











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La captura, producción y consumo de especies acuícolas presenta un constante y acelerado incremento global. Según los datos recopilados por la FAO y presentados en su reporte sobre el "Estado mundial de la pesca y la acuicultura" en 2018, la producción mundial alcanzó cerca de 171 millones de toneladas durante el año 2016, donde la acuicultura, incluido el camarón, aportó el 47%. Igualmente, en términos de consumo de peces y mariscos mundial, se presentó un aumento per cápita desde 9.0 kg en 1961 hasta 20.2 kg en 2015, con un incremento medio aproximado de 1.5% anual (FAO, 2018). Ese reporte indica que el aporte mundial de las capturas de especies acuícolas se encuentra prácticamente estancado, con un promedio anual de 90.9 ± 1.27 millones de toneladas, en tanto que el aporte de la acuicultura, en sus diferentes modalidades, presenta un crecimiento anual promedio de 3.64 millones de toneladas para el mismo periodo, llegando a los 80 millones de toneladas (FAO, 2018), tal como se observa en la Figura 1.
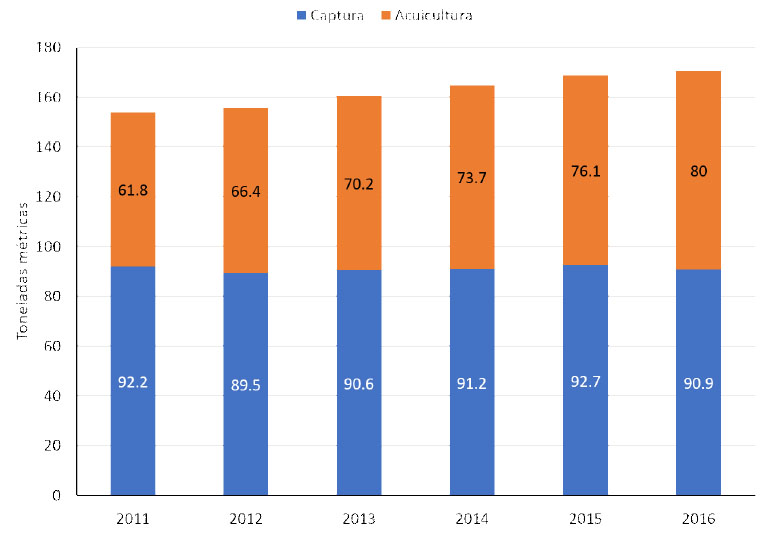
Figura 1 Aportes relativos de las capturas naturales y de la acuicultura durante el periodo 2011-2016 (en millones de toneladas métricas). Fuente: FAO (2018)
A pesar de estos datos, la incidencia de enfermedades infecciosas en los cultivos, constituyen la principal amenaza para la camaronicultura (Bondad-Reantaso et al., 2001; Aguirre-Guzmán et al., 2005; Morales-Covarrubias, 2010; Cuéllar-Anjel et al., 2014; Gómez-Gil et al., 2015; Peña-Navarro y Varela-Mejías, 2016; Morales-Covarrubias et al, 2018a). Los camarones de cultivo son frecuentemente atacados por múltiples agentes infecciosos, incluyendo diferentes especies de virus, bacterias, hongos y parásitos (Lightner, 1996; Leyva-Ordaz et al., 2010; Cuéllar-Anjel et al., 2014; OIE, 2018).
El incremento en la incidencia de los brotes infecciosos y su impacto en la producción ha dado lugar al desarrollo gradual de diversas técnicas diagnósticas (Prieto y Rodríguez, 1993; Lightner, 1996), las cuales han respondido a las necesidades específicas del momento de su implementación. Así, de procedimientos básicos, se ha ido evolucionando hacia ensayos diagnósticos más rigurosos y específicos, lo cual ha facilitado el abordaje de los eventos infecciosos, siendo común la combinación de técnicas (Lightner, 1996; Cuéllar-Anjel, 2008). Entre estas, se tienen técnicas y procedimientos para el análisis clínico y en fresco, pruebas microbiológicas, histopatológicas, de bioensayo, uso de anticuerpos, hibridación in situ, inmunohistoquímica y las técnicas moleculares de PCR (Bell y Lightner, 1988; Prieto y Rodríguez, 1993; Lightner, 1996; Morales-Covarrubias, 2013; OIE, 2018).
Delimitando el alcance de este documento al diagnóstico bacteriano, se encuentran una gran diversidad de agentes involucrados en brotes, incluyendo múltiples especies y cepas de Vibrio spp (Prieto y Rodríguez, 1993; Lightner, 1996; Morales-Covarrubias, 2013), así como bacterias Gram positivas, incluyendo estreptococos (Hasson et al., 2009; Morales-Covarrubias et al., 2018a); espiroplasmas (Nunan et al., 2004; Heres et al., 2011) y bacterias intracelulares tipo Rickettsia como la Hepatobacter penaei (Lightner, 1996; Morales-Covarrubias, 2013; OIE, 2018).
