











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La población de ganado vacuno en la provincia de Chimborazo, Ecuador, para el año 2019 fue de 315 498 cabezas, lo que representa el 7.33% a nivel nacional y 14.2% en la región Sierra, ocupando el segundo lugar en este censo en la mencionada región. En esta provincia, la población de bovinos mestizos son los más frecuentes con 93 539 cabezas, seguidos de animales de raza Holstein, Pardo Suizo y Jersey (INEC, 2020). En la zona andina de Ecuador, las condiciones climáticas, edáficas y alimentarias son mayores para los genotipos más exigentes en sistemas de pastoreo, por lo que la elección de la genética adecuada para la zona de mayor potencial lechero de la Sierra de Ecuador constituye un problema práctico (Lascano et al., 2017; Arcos et al., 2018).
La producción diaria de leche en la provincia de Chimborazo en 2019 fue de 787 108 kg de leche (10.2% a nivel nacional) (INEC, 2020). El precio por litro de leche al productor en Ecuador está indexado al precio de la venta de leche pasteurizada Ultra High Temperature (UHT), de allí que se el productor recibe 0.42 USD por litro, pudiendo alcanzar hasta 0.50 USD por litro por bonificaciones de calidad según el Acuerdo Ministerial 394 (MAGAP, 2018).
Las unidades productivas lecheras del Ecuador se caracterizan por una vida útil de 12 años, empezando a producir las hembras bovinas a los dos años y con peso al primer servicio de 250 kg (Chávez y Gavilánez, 2019). La mayoría de las ganaderías carecen de registros genealógicos y productivos adecuados, lo que dificulta la estimación de los parámetros genéticos, restringiendo el avance genético (Larrea et al., 2019).
Se requiere utilizar modelos matemáticos que permitan separar los efectos genéticos de los no genéticos para realizar la valoración (Martínez et al., 2012). La obtención de valores genéticos predichos, también conocido como evaluación genética, permite la identificación de los mejores animales, con objeto de que sean utilizados como padres de la siguiente generación, por lo que se usa el mejor predictor lineal insesgado (BLUP Best Linear Unbiased Predictor) (Valencia et al., 2003; Muñoz y Gonzáles, 2016). Los valores genéticos se calculan simultáneamente para vacas y toros usando el programa denominado modelo animal, el cual permite calificar a los animales por su mérito genético y estimar las diferencias genéticas entre animales (Salinas et al., 2016), por medio de la estimación de habilidades de transmisión predichas (Holstein Foundation, 2018). También permite estimar de forma simultánea los efectos ambientales y genéticos que producen la observación (Meza et al., 2017).
En la actualidad existen métodos que permiten la evaluación genética de los animales en producción, de los que están por entrar en actividad productiva y aún de aquellos que no cuentan con registros de producción (Martínez et al., 2017). El modelo animal permite que todas las relaciones genéticas entre los animales contribuyan a la conectividad de los datos, aunque la calidad de las evaluaciones genéticas depende de muchos factores, en particular de la estructura de la información (Magaña et al., 2013); adicionalmente, este modelo actúa de tal forma que considera que los animales sin progenitores pertenecen a la generación base y se supone que están extraídos al azar de una población de media cero y de varianza genética que tenían los animales fundadores de la población (Quijano y Montoya, 1998).
En los cruzamientos, los productores tienen como objetivo explotar las características favorables de las razas alternativas, eliminar los efectos negativos asociados con la endogamia y sacar provecho de la heterosis (Quinteros y Marini, 2017). Los cruzamientos y la selección son las únicas herramientas disponibles para la mejora animal, permitiendo aumentar la productividad rápidamente (Madalena, 2001). La estimación de los efectos de cruzamiento permite predecir el comportamiento productivo esperado de diferentes cruzas y establecer esquemas de evaluación genética para seleccionar a los reproductores a través de su valor genético aditivo, sin los sesgos ocasionados por los efectos de heterosis y recombinación en poblaciones multirraciales (Román et al., 2013).
