la copa de los árboles y fuera de ella en un sistema silvopastoril, donde el componente arbóreo fue el faique y el componente forrajero fue el pasto guinea (Panicum maximum Jacq.) para determinar su importancia dentro del sistema de producción pecuario de la zona de Cajamarca, Perú.
MATERIALES Y MÉTODOS
Localización
La fase experimental se realizó en la región de selva alta (2016), distrito de Bellavista, provincia de Jaén, Región Cajamarca (Perú), a 729 msnm. La zona está clasificada de acuerdo con Holdridge (2000) comobosqueseco premontanotropical(bs-PT). La temperatura media anual es de 24.2 °C con máximas y mínimas promedio de 34.0 y 18.5 ºC, respectivamente. La precipitación anual es 810 mm, siendo la mayor en marzo (197 mm) y la menor en noviembre (16 mm) (SENAMHI, 2018). El componente pasto contempló 8 ha de Panicum maximum Jacq. (monocultivo naturalizado), establecido hace 20 años, en asociación con la especie arbórea leguminosa nativa Acacia macracantha, conocida comúnmente como faique.
El sistema es manejado en secano (sin riego) con sistemas de pastoreo rotativo cada seis semanas. La pastura residual tuvo una altura promedio de 60 y 20 cm en época lluviosa y seca, respectivamente. El componente animal estuvo representado por ganado vacuno criollo de doble propósito. La evaluación se realizó en la época lluviosa (marzo) y seca (agosto) de 2017.
Análisis de Suelo
El muestreo de suelo (6 muestras en total) se hizo a 30 cm de distancia del punto de colección de las muestras de pasto, pero a 25 a 30 cm de profundidad, que es donde se encuentra la mayor proporción del sistema radicular del Panicum maximum Jacq. La humedad del suelo se midió por gravimetría (Martínez, 1971). Las muestras consistieron en 1 kg de suelo utilizando envases con cierre hermético para evitar la pérdida de humedad, y luego fueron enviadas al Laboratorio de Suelos, Plantas, Agua y Fertilizantes de la Universidad NacionalAgraria la Molina, Lima donde se colocaron en estufa a 105 °C durante 24 h. La temperatura del suelo se midió dentro del excluidor de crecimiento del pasto, a una profundidad de 15 a 20 cm de la superficie, mediante un geotermómetro (Reotem®, rango: -10 hasta 90 ºC, con 0.5 ºC de precisión) por 5 minutos.
Categorización de los Árboles
La densidad promedio fue de 80 árboles dispersos por hectárea. La categorización del tamaño de los árboles se hizo mediante la medición del Diámetros a la Altura del Pecho (DAP), según la FAO (2004), utilizando la siguiente ecuación: D = C/π, donde D: diámetro (cm), C: circunferencia (cm), π: 3.1416.
Para ello se utilizó una cinta métrica para medir la circunferencia (C) a la altura del pecho (1.3 m).
Las categorías del tamaño de los árboles por hectárea fueron: árboles en regeneración (DAP <7.5 cm) 13.4%, jóvenes (DAP 7.5-24 cm) 50.6%, pequeños (DAP 25-37 cm) 17.3% y medianos (DAP 38-49 cm) 18.5%. Los muestreos de pasto y suelo se realizaron en la cuarta semana de rebrote del pasto, en época lluviosa (enero-marzo) y seca (julio-setiembre).
Crecimiento del Pasto
Se utilizaron cuatro potreros de 2 ha cada uno. En cada portero se instalaron tres excluidores (4 m2) a base de alambre de púas para evitar el pastoreo, distribuidas de la siguiente manera: fuera la copa del árbol (FCA), bajo la copa del árbol joven (BCAj) y bajo la copa del árbol mediano (BCAm). El área de muestreo fue de 1 m2 para eliminar el efecto del borde (ASRM, 1962). La tasa de crecimiento (método de corte y separación; Geenty y Rattray, 1987) fue el forraje disponible dentro del excluidor un mes después del corte (10 cm del nivel del suelo), expresado en kilogramos de forraje seco por hectárea por día (kg MS/ha/d). Para el pesaje se utilizó una balanza digital OHAUS®, con capacidad de 3000 g y precisión de 0.1 g.