Esta diversidad etiológica representa un verdadero desafío para la obtención de un diagnóstico certero, ya que las técnicas disponibles no se ajustan, necesariamente, a todos los casos. Asimismo, la aparición de patologías emergentes con diferencias genotípicas o fenotípicas sutiles, así como los tropismos y lesiones que presentan (Lightner, 1996; Nunan et al., 2004; Cuéllar-Anjel et al., 2014; Varela-Mejías, 2018) complican el panorama. Con el fin de exponer este tema, se presenta un resumen no exhaustivo de las principales patologías bacterianas reportadas para camarones de cultivo durante las etapas de engorda, así como sus principales características, lesiones y técnicas diagnósticas comúnmente utilizadas.
VIBRIOSIS SISTÉMICA
Esta es una de las primeras enfermedades bacterianas reportada para camarones en Latinoamérica. Puede causar mortalidades superiores al 90% de las poblaciones afectadas (Prieto y Rodríguez, 1993; Lightner, 1996).
Como agentes causales, han sido reportadas múltiples especies del género Vibrio, incluyendo, pero no limitándose, a cepas de Vibrio harveyi, V. parahaemolyticus, V. vulnificus, V. penaeicida, V. nigripulchritudo, V. alginolyticus, V. owensii y V. campbellii (Lightner, 1988; Prieto y Rodríguez, 1993; Lightner, 1996; Soto et al., 2010; Morales-Covarrubias y Gómez-Gil, 2014; Gómez-Gil et al., 2015; Cuéllar-Anjel y Brock, 2018).
Sus signos clínicos son inespecíficos e incluyen la presencia de camarones nadando erráticamente en la superficie y al borde de los estanques, reducción en el consumo de alimento, así como debilidad y mortalidades crecientes. En algunos brotes es posible el desarrollo de bioluminiscencia, dependiendo de la cepa causante (Johnson, 1995; Lightner, 1996; Peña-Navarro y Varela-Mejías, 2016).
El diagnóstico presuntivo de esta enfermedad puede realizarse mediante análisis en fresco. Para ello, es conveniente combinar las observaciones en campo, así como las observaciones macroscópicas y microscópicas de los animales que se van a analizar. Al microscopio, es posible observar bacterias mótiles en hepatopáncreas y hemolinfa (Lightner, 1996; Morales-Covarrubias y Gómez-Gil, 2014). Adicionalmente, se puede afectar el tiempo de coagulación de la hemolinfa, extendiendo el tiempo requerido debido a la presencia de bacterias (Morales y Cuéllar-Anjel, 2014).
En el cultivo bacteriológico se pueden utilizar tanto medios generales, como el Agar Marino, como medios selectivo-diferenciales como el agar TCBS (Agar Tiosulfato Citrato Sacarosa, por sus siglas en inglés) y el Cromagar. Estos análisis se realizan en muestras de animales; hemolinfa u órganos, así como en agua, superficies y sedimentos de los estanques de cultivo (Prieto y Rodríguez, 1993; Lightner, 1996; Morales-Covarrubias y Gómez-Gil, 2014; Gómez-Gil et al., 2015).
Histopatológicamente, durante la vibriosis sistémica se detectan nódulos hemocíticos que en ocasiones pueden estar melanizados, con centros sépticos y ubicados en múltiples tejidos, incluyendo branquias, hepatopáncreas, glándula antenal, músculo esquelético, corazón, senos hemales, lóbulos hematopoyéticos, órgano linfoide y gónadas, entre otros (Lightner, 1996; Morales-Covarrubias, 2004; Peña-Navarro y Varela-Mejías, 2014).
El hepatopáncreas puede presentar daños en las células epiteliales, las cuales en algunos casos estarían hipertrofiadas, con los núcleos picnóticos y hay necrosis tubular, unido a una fuerte infiltración hemocítica. Es posible observar bacterias extracelulares individuales o en cúmulos, presentes como masas basofílicas (Martin et al., 2004; Morales-Covarrubias, 2004; Peña-Navarro y Varela-Mejías, 2014; Varela-Mejías, 2018).
NECROSIS AGUDA DEL HEPATOPÁNCREAS (APHND)
Necrosis Aguda del Hepatopáncreas (AHPND)
Se trata de un caso particular de vibriosis, reportándose inicialmente en Asia en 2009, causando altas mortalidades en camarones Penaeus vannamei y P. monodon en China durante los primeros 35 días de cultivo, para luego propagarse a Vietnam en 2010, Malasia en 2011 y Tailandia en 2012 (NACA, 2012; Cuellar-Anjel, 2013, FAO, 2013), así como a Filipinas (de la Peña et al., 2015).