Dado que las diferencias en el mérito genético permiten incorporar al hato animales seleccionados por su alto valor genético para las características económicas productivas que se desean mejorar, en función de las decisiones que considere cada criador (Larrea et al., 2019), se contempló como objetivo estimar los parámetros genéticos (heredabilidad y repetibilidad) y predecir el mérito genético para la producción de leche ajustada a 305 días (PL) de los bovinos de un hato lechero en la provincia de Chimborazo, para ser utilizados como criterio de selección del hato.
MATERIALES Y MÉTODOS
Área del Estudio y Animales
La investigación se realizó en la hacienda «La Isabela de Sasapud», ubicada en la parroquia Matriz, cantón Chambo, provincia de Chimborazo, Ecuador. La zona se sitúa en el piso climático premontano alto a 3340 msnm, con una pluviosidad anual de 1031 mm y temperatura promedio anual de 12.6 °C. La hacienda posee 61 ha, siendo 36 ha de bosque primario montano alto con topografía superior al 40% de inclinación y las restantes 25 ha se usan para potreros compuestos principalmente de mezclas forrajeras de ryegrass (Lolium perenne), trébol blanco (Trifolium albus) y llantén forrajero (Plantago major) con intervalo de pastoreo de 32 días.
Se analizó el registro genealógico y productivo de 289 animales con 349 lactancias entre los años 2000 y 2019, provenientes de la base de datos que registra la explotación (Cuadro 1). Se consideró la genealogía, las 18 composiciones raciales entre puras y cruzadas entre sí de Holstein (H), Jersey (J), Sueco rojo (S) y Montbéliarde (M), el número de lactancias por vaca (1-9), el año de inicio de la lactancia (2000-2019), y como variable dependiente la PL. Se eliminaron 12 registros ajustados por presentar producciones menores a 1000 kg de leche, por lo que se consideraron 337 lactancias para este estudio.
Cuadro 1 Distribución de animales y lactancias por composición racial
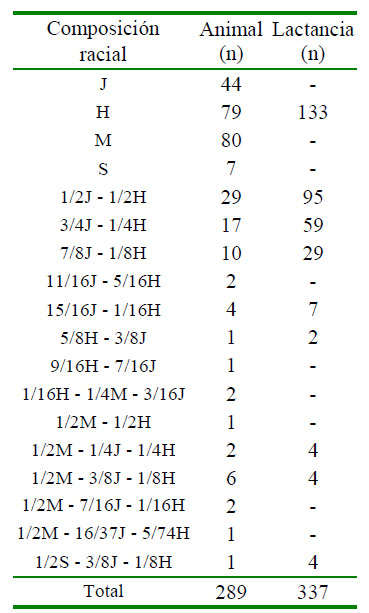
H = Holstein; J = Jersey; S = Sueco rojo; M= Montbéliarde
Animales de algunas razas y cruces no poseían información de producción, por ser animales fundadores, introducidos desde otras explotaciones o por no haber iniciado su vida productiva. Sin embargo, fueron considerados para establecer el coeficiente de consanguinidad de cada animal e incluirlo en el modelo. La explotación en un principio utilizó toros de raza Jersey para monta directa, posteriormente introdujo la inseminación artificial (IA) utilizando semen congelado de toros Holstein, Jersey, Sueco rojo y Montbeliarde, manejando un sistema de reproducción mixto (monta directa e inseminación artificial). Se procedía con la monta a las hembras que retornaban en celo después de la segunda inseminación. Desde 2012 solo se emplea IA con semen de toros Holstein, Jersey y Montbéliarde manteniendo un cruce rotacional trihíbrido.