Valor Nutritivo del Pasto
Se colectaron 12 muestras de pasto bajo la copa de los árboles y fuera de ellos. Se muestreó a tres distancias: 1, 2, 4 m del tronco de los árboles. Las muestras fueron secadas a 60 °C durante 48 horas, y luego fueron pasadas por el molino (Wiley®) con tamiz de 1 mm. Se determinó: proteína cruda (PC) por el método Kjeldahl (AOAC, 2012), fibra detergente neutro (FDN) por el método Van Soest (Van Soest, 1985), digestibilidad in vitro de la materia seca (DIVMS) por el método de Tilley y Terry (1963), calcio y fósforo por el método digestión ácida (AOAC, 2016).
Análisis Estadístico
El análisis de variancia de los valores contenidos de tasa de crecimiento, PC, FDN, calcio, fósforo y DIVMS se hizo bajo un diseño de bloques completamente al azar con arreglo factorial 3 x 2, donde los bloques fueron los potreros, los factores fueron las épocas del año y la ubicación del pasto. Para la evaluación de comparaciones individuales de los efectos se utilizó la prueba de Duncan a un nivel de significancia de 0.05. Se utilizó el paquete estadístico R.
RESULTADOS Y DISCUSIÓN
Características del Suelo
El Cuadro 1 indica que los suelos del área experimental eran moderadamente alcalinos con valores de pH entre 8.01 y 8.23 para las tres áreas de medición, siendo ligeramente superior a los valores reportados por Núñez (2017) en la misma zona, pero sin considerar la presencia de árboles. La textura del suelo fue la misma para las tres áreas evaluadas con lecturas Franco-Arcillo-Arenoso, lo que demuestra que son suelos de textura moderadamente fina, buena aireación y disponibilidad de agua, ayudando a los microorganismos en el suelo a sobrevivir, beneficiando la absorción de nutrientes y favoreciendo el desarrollo de las plantas (Navarro, 2013). La cantidad de materia orgánica en el suelo aumenta bajo la copa de los árboles, debido a que esta especie de árbol es de tipo caducifolia (reciclaje de hojarasca).
Cuadro 1 Características del suelo alrededor de árboles de faique (Acacia macracantha)
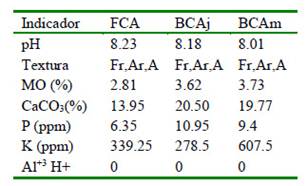
Fuera de la copa del árbol (FCA), bajo la copa del árbol joven (BCAj), bajo la copa del árbol mediano (BCAm) Materia orgánica (MO)
Los niveles de fósforo y calcio en el suelo bajo la copa de los arboles fueron mayores (Cuadro 1), debido a que la raíces de los arboles tiende a extraer minerales de capas más profundas del suelo, efecto de bombeo de nutrientes (Sadeghian et al., 1998). Similar tendencia mostró el potasio. No se detectaron niveles de aluminio en los suelos debido a que son suelos ligeramente alcalinos.
Variables de Suelo
La temperatura del suelo fue estadísticamente inferior (p<0.05) fuera de la copa de los árboles, habiendo 12.6 y 8.9% mayor temperatura en épocas lluviosa y seca, respectivamente; mientras que la humedad del suelo fue menor fuera de la copa de los árboles en épocas lluviosa (7.0%) y seca (34%) (Cuadro 2). La mayor humedad bajo la copa de los árboles crea un microclima apropiado para el normal crecimiento de la pastura en esta zona, siendo crucial en las épocas de menor precipitación. La mayor humedad del suelo bajo la copa de los árboles se debe a la disminución de la evaporación del suelo y la evapotranspiración de la planta (Ludwig et al., 2001).
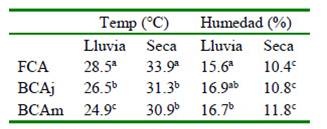
Tasa de Crecimiento
La tasa de crecimiento fue significativamente mayor en la época lluviosa (71.1, 58.8 y 51.2 kg MS/ha/día fuera de la copa, bajo la copa del árbol joven y mediano, respectivamente) que en la época (29.6, 24.1 y 19.1 kg MS/ha/día, respectivamente) (p<0.05; Figura 1). La disminución de la tasa de crecimiento bajo la copa de los árboles se debe a que los pastos guinea son de ciclo fotosintético tipo C4, alcanzando su máxima producción con altos niveles de intensidad lumínica (Obispo et al., 2008). Así mismo, el mayor crecimiento fuera de la copa de los árboles está relacionado con la menor calidad del pasto, debido a la rápida acumulación de lignina y celulosa en sus tejidos (Nuñez et al., 2019).