Es causada por cepas de Vibrio. La primera especie identificada fue el V. parahaemolyticus. Las cepas contienen un plásmido de aproximadamente 69 kb, portando genes que codifican para una toxina binaria, llamada PirA y PirB (Tran et al., 2013; Kondo et al., 2015; Yang et al., 2014; Han et al., 2015; Lee et al., 2015). Actualmente han sido confirmadas cepas de V. parahaemolyticus, V. harveyi, V. campbellii y V. owensii, sin descartar la aparición de nuevas especies o cepas (Tran et al., 2013; Joshi et al., 2014; Pantoja y Lightner, 2014a; Han et al., 2015; Kondo et al., 2015; Ahn et al., 2017; Varela-Mejías et al., 2017; CuéllarAnjel y Brock, 2018; Liu et al., 2018; VarelaMejías, 2018).
AHPND fue detectado en México en 2013, generando mortalidades cercanas al 100% y disminución de producción (Nunan et al., 2014; Pantoja y Lightner, 2014a), continuando su avance por el continente, para detectarse años más tarde en América del Sur (Restrepo et al., 2016; Ahn et al., 2017; Saavedra-Olivos et al., 2018). En 2016, el AHPND fue incluido en la lista de enfermedades de declaración obligatoria de la Organización Mundial de Salud Animal (OIE, 2018).
Su signología clínica no es específica. Incluye una marcada palidez en el hepatopáncreas, atrofia del órgano, debilidad e intestino vacío o con contenido entrecortado (Tran et al., 2013; Pantoja y Lightner, 2014a; OIE, 2018). Mediante análisis en fresco, durante la fase aguda, es posible observar hepatopáncreas con deformaciones tubulares severas, con desprendimientos celulares masivos en ausencia de melanosis, especialmente en las fases avanzadas de la enfermedad (Morales-Covarrubias et al., 2018b).
Histopatológicamente, el AHPND se inicia con desprendimientos de las células epiteliales de los túbulos hepatopancreáticos y disminución de la mitosis. Hay una reducción de las reservas, atrofiando al hepatopáncreas. Las lesiones avanzan desde las zonas proximales de los túbulos hacia las zonas distales (Pantoja y Lightner, 2014a). En fases más avanzadas se observan bacterias basófilas, mayormente oportunistas. Luego continúa el desprendimiento celular y se inician los procesos inflamatorios con acumulación de hemocitos (Cuéllar-Anjel et al., 2012; Pantoja y Lightner, 2014a; Varela-Mejías y Peña-Navarro, 2014). En la fase terminal se generaliza la destrucción del hepatopáncreas por los desprendimientos celulares y por la acción bacteriana. Se observa una severa melanosis y necrosis multifocal, edema e infecciones bacterianas secundarias (CuéllarAnjel et al., 2012; Lightner et al., 2013; Tran et al., 2013; Pantoja y Lightner, 2014a). En esta etapa, es muy difícil distinguir entre AHPND y una necrosis séptica del hepatopáncreas, requiriéndose de técnicas de confirmación adicionales (Cuéllar-Anjel et al., 2012; Varela-Mejías y Peña-Navarro, 2016; Varela-Mejías et al., 2017).
El diagnóstico molecular del AHPND, requiere de ir más allá de identificar a la especie bacteriana. Se debe basar en la detección de las secuencias que codifican para las toxinas PirA y PirB, presentes en un plásmido extracromosómico (Tran et al., 2013; Han et al., 2015; Lee et al., 2015).
ESPIROPLASMOSIS
A inicios de 2002 se presentaron fuertes mortalidades atípicas en camarones P. vannamei cultivados en la costa del Caribe colombiano. Durante el siguiente periodo de siembra del mismo año, estanques adicionales en la misma granja experimentaron altas mortalidades. El agente causal identificado fue una cepa de Spiroplasma patógena (Spiroplasma penaei sp. nov.), la cual se propagó a granjas vecinas, generando mortalidades de hasta 90% de las poblaciones afectadas. Este fue el primer caso reportado de espiroplasmas patogénicas aisladas de crustáceos (Nunan et al., 2004; 2005; Altamiranda et al., 2011; Pantoja y Lightner, 2014b).
