











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
La Tricología es una ciencia que emplea métodos no invasivos para la identificación de los mamíferos en base al análisis de la morfología del pelo, mediante la combinación de las principales características de la medula, corteza y cutícula (Quadros y Monteiro-Filho, 1998).
Teerink (1991) clasifica a los pelos en dos clases: (1) Pelos de cobertura o «underhair», los cuales funcionan como una capa que atrapa aire y aísla al animal de la intemperie. Dichos pelos son poco pigmentados, numerosos, ondulados y con constricciones. Se encuentran tres tipos de pelos de cobertura: lana (pelos largos, suaves y rizados), pelaje (pelos cortos, finos y abundantes) y vello (pelos finos y cortos) (Martin et al., 2001); y Pelos de guardia o «guard hair», los cuales ayudan a definir patrones de coloración, protegen los pelos de cobertura y repelen el agua. Estos pelos son largos, gruesos, fuertemente pigmentados y sobresalen por encima del resto de pelos (Teerink, 1991; Fernández y Rossi 1998; Monroy-Vilchis et al., 2005). El pelo sufre notables modificaciones en algunos grupos de mamíferos, pudiendo constituirse como cerdas, espinas o placas (Cabrera, 1922), mientras que en la gran mayoría de grupos se presenta un pelo suave al tacto y sin mayores cambios estructurales macroscópicos y/o microscópicos. No obstante, el pelo de los murciélagos no presenta médula, característica única dentro del grupo de los mamíferos (Sessions et al., 2014).
El estudio de pelos en murciélagos se remonta a los mediados del Siglo XX, donde se describe y utiliza a los pelos como potenciales caracteres taxonómicos para distinguir entre especies de Norte América (Mathiak, 1938; Nason, 1948; Benedict, 1957). Posteriormente, la iniciativa se aplicó en algunas familias de murciélagos utilizando nuevos enfoques metodológicos en Sudamérica (Van Staaden y Jones, 1997), Centro América (Baca y Sánchez-Cordero, 2004; Schaetz et al., 2009), Norte América (Moore y Braun, 1983; Amman et al., 2002; Debelica y Thies, 2009) e India (Kumar et al., 2014).
Si bien, algunos autores consideran que la estructura del pelo posee un valor taxonómico limitado en murciélagos (Nason, 1948; Benedict, 1957), otros los han utilizado ampliamente para construir claves taxonómicas en base a los patrones cuticulares de las escamas, forma de la célula cortical y patrones de pigmentación (Mathiak, 1938; Moore y Braun, 1983). Por tanto, considerando el potencial de la microestructura del pelo como carácter diagnóstico y la escasa información sobre pelos de murciélagos en Sudamérica (Benedict, 1957; Van Staaden y Jones, 1997), el presente estudio tuvo como objetivo describir la microestructura cuticular de los pelos de guardia de los murciélagos presentes en el desierto costero del sur de Perú.
MATERIALES Y MÉTODOS
Quadros y Monteiro-Filho (1998) observaron que los procedimientos de taxidermia no dañan o alteran los patrones medulares y cuticulares de los pelos, lo cual permitió utilizar pelos de colecciones científicas de Museos de Historia Natural en este estudio como material de referencia para la identificación de las especies de mamíferos que se encuentran en egagrópilas, excrementos, etc. Por consiguiente, se examinaron pelos de 16 de las 18 especies de murciélagos reportadas para el Departamento de Arequipa, Perú (Pari et al., 2015; Medina et al., 2018; Málaga et al., 2020), los cuales constituyen una muestra representativa del ensamble de murciélagos que ocurre en el Desierto Costero del sur de Perú (Cuadro 1).
Los pelos fueron obtenidos de 45 especímenes depositados en la colección científica de mamíferos del Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa (MUSA), los cuales se agruparon taxonómicamente en 16 especies, 13 géneros y 4 familias. Tres individuos por especie fueron seleccionados al azar para colectar 10 muestras de pelo de la zona interescapular de cada murciélago, siguiendo la metodología de Arita y Aranda (1987) y Quadros y Monteiro-Filho (2006). Los pelos fueron limpiados con solución jabonosa y alcohol al 70%, según Kshirsagar et al. (2009) y Fasola et al. (2005), procesándose de 20 a 30 pelos por especie.
Los pelos de los murciélagos se clasifican en tres tipos: lanosos, primarios y secundarios, en los cuales se distinguen tres regiones: la raíz, que incluye el bulbo del pelo; el tallo, que constituye la mayor longitud del pelo; y la espátula, apéndice típico con forma de lanza (Pierallini et al., 2004). Por tanto, la descripción macroscópica fue realizada en base a la coloración, número de bandas (bandeo) y largo de los pelos (LP), mientras que para la descripción microscópica se utilizó la parte distal o espátula del pelo de guardia secundario, siguiendo las definiciones y esquemas reportadas en la literatura científica (Nason, 1948; Benedict, 1957; Arita yAranda, 1987; Amman et al., 2002; Pierallini et al., 2004; Schaetz et al., 2009).
Se obtuvieron microfotografías de la cutícula, parte distal de los pelos, con ayuda de un Microscopio Electrónico de Barrido (MEB) FEI Inspect TM S50, conectado al programa xT microscope Control. Se trabajó con una magnificación de 2600x, SPOT = 4-5, HV = 3.5-10 kV, Presión = 1.06E-06 - 2.01E-05 Torr, WD = 9.6-11.4 mm, HFW = 115 µm.