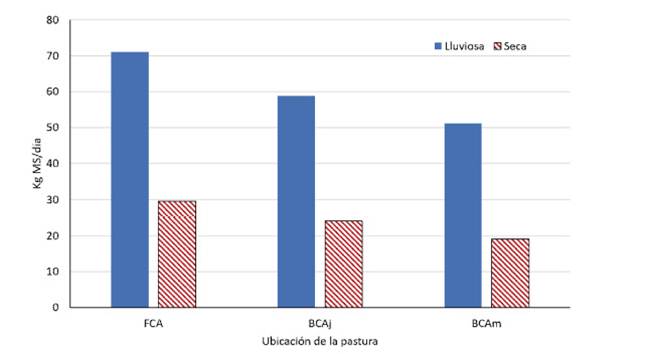
Figura 1 Tasa de crecimiento (kg MS/ha/día) de la pastura Panicum maximum Jacq, en tres ubicaciones: Fuera de la copa del árbol (FCA), bajo la copa del árbol joven (BCAj), bajo la copa del árbol mediano (BCAm)
Las tasas de crecimiento encontradas para las dos épocas son superiores a los reportadas por Nuñez (2017) a 30 días de rebrote, quien reporta valores de 46.3, 18.4 y 8.2 kg MS/ha/día en las épocas lluviosa, inicio de lluvias y lluviosa respectivamente, sin considerar la ubicación del pasto respecto a la presencia de árboles. En el trópico peruano (ceja de selva), las tasas de crecimiento y calidad del forraje están sujetas a las precipitaciones pluviales, alcanzando una capacidad de carga de las pasturas de hasta 3.0 UA/ha (Nuñez et al., 2019).
Proteína Cruda
El porcentaje de PC fue mayor en la época lluviosa en comparación con la época seca (p<0.05). Así mismo, estos valores se incrementan con la edad del árbol (Cuadro 3), lo cual explica la mejora en la calidad del pasto al mantener el faique en los potreros. Esta mejora significa 3.3 y +3.1% de proteína adicional al mantener los árboles medianos (38-49 cm, DAP) en la época lluviosa y seca, respectivamente.
Cuadro 3 Valores de proteína cruda (%) del Panicum maximum Jacq. alrededor de árboles de faique (Acacia macracantha)
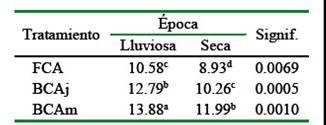
Fuera de la copa del árbol (FCA), bajo la copa del árbol joven (BCAj), bajo la copa del árbol mediano (BCAm)
Los altos contenidos de PC en los pastizales bajo la copa de los arboles podrían estar asociados con un aumento del flujo de nitrógeno en el suelo, especialmente cuando la especie arbórea es una leguminosa que tiene potencial para contribuir a la fijación biológica de N (Xavier et al., 2014); además, los pastos bajo sombra son fisiológicamente más jóvenes que los pastos a pleno sol. En este sentido, Ribaski y Menezes (2002) encontraron una mejora sustancial en PC (+16.27%), al evaluar el valor nutritivo del pasto bufel (Cenchrus ciliaris) bajo la copa del algarrobo (Prosopis juliflora).
Fibra Detergente Neutro (FDN) y Digestibilidad in vitro de la Materia Seca (DIVMS)
Los valores de FDN Panicum maximum entre tratamientos en la época lluviosa fueron estadísticamente similares, aunque con tendencia a ser menores al incrementar los niveles de sombra. Sin embargo, en la época seca el valor promedio de la FDN disminuyó con la sombra (p<0.05; Cuadro 4). La ligera disminución de FDN registrada en este estudio bajo sombra de los árboles en época lluviosa podría estar asociada con las paredes celulares más delgadas del esclerénquima en plantas que se cultivan en sombra que aquellas en condiciones de pleno sol (Deinum et al.,1996), ya que estos pastos presentan hojas más delgadas. Como dato comparativo se puede mencionar el trabajo de Paciullo et al. (2016), que en niveles de sombra de 0, 37 y 58% reportaron valores de 65.2, 64.2 y 64.2% de FDN.