Estas bacterias poseen formas helicoidales y son móviles, pese a no presentar flagelos. De 195 nm de diámetro promedio y sin pared celular verdadera. Su rango de temperatura de crecimiento es de 20 a 37 ºC y el crecimiento óptimo ocurre en 28 ºC en caldo de M1D suplementado con 2% cloruro de sodio (Nunan et al., 2005; Pantoja y Lightner, 2014b). Clínicamente, los camarones infectados no muestran lesiones específicas. La condición de "camarones parados" ha sido reportada, donde camarones muertos se encuentran flotando como si estuvieran parados sobre la cola (Pantoja y Lightner, 2014b).
Su histopatología revela la presencia de lesiones típicas de una infección bacteriana sistémica. Esta incluye reacciones inflamatorias como formación de nódulos hemocíticos y fagocitosis, en ocasiones acompañada de melanizaciones y fibrosis. Entre los órganos y tejidos afectados se encuentran el cordón nervioso ventral, el músculo esquelético, corazón, glándula antenal, órgano linfoide y el tejido conectivo fibroso del hepatopáncreas, branquias y tejido subcuticular de apéndices (Nunan et al., 2004; Heres et al., 2011; Pantoja y Lightner, 2014b; MoralesCovarrubias et al., 2018b).
La presencia de S. penaei puede ser verificada en las lesiones observadas usando hibridación in situ con sondas de ADN específicas (Heres et al., 2011; Lightner y Pantoja, 2014; Morales-Covarrubias et al., 2018b). Además, se dispone de métodos moleculares de PCR convencional y PCR en tiempo real (Nunan et al., 2004, 2005; Altamiranda et al., 2011; Pantoja y Lightner, 2014b).
HEPATOPANCREATITIS NECROTIZANTE
La hepatopancreatitis necrotizante (NHP) se detectó inicialmente en Texas, USA, en 1985, detectándose posteriormente en Centro y Sur América (Lightner, 1996; Vincent y Lotz, 2007; Morales-Covarrubias, 2010; OIE, 2018).
El NHP es causado por la bacteria intracelular Hepatobacter penaei, Gram negativa, mótil mediante flagelos. Presenta dos morfotipos, la helicoidal de 0.25 x 3.5 ìm y la forma bacilar predominante de 0.25 x 0.9 ìm (Lightner, 1996; Nunan et al., 2013; Morales y Cuellar-Anjel, 2014). Esta bacteria infecta y se multiplica en el citoplasma de las células epiteliales del hepatopáncreas, alcanzando altas concentraciones en los tejidos afectados (Lightner, 1996; MoralesCovarrubias, 2008; OIE, 2018).
Los signos clínicos para esta enfermedad tampoco poseen valor diagnóstico y se requiere análisis específicos. En brotes severos y persistentes puede causar mortalidades acumuladas superiores al 90% de las poblaciones afectadas, tanto en cultivos de Penaeus vannamei como de P. stylirostris (Lightner, 1996; Vincent y Lotz, 2007; Morales-Covarrubias, 2010, 2013; Morales y Cuellar-Anjel, 2014; OIE, 2018).
Estas bacterias, por ser intracelulares obligadas, no pueden ser cultivadas en los medios artificiales típicos, así que durante el abordaje de los posibles casos se recurre a un análisis clínico presuntivo. Los animales presentan hepatopáncreas atrofiados, cuerpos delgados y es común el oscurecimiento de los bordes de los pleópodos. La textura de la cutícula es áspera y hay pérdida de tono muscular (Lightner, 1996; MoralesCovarrubias, 2010; OIE, 2018). El diagnóstico presuntivo del NHP se basa en la signología clínica de los animales, la historia del caso y la observación mediante análisis en fresco de reducción de las reservas en hepatopáncreas, estrangulaciones tubulares, atrofia, melanizaciones y presencia de granulomas (Lightner, 1996; MoralesCovarrubias, 2010).
Histopatológicamente, el NHP causa una disminución inicial de las reservas, acompañado de un desprendimiento de células epiteliales de los túbulos hepatopancreáticos. En algunas células se observa material granular en los citoplasmas, compuesto por masas de bacterias basofílicas. La formación de granulomas basofílicos es característica. Además, se puede observar estrangulaciones tubulares, encapsulaciones de tejidos afectados y necrosis tubular multifocal (Lightner, 1996; Morales-Covarrubias, 2010; Peña-Navarro y Varela-Mejías, 2016, Varela-Mejías, 2018).
La confirmación del NHP se da mediante histopatología, hibridación in situ o PCR (Lightner, 1996; Morales-Covarrubias, 2010; Varela-Mejías y Peña-Navarro, 2016; OIE, 2018; Varela-Mejías, 2018).