Ajuste de Lactancia
Se utilizó la función gama incompleta propuesta por Woods en 1967, por la bondad de ajuste a los datos de producción de leche y la capacidad predictiva de los rendimientos de leche por lactancia siguiendo la metodología aplicada por Gebreyohannes (2013) y Vargas (2016). Se aplicó el modelo lineal general (PROC GLM) en SAS v 9.4 (2013), y los resultados se obtuvieron del análisis de regresión polinómica de tercer orden, cuya función que está representada por PL = atbe-ct, donde PL = Producción de leche ajustada a 305 días (kg); t = Periodo de tiempo en el que se estimó la producción láctea (1-305 días); a, b, c = Parámetros de la función gamma incompleta; e = Base de los logaritmos naturales.
Aplicando el logaritmo natural a la función gama incompleta, se obtiene: Ln(Y) = ln(a) + bLn(t)-ct, donde Ln = Logaritmo natural; Y = Producción de leche (kg); t = Periodo de tiempo en el que se estimó la producción láctea (1-305 días); a, b, c = Parámetros de la función gamma incompleta.
Análisis Estadístico
En el modelo se consideró los efectos aleatorios genético aditivo, ambiente permanente y residual (ambiente temporal) y dentro de los efectos fijos se incluyó la composición racial, número de lactancia y año de inicio de cada lactancia de los individuos en estudio, usando el siguiente modelo: Yijklm = μ + Ai + Bj + Ck +Dl +Em + eijklm, donde Yijklm = Vector de los registros de producción de leche ajustada a 305 días (kg); μ = Promedio general de la leche ajustada a 305 días (kg); Ai = Efecto aleatorio del i-ésimo efecto genético aditivo; Bj = Efecto fijo de la j-ésima composición racial, j=1…18; Ck = Efecto fijo de la k-ésima lactancia, k=1…10; Dl = Efecto fijo del l-ésimo año de inicio la producción de leche, l=1…21; Em = Efecto aleatorio del m-ésimo efecto del ambiente permanente; y eijklm = Efecto residual.
El análisis de la varianza para los efectos fijos que intervienen en la PL fue realizado mediante PROC GLM del paquete estadístico SAS v.9.4 (2013). En caso de encontrar diferencia significativa dentro de cada efecto fijo, se procedió a separar las medias con el test de Duncan.
Los componentes de la varianza requeridos para la estimación de los parámetros genéticos y valores de cría, fueron obtenidos por el método de máxima verosimilitud restringida, empleando el programa VCE v. 6.0 (Groeneveld et al., 2008), obteniendo la heredabilidad (h2) y la repetibilidad (r) de la PL. Para estimar el mérito genético y el coeficiente de consanguinidad de cada animal se corrieron los registros en el programa MTDFRELM (Boldman et al., 1995). El modelo incluyó en ambos programas los efectos fijos de composición racial, número de lactancia y año de inicio de lactancia, y como efectos aleatorios los animales analizados, el ambiente permanente y el residuo (ambiente temporal). El modelo empleado en notación matricial fue: Y = xb + Za + Zp + e, donde Y = Vector de los registros de producción de leche ajustada a 305 días (kg); a = Vector de los efectos aleatorios genéticos aditivos directos; b = Vector de los efectos fijos incluyendo los grupos contemporáneos definidos por la combinación de la composición racial, lactancia y año de inicio de lactancia; p = Vector del efecto ambiental permanente; X, Z = Matrices de incidencia que relacionan las observaciones en Y con los respectivos efectos; y e = Vector de residuales aleatorios.
Una vez estimado el valor de cría correspondiente para cada animal, se realizó una distribución de frecuencias de nueve clases aplicando la metodología propuesta por Larrea et al., (2019), con la finalidad de clasificar los animales de las diferentes categorías de acuerdo con su valor genético, para así determinar el grupo genético élite de la explotación.
RESULTADOS Y DISCUSIÓN
Consanguinidad
Se encontró 11 animales con endogamia, cuyos coeficientes fluctuaron entre 0.004 a 0.250 con promedio de 0.107 (± 0.115); los cruces raciales que presentaron consanguinidad fueron 3/4J-1/4H, 7/8J-1/8H, 15/16J-1/16H, distribuidos en los siguientes valores relativos de 63.6, 27.3 y 9.1% de los animales consanguíneos, respectivamente. En general, se reconoce que la endogamia afecta negativamente a la producción de leche y la supervivencia, siendo así que la depresión endogámica puede causar importantes pérdidas económicas para los productores lecheros (Domínguez et al., 2014; Andere et al., 2017).