Cuadro 4 Valores de fibra detergente neutro (FDN) y digestibilidad in vitro de la materia seca (DIVMS) del Panicum maximum Jacq. alrededor de árboles de faique (Acacia macracantha)
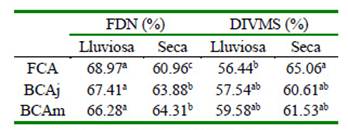
Fuera de la copa del árbol (FCA), bajo la copa del árbol joven (BCAj), bajo la copa del árbol mediano (BCAm) a,b,c Letras diferentes dentro de columnas indican diferencias significativas (p<0.05)
La DIVMS no presentó diferencias estadísticas entre tratamientos, aunque los valores fueron ligeramente superiores bajo la copa de los árboles en la época lluviosa, coincidiendo con el estudio de Paciullo et al. (2011). Por su parte, Ribaski y Menezes (2002), indican que en el pasto bufel (Cenchrus ciliaris), la DIVMS llega a 44.1% cuando se encuentra próximo al tronco del algarrobo (Prosopis juliflora), en comparación con el 41.1% obtenido fuera de la copa del árbol.
Calcio y Fósforo
El contenido de calcio en el pasto presentó diferencias estadísticas (p<0.05) entre tratamientos, alcanzando mayores valores bajo la copa del árbol joven (0.83%) y bajo la copa del árbol mediano (0.92%) en la época lluviosa (Cuadro 5). En la época seca se presentó la misma tendencia, pero sin diferencia estadística significativa. Por su parte, Ribaski y Menezes (2002) reportan tendencias similares bajo y fuera de la copa del árbol (2.1 y 1.9%, respectivamente en el pasto bufel (Cenchrus ciliaris) bajo la copa del algarrobo (Prosopis juliflora). Este comportamiento se debe probablemente por la capacidad de la especie arbórea (raíz pivotante y profunda) para extraer el calcio desde la parte más profunda, sumado al reciclaje de la hojarasca que es aprovechada por la pastura (Sadeghian et al., 1998; Romero 2018).
La cantidad de fósforo en la pastura presentó diferencias estadísticas (p<0.05) para el caso de fuera de la copa del árbol (0.19%) y bajo la copa del árbol mediano (0.24%) en la época lluviosa, mientras que mostró similar tendencia en la época seca, pero sin diferencias significativas (Cuadro 5).
Cuadro 5 Valores de calcio y fosforo del Panicum maximum Jacq. alrededor de árboles de faique (Acacia macracantha)
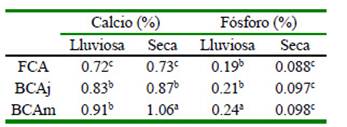
Fuera de la copa del árbol (FCA), bajo la copa del árbol joven (BCAj), bajo la copa del árbol mediano (BCAm) a,b,c Letras diferentes dentro de variables indican diferencias significativas (p<0.05)
La menor disponibilidad del fósforo en la pastura en época seca se debe a su poca disponibilidad en el suelo (Cuadro 1), sumado a que el agua constituye un vehículo natural de transporte (difusión) del fósforo en el suelo. La mayor disponibilidad (flujo) estacional (época lluviosa) favorece la absorción por la pastura (Casanova y Eduardo, 2005). Similar comportamiento estacional en este sistema silvopastoril fue encontrado por Núñez (2017). El fósforo es un mineral importante en la alimentación de los rumiantes; sin embargo, su concentración en el pasto puede estar alrededor de 0.2% (Tejos, 2001).
CONCLUSIONES
( La presencia de árboles dentro de los potreros mejoró el contenido nutricional de la pastura en términos de proteína (+3.3 y +3.06%), calcio (+0.03 y +0.1%) y fósforo (+0.19 y +0.33%) en época lluviosa y seca, respectivamente.
( La presencia de árboles dentro de los potreros mejoró el porcentaje de nutrientes (calcio: +30.7% y fósforo: +37.69%) disponibles en el suelo, aumentando su fertilidad; lo cual se reflejó en la mejor calidad nutricional de la pastura.