ESTREPTOCOCOSIS
Estreptococosis
Contrario a lo que ocurre con las patologías causadas por especies Gram negativas, como Vibrio spp o el NHP, las infecciones por bacterias Gram positivas en crustáceos son poco comunes. Han sido reportadas en langostinos de agua dulce Macrobrachium rosenbergii (Hasson et al., 2009; Varela-Mejías y Valverde-Moya, 2018) y en langostas del género Homarus sp (Stewart et al., 2004). No obstante, son inusuales en camarones peneidos (Hasson et al., 2009; Morales-Covarrubias et al., 2018a).
En los brotes infecciosos por estreptococos no se reportan signos clínicos de valor diagnóstico. Los animales fuertemente infectados mueren y quedan en el fondo de los estanques. En ocasiones, los camarones muertos parecen tener las cutículas intactas, pero presentan sectores del músculo caudal ausente, lo que supondría una degradación de tejidos desde el interior hacia el exterior (Hasson et al., 2009). Estos autores determinaron por el perfil bioquímico que la cepa era Gram positiva, no hemolítica, no mótil, oxidasa y catalasa positivas, la cual es reconocida por cebadores (primers) universales para estreptococos. El perfil bioquímico no produjo resultados concluyentes, señalando entre las posibles especies a Streptococcus uberis y S. parauberis.
Por su parte, Morales-Covarrubias et al. (2018a), describen a la especie Streptococcus penaeicida, Gram positiva, no formadora de esporas, anaeróbica facultativa, catalasa negativa y no mótil. Sus colonias, desarrolladas en agar sangre, son circulares, no pigmentadas, con diámetros de 0.75-1.0 mm, y con actividad hemolítica a 37 ºC. Esta cepa puede crecer en ambientes entre 0 y 5.5% de cloruro de sodio y con pH entre 5.5 y 10.
Histológicamente, se observa la presencia de cocos Gram positivos libres en la hemolinfa, senos hemales y tejidos vascularizados. Se pueden presentar pequeños agregados hemocíticos y números variables de cocos extracelulares, así como hemocitos cargados de bacterias y pequeños nódulos en el corazón, branquias, músculo esquelético, intestino anterior y posterior, cordón nervioso, tejidos conectivos, senos hemales de la glándula antenal y hepatopáncreas. Se observa una fuerte vacuolización del órgano linfoide y una pérdida de su estructura normal (Hasson et al., 2009; MoralesCovarrubias et al., 2018a).
USOS Y LIMITANTES DE LAS TÉCNICAS
La diversidad de agentes infecciosos en los camarones de cultivo, sus mecanismos de acción y sus particularidades biopatológicas, constituye sin duda, un desafío en la obtención de diagnósticos rápidos, oportunos y confiables.
En el abordaje de la diagnosis, basado en la signología clínica en camarones, no es más que una técnica introductoria que permite suponer que "algo no está bien". La falta de especificidad y las similitudes clínicas entre las diferentes enfermedades hacen que su utilidad sea nula para la identificación concluyente de algún agente causal. Lightner (1996), por ejemplo, cita la presencia de hepatopáncreas pálidos o con volúmenes reducidos, anorexia y tractos vacíos durante los brotes de vibriosis sistémicas; sin embargo, estos signos también han sido descritos para otras patologías como el NHP (Vincent y Lotz, 2005) y el AHPND (Tran et al., 2013).
El análisis en fresco presenta algunas características que han facilitado su difusión y aplicación. Se requiere de poco equipo, la inversión de recursos es mínima y el tiempo de entrenamiento para el personal es corto. Este análisis puede ser realizado en granja y brinda información sobre el estado general de los camarones, principalmente sobre los parámetros relacionados a las condiciones del hepatopáncreas, así como a las parasitosis internas o infestaciones por epibiontes. El tiempo de respuesta es mínimo, lo que permite procesar un gran número de animales (Lightner, 1996; Morales-Covarrubias, 2010). Su principal limitante lo constituye la baja sensibilidad y especificidad del método. No obstante, en el caso específico de patologías bacterianas, es posible brindar resultados preliminares, con los cuales se pueden iniciar acciones correctivas en granja, pero requiere de otras técnicas más confiables para la obtención de resultados concluyentes. Además, se corre el riesgo de no detectar lesiones que se encuentren fuera de los tejidos procesados.
Las técnicas bacteriológicas tradicionales, basadas en el uso de medios de cultivo, y las pruebas de perfil bioquímico, han permitido desarrollar conocimientos de gran valor en lo referente a brotes por bacterias extracelulares. Prieto y Rodríguez (1993), Lightner (1996), Gómez-Gil et al. (2015), entre otros, presentaron modelos diagnósticos que actualmente continúan siendo de gran utilidad. La aplicación de medios de cultivo se utiliza en forma rutinaria, no solo como técnica diagnóstica, sino además, como soporte para enriquecimientos de cepas bacterianas, para su utilización posterior en otras técnicas; así Hasson et al. (2009) y Tran et al. (2013) usaron medios de cultivo para aislamientos iniciales, previo a las pruebas moleculares aplicadas a los estreptococos bioensayos.