Desempeño de la Producción de Leche Ajustada a 305 Días (PL)
El valor promedio de la PL fue de 2972.1 ± 51.1 kg, con rangos entre 1053.0 y 5843 kg. Se encontró diferencias significativas en el efecto fijo de la composición racial (p<0.05) y por número de lactancia y año (p<0.01).
Parámetros Genéticos
Los estimados de componentes de la varianza para la PL fueron: varianza aditiva (σ2 ) 159 112, varianza del ambiente permanente (σ2 ) 148 075, varianza residual (σ2 ) 300 389 y varianza fenotípica (σ2 ) 607 576, con los que se obtuvieron una h2 de 0.26 ± 0.16 y una r de 0.51 ± 0.15. El efecto genético aditivo para PL fue moderadamente heredable, lo que indica que con este estimador es factible aumentar la producción de leche por lactancia en el hato lechero en estudio, a través de la selección de animales sobresalientes (Ríos at al., 2012), a la vez que el valor de repetibilidad se debió a que la σ2 fue menor que la σ2 , lo que indica baja variación genética y la ocasionada por el ambiente permanente en relación con la variación ambiental temporal para el rasgo estudiado.
La h2 calculada fue superior a los valores reportados en ganado Holstein por Hidalgo (2019) con 0.16 ± 0.03 en Perú, Salem y Hammoud (2016) con 0.15 ± 0.05 en Egipto; así como por Cañas et al. (2012) con 0.17, Arango y Echeverri (2014) con 0.19 y Zambrano et al. (2014) con 0.24 ± 0.08 en Colombia. Con respecto al grupo racial, en ganado Jersey, Arango y Echeverri (2014) reportaron una heredabilidad de 0.20 y Zambrano et al (2014) de 0.34 ± 0.30, ambos en Colombia y Okeke et al., (2019) en Nigeria con 0.35; mientras que Román (2006) reportó 0.12 ± 0.04 en cruces multirraciales en un sistema de doble propósito en el trópico mexicano y Ríos at al. (2012) con 0.28 ± 0.08 en cruce Holstein y Pardo suizo.
La r en la población estudiada para PL fue superior a los valores reportados en ganado Holstein por Hidalgo (2019) con 0.28 ± 0.03 en Perú, Salem y Hammoud (2016) con 0.19 ± 0.03 en Egipto, Toledo et al. (2014) con 0.41 ± 0.04 en México. Asimismo, por Frioni (2012) con 0.45 ± 0.002 en bovinos Holando Uruguayo, Zambrano et al. (2014) con 0.34 ± 0.30 en Jersey en Colombia, Román (2006) con 0.34 ± 0.02 en cruces multirraciales en el trópico mexicano y Ríos et al. (2012) con 0.40 en cruce Holstein x Pardo Suizo. El resultado obtenido solo fue inferior al reporte de Okeke et al. (2019) en Jersey en Nigeria con 0.57.
Mérito Genético por Animal
En el Cuadro 2 se observa que, al clasificar los valores genéticos predichos en nueve clases con intervalo de 56.43 kg de PL, la mayor frecuencia (52.94%) corresponde a los animales de la clase incierta con valores genéticos entre -15.71 a +40.72 kg de PL, dentro de los cuales se incluyen los animales con valores genéticos predicho cero. Se observa además que 23.87% de los animales presentó valores genéticos positivos que corresponde entre las clases ligeramente alto y muy alto, los mismos que pueden ser considerados como animales élite para el desarrollo genético futuro de la explotación.