Las técnicas de bacteriología tradicional ofrecen también herramientas que continúan siendo de gran aplicabilidad, como lo son la determinación in vitro de sensibilidad o resistencia a antimicrobianos y en ensayos de mínimas concentraciones inhibitorias (Prieto y Rodríguez, 1993; Lightner, 1996; GómezGil et al., 2015). Pese a ello, algunas patologías bacterianas se salen del rango de acción y la capacidad diferencial de estos sistemas. La bacteria causante del NHP (Vincent y Lotz, 2005; Morales-Covarrubias, 2014; OIE, 2018) y la Spiroplasma penaei (Nunan, 2004; 2005), no permiten ser cultivadas en medios artificiales generales o diferenciales, o su cultivo es muy difícil.
Otra limitante sobre las técnicas bacteriológicas surge con las especies causantes del AHPND, pese a ser cultivables en medios artificiales (Gómez-Gil et al., 2015). Nos enfrenta al dilema de no disponer de medios que permitan establecer la presencia del plásmido toxigénico, y menos aún, si están o no presentes las secuencias que codifican para las toxinas PirA y PirB. No basta identificar la especie, sino que se requiere conocer su contenido genético específico (OIE, 2018). Para el diagnóstico del AHPND se han propuesto diferentes estrategias, muchas de ellas basadas en el uso del PCR, diseñadas para detectar los genes que codifican para las toxinas (Tran et al., 2013; Joshi et al., 2014; Nunan et al., 2014), las cuales han demostrado ser sumamente sensibles y específicas. Asimismo, el PCR también es útil para diagnosticar otras patologías como el NHP (Lightner, 1996; Vincent y Lotz, 2005; Morales-Covarrubias, 2014; OIE, 2018).
La sensibilidad del análisis por las diferentes técnicas de PCR depende mucho de variables relacionadas con la ejecución de la técnica, por lo que requiere de un proceso cuidadoso de estandarización y validación del procedimiento antes de ser implementada en un laboratorio. Esto hace que la interpretación de los resultados deba ser realizada con cautela y por personal calificado.
Cuadro 1 Resumen de variables a considerar para la implementación y selección de técnicas de diagnóstico para enfermedades bacterianas en camarones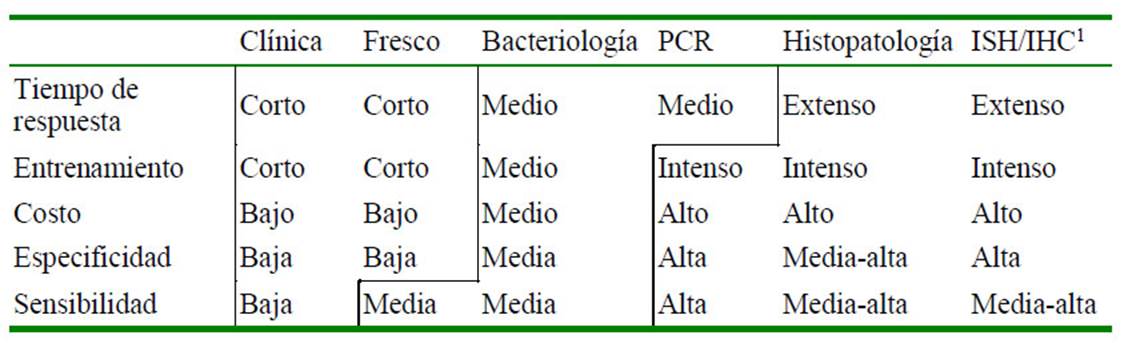
1 Hibridación in situ / Inmunohistoquímica
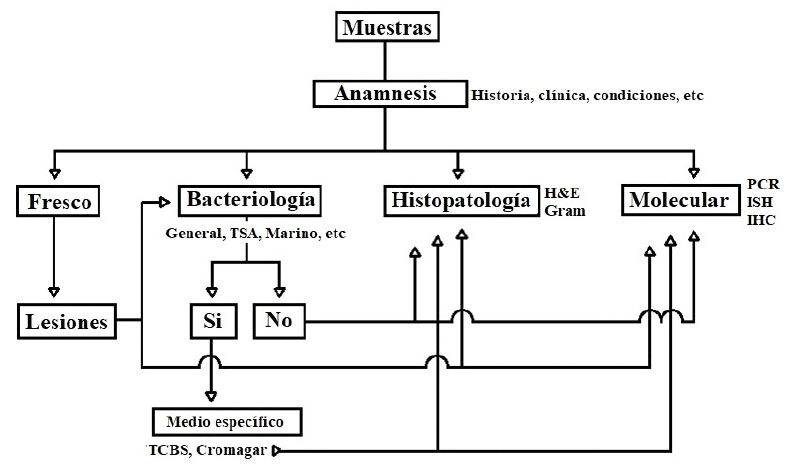
Figura 2 Flujograma general de análisis de enfermedades bacterianas en camarones. TSA: Agar tripticasa-soya; TCBS: Agar tiosulfato-citrato-bilis-sacarosa; H&E: Hematoxilina-eosina; PCR: Reacción en cadena de la polimerasa; ISH: Hibridación in situ; IHC: Inmunohistoquímica
En la actualidad, el rápido desarrollo tecnológico en el campo diagnóstico ha permitido la aparición de diferentes variantes de esta técnica que se fundamentan en la detección de ácidos nucleicos, ya sean ADN o ARN haciéndose cada vez más accesible y permitiendo la obtención de resultados preliminares incluso a nivel de campo con pruebas amigables fáciles de aplicar e interpretar (Minardi et al. 2019), por lo que en los últimos años se ha vuelto muy flexible para ser aplicada a diferentes condiciones, según sea el tipo de patógeno que se desee diagnosticar o si únicamente se busca una interpretación cualitativa o cuantitativa (Méndez-Álvarez y Pérez-Roth, 2004; Minardi et al. 2018).
Existen otras herramientas de diagnóstico molecular que complementan a la técnica de PCR, como lo son el uso de enzimas de restricción, el uso de fragmentos de ADN polimórficos aleatorios (RAPD, por sus siglas en inglés), útiles en diagnóstico de patógenos y en el desarrollo de cebadores (primers); otras técnicas como LAMP que amplifica ácidos nucleicos por un método isotérmico sin utilizar el clásico termociclador de PCR (Kongrueng et al., 2015; VarelaMejías et al., 2017; Chakraborty et al., 2019); FISH o Hibridación fluorescente in situ que detecta ARN o ADN de patógenos a partir de hibridación del ácido nucleico diana con sondas fluorescentes en diferentes tejidos; MPP o Sondas de candado molecular, que se conforman por dos cadenas de oligonucleótidos que se fijan a una secuencia de ácidos nucleicos diana, a la que una vez unidas, se cierran en forma circular para el reconocimiento específico de fragmentos que pueden contener polimorfismos de un solo nucleótido (SNP's) o bien, fragmentos de ADN de diferentes patógenos mediante pruebas multiplex. Otra técnica muy útil en la actualidad es el uso de microarrays, que tiene como ventaja la detección de múltiples patógenos de forma más eficiente que un PCR multiplex por medio de sondas marcadas con fluorescencia. Una ventaja adicional de esta técnica es el uso de pequeñas secuencias de ácidos nucleicos por lo que es posible obtener resultados, aún con ADN degradado (Chakraborty et al. 2019).
En el diagnóstico molecular, un resultado positivo indica que el patógeno se encuentra presente; es decir, se detecta la presencia de ácidos nucleicos (un fragmento del genoma) del agente infeccioso de interés, pero como en cualquier otra prueba diagnóstica, no se garantiza necesariamente que exista evidencia clínica de enfermedad. Esto es muy importante para el diagnóstico precoz de enfermedades, ya que permite acciones encaminadas a la mitigación de signos clínicos, antes que el agente infeccioso estimule la respuesta inmune que desencadena los procesos patológicos (Pérez-Roth y Méndez-Álvarez, 2004).
Es en estos casos donde las técnicas histopatológicas presentan gran valor. La presencia de lesiones permite establecer el estado de infección y el grado de compromiso tisular. Asimismo, las lesiones causadas por algunas patologías han sido claramente caracterizadas (Lightner, 1996; Pantoja y Lightner, 2014). Además, mediante estas técnicas es factible realizar diagnósticos diferenciales (Pantoja y Lightner, 2014; VarelaMejías y Peña-Navarro, 2016; Varela-Mejías, 2018).
Entre las ventajas de la histopatología se encuentra la posibilidad de establecer la identidad de los patógenos presentes, las coinfecciones y el estado sanitario general de los animales. Desafortunadamente, la técnica no es infalible, pues puede darse infecciones latentes o iniciales, en las cuales el daño celular aun no es evidente, o en el extremo contrario, infecciones terminales de AHPND, las cuales son difíciles de distinguir de infecciones por otras cepas no causantes de AHPND. Las observaciones histopatológicas brindan especificidad, pero su sensibilidad es limitada. Requiere, además de tiempo y personal altamente calificado (Pantoja y Lightner, 2014). Por otro lado, algunas patologías son difíciles de distinguir entre ellas; así, la estreptococosis, la espiroplasmosis y la vibriosis sistémica poseen grandes similitudes anatomopatológicos y tropismos celulares comunes (Lightner, 1996; Vincent y Lotz, 2005; Hasson et al., 2009; Heres et al., 2011; OIE, 2018), de modo que se requiere combinar técnicas adicionales.