Cuadro 2 Distribución de las clases de las vacas de acuerdo con el mérito genético en la población estudiada
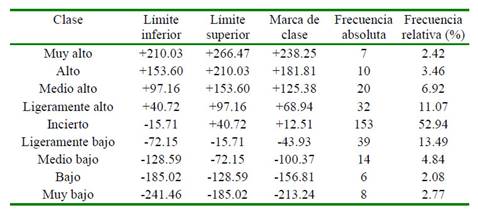
Las frecuencias absolutas y relativas de los valores genéticos predichos para los animales evaluados variaron entre -241.46 a +266.47 kg de PL (Cuadro 2), distribuyéndose el 49.48% con valores positivos, 31.41% negativos y 19.38% con valor cero. Este último se justifica por el número de animales que no presentaron registros de producción y no poseían conexión de parentesco con animales que presentaban registros o eran fundadores. Los animales que se encuentran actualmente en el inventario de la explotación poseen valores genéticos entre -129.64 a +225.29 kg de PL, con un alto porcentaje de animales con valor genético positivo (58.54%) y en menor proporción los animales con valor genético negativo y cero (29.26 y 12.20%, respectivamente).
Desempeño por Composición Racial
El mayor desempeño productivo se encontró en los animales de composición racial trihíbrida: (1/2M-1/4J-1/4H, 1/2M-3/8J-1/8H y 1/2S-3/8J-1/8H), seguidos por los animales de composición racial dihíbrida (3/4J-1/4H, 7/8J-1/8H, 1/2J-1/2H y 5/8H-3/8J) a excepción de los animales con mestizaje 15/16J-1/16H, siendo este último cruce el que presentó menor rendimiento y difirió con las demás composiciones. De otra parte, los animales Holstein, quienes tuvieron la mayor cantidad de registros, obtuvieron un rendimiento promedio de 2795.7 ± 82.5 kg (Cuadro 3). De la misma forma, los valores genéticos tuvieron un comportamiento similar al orden de los promedios de PL, destacándose los cruces 1/2M-3/8J-1/8H, 1/2M-1/4J-1/4H y 1/2S-3/8J-1/8H con +1228.1, + 1106.9 y +1044.7 kg de PL, respectivamente.
Cuadro 3 Valor genético1 de la producción de leche ajustada a 305 fías con respecto a la composición racial
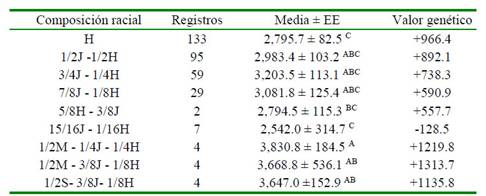
1 Valor de probabilidad = 0.0232
J = Jersey; H = Holstein; M = Montbéliarde; S = Sueco rojo EE = Error estándar
Letras diferentes dentro de la misma columna indican diferencias significativas (p<0.05)
Cuadro 4 Valor genético1 de la producción de leche ajustada a 305 días con respecto al número de lactancias
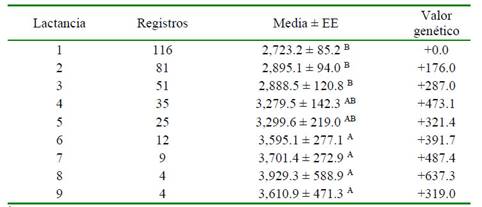
1 Valor de probabilidad = 0.0007 EE = Error estándar
A,B Letras diferentes dentro de la misma columna indican diferencias significativas (p<0.01)
La principal ventaja de realizar cruzamientos es la heterosis, que permite introducir caracteres deseables en una población en la que no han existido anteriormente, lo que conduce a un aumento en el nivel genético en una descendencia híbrida en comparación con el promedio de las razas parentales (Kumar et al., 2016).
Los sistemas de cruzamiento mediante la incorporación de semen, embriones y sementales Bos taurus importados para uso en hembras locales es una herramienta de mejoramiento genético de importancia (Román, 2006; Toalombo et al., 2019). Es difícil mejorar los rasgos de baja heredabilidad como la fertilidad, rendimiento de leche y longevidad a través de la reproducción dentro de una raza pura, pero se mejoran en gran medida a través del cruzamiento (Kumar et al., 2016).
Desempeño por Lactancia
En la Cuadro 5 se observa que los mayores promedios corresponden a los animales con mayor número de lactancias (>4), con medias de PL entre 3279.5 ± 142.3 y 3929.3 ± 588.9, valores que siguen un patrón ascendente hasta la octava lactancia. Al analizar el valor genético de cada lactancia, se observa que siguen un patrón diferente al de los promedios, descendiendo en la quinta y con ascenso a partir de la sexta a la octava lactancia, donde se encontró el valor máximo (+632.8), para volver a descender en la novena campaña.
En número de lactancias, la edad a disposición o el último parto y la supervivencia a una edad fija se han incluido en las medidas de longevidad (Miglior et al., 2017). Se ha demostrado que el cruzamiento mejora la longevidad del ganado lechero; sin embargo, se reconoce que el potencial de producción total de leche de la vaca se obtiene a la tercera o cuarta lactancia, suponiendo que es cuando la vaca puede ser más rentable; por lo tanto, a medida que la vaca envejece, los rasgos funcionales pueden deteriorarse, y en muchos casos la vaca será sacrificada antes de que alcance su potencial de producción de leche (Clasen et al., 2017).
La longevidad ha adquirido importancia como indicador global del bienestar animal. Los estudios indican que la longevidad es heredable, habiéndose incluido en las evaluaciones genéticas de rutina (De Mello et al., 2014; Heise et al., 2016). Hernández et al. (2011) plantearon que cuando las lactancias posteriores a la primera lactancia eran incluidas en la estimación del valor genético, se creaban más conexiones entre sementales permitiendo incrementar la exactitud de las evaluaciones.
Desempeño por Año de Inicio de Lactancia
Los mayores promedios por lactancia correspondieron a los años 2013 y 2006 con 3932.6 ± 303.9 y 3732.8 ± 184.4, respectivamente. El mérito genético para el año de inicio de lactancia sigue las tendencias de los promedios de PL por año, las mismas que se vieron influenciadas por lo cambios en el manejo y por los efectos climáticos de alta precipitación que se dan en la zona, lo cual no permite mantener un normal desenvolvimiento productivo, ya que los pastizales se ven afectados por los excesos de precipitación (Cuadro 5).
Cuadro 5 Valor genético1 de la producción de leche con respecto al año de inicio de la lactancia
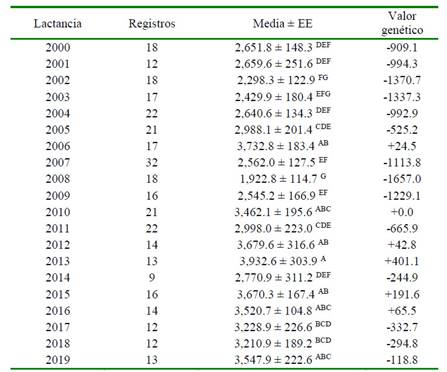
1 Valor de probabilidad = 0.0001 EE = Error estándar
A,B,C,D,F Letras diferentes dentro de la misma columna indican diferencias significativas (p<0.01)
El mayor mérito genético se obtuvo en el año 2013 con +401.1 kg de PL. Se asume que al incrementar el número de animales trihíbridos aumentará la producción de leche, aprovechando el efecto de la heterosis.
CONCLUSIONES
Los estimados de heredabilidad y repetibilidad para la producción de leche ajustada a 305 días (PL) son similares a los reportados en la literatura científica.
Los efectos fijos de composición racial, número de lactancia y año de inicio de lactancia, mostraron fueron significativos para la PL, destacándose los cruces trihíbridos (1/2M-3/8J-1/8H, 1/2M-1/4J1/4H y 1/2S-3/8J-1/8H) por efecto de la heterosis.
Los animales con mayor número de lactancias tienen mayor mérito genético.