Cuadro 2 Grado de confianza de las diferentes técnicas diagnósticas.
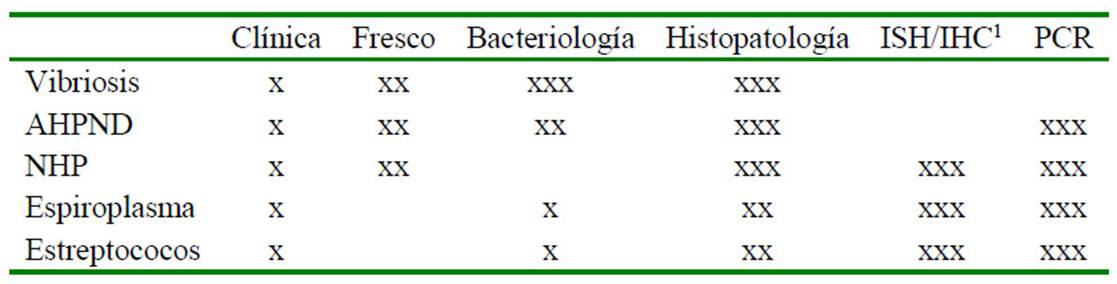
x: aplicabilidad limitada; xx; aplicabilidad moderada; xxx; aplicabilidad elevada
ISH: Hibridación in situ; IHC: Inmunohistoquímica; PCR: Reacción en cadena de la polimerasa
AHPND: Necrosis aguda del hepatopáncreas; NHP: Hepatopancreatitis necrotizante
Algunas de las limitantes de la histopatología son compartidos con las técnicas de hibridación in situ o inmunohistoquímica. En ellos, si la presencia del patógeno no es lo suficientemente alta, es posible que no sea posible observar su reacción con las sondas usadas, pese a estar presente. Su costo, tiempo de obtención de resultados y necesidad de personal calificado son también considerables. Presenta sin embargo, una ventaja sobre la histopatología en lo referente a la especificidad de los resultados (Cuéllar-Anjel, 2018; Lightner, 1996).
Con esta panorámica general, es claro que todos los métodos de diagnóstico poseen limitantes, su aplicabilidad está en función al patógeno, a sus características y al tipo de daño que causan en los camarones. El Cuadro 1 presenta un resumen de las principales variables a considerar para la implementación de las diferentes técnicas en un laboratorio de diagnóstico.
No es posible seleccionar una de estas técnicas excluyendo a las demás. Las técnicas desarrolladas incrementan su confiabilidad si se utilizan en forma complementaria, de allí que conviene combinarlas según el caso particular. La experiencia del analista es fundamental para seleccionar la técnica o técnicas adecuadas para cada caso. Pese a no existir rutas rígidas o infalibles de diagnóstico, es posible desarrollar, a modo de abordaje general, pasos secuenciales de análisis, tal como se presentada en la Figura 2. Este flujograma es orientativo y pretende sugerir rutas sobre las técnicas a seguir durante los análisis; no obstante, puede ser modificada en función a la información disponible del caso y a la experiencia del analista.
En el Cuadro 2 se presentan las principales técnicas recomendadas para las patologías descritas en este documento. Se debe considerar que la obtención de un diagnóstico confiable supone del uso de al menos dos de estas técnicas correctamente aplicadas con resultados concordantes. Finalmente, se debe tener claro que estas enfermedades no son excluyentes entre ellas, y la presencia de una de ellas, no implica la ausencia de las demás. Es posible por tanto la ocurrencia de coinfecciones, tanto entre ellas como con otras patologías.
CONCLUSIONES
Las enfermedades bacterianas poseen una gran importancia en la camaronicultura, tanto por la variabilidad de agentes causales, como por el impacto que generan sobre los cultivos.
Se dispone de una variedad de técnicas diagnósticas, cada una con diferentes grados y rangos de aplicabilidad y efectividad.
La sensibilidad y especificidad de las técnicas diagnósticas es variable; sin embargo, la combinación de dos o más de ellas mejora las posibilidades de obtener un diagnóstico certero.
Cada una de las técnicas requiere de un entrenamiento adecuado y de equipos apropiados. La selección de las técnicas a usar dependerá del caso particular, de las posibilidades y recursos del laboratorio y del tiempo disponible para su procesamiento.
Las enfermedades infecciosas en los camarones no son excluyentes entre ellas. La detección de un agente no implica ausencia de los demás