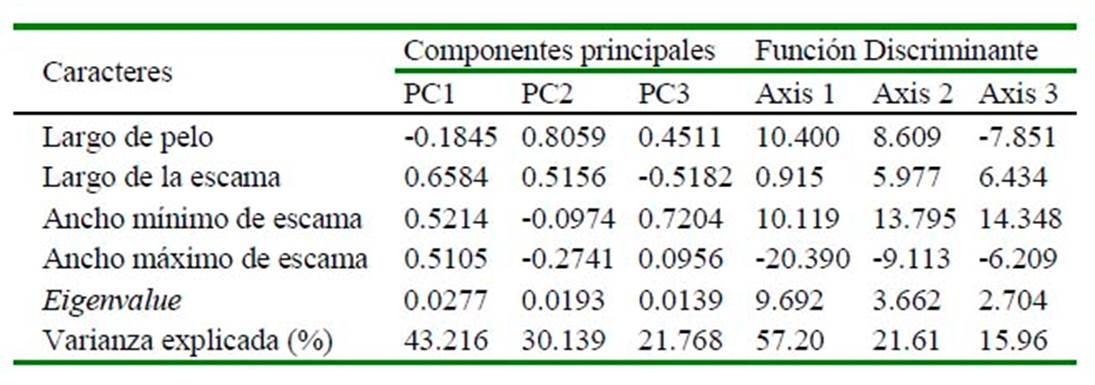
Se registraron las siguientes medidas: Largo de escama (LE), Ancho máximo de escama (AMaxE) y Ancho mínimo de escama (AMinE), las cuales permitieron estimar el Índice de Escama (IS = LE/AMaxE) y el Índice de Ancho de Escama (WI = AMaxE/ AMinE) (Debelica y Thies, 2009). Análisis de componentes principales (ACP) fueron llevados a cabo usando una matriz de varianzacovarianza en base a las cuatro medidas tomadas, las cuales fueron normalizadas usando la transformación logarítmica. Los primeros tres componentes fueron obtenidos y los pesos de todas las variables fueron evaluados como una medida de importancia en esta separación preliminar. Las diferencias estadísticas entre grupos fueron evaluadas usando análisis de función discriminante (AFD) (Brown y Wicker, 2000) y la validación cruzada fue evaluada usando la opción «leave out» en el programa PAST. Los análisis se realizaron con el software PAST 3.0 para Windows (Hammer, 2013).

RESULTADOS
Se encontraron cinco tipos de patrones cuticulares coronales, los cuales permiten diferenciar a los especímenes a nivel de familia (Cuadro 2). Los murciélagos Phyllostomidae presentan los patrones cuticulares adpreso y alternado, los Furipteridae tienen un patrón divergente, los Molossidae presentan patrones divergente y divaricado y los Vespertilionidae es de tipo hastada asimétrico (Figura 1, Figura 2).
Así mismo, se reconocieron seis configuraciones del borde de las escamas, permitiendo diferenciar fácilmente los especímenes hasta el nivel de subfamilia de la siguiente manera:
Desmodontinae (Desmodus rotundus): Escamas adpresas con bordes hendidos.
Phyllostominae (Platalina genovensium) y Glossophaginae (Glossophaga valens): Escamas alternadas con bordes simples y emarginados.
Sternodermatinae (Sturnira bogotensis): Escamas alternadas con bordes denticulados.
Tomopeatinae (Tomopeas ravus): Escamas divaricadas con bordes denticulados.
Molossinae (Molossus molossus, Nyctinomops macrotis y Promops davisoni) presentan escamas divergentes y bordes dentados. (Mormopterus kalinowskii, Nyctinomops aurispinosus, N. laticaudatus y Tadarida brasiliensis) presentan escamas divaricadas, ambas con bordes dentados.
Los Cuadro 2 y Cuadro 3 detallan los caracteres macro y microscópicos de los pelos de guardia de cada una de las especies revisadas.
Con base a las medidas tomadas (Cuadro 3), los tres primeros componentes del ACP recuperaron el 95.12% de la variación total, observándose cierto grado de agrupación entre las especies que pertenecen a una misma familia o subfamilia (Figura 3). El componente principal 1 (CP1) recuperó el mayor porcentaje de variación (43.22%), siendo el Largo de escama el carácter que presentó el mayor valor positivo (Cuadro 4). El AFD mostró que 57.2 y 21.61% de la variación es explicado por el primer y segundo eje canónico (Cuadro 4). La matriz de factores discriminantes indica que el Largo de pelo tiene una alta correlación positiva con el axis de la primera función discriminante, mientras que el Ancho mínimo de escama con la segunda función discriminante. El AFD muestra una correcta asignación de los individuos a los grupos previamente definidos; sin embargo, utilizando la prueba de Jackknifed, solo 56.88% de los casos estarían correctamente asignados.
En base a la combinación de datos morfológicos y morfométricos se desarrolló una clave dicotómica para determinar las especies de murciélagos del Desierto costero en el sur de Perú (Cuadro 5).
DISCUSIÓN
El pelo presenta resistencia a factores físicos, químicos, mecánicos y biológicos (Quadros y Monteiro-Filho, 1998), motivo por el cual es ampliamente utilizado en la industria textil, criminalística, ecología trófica y paleontología. No obstante, la falta de catálogos taxonómicos de pelos reduce la calidad de sus productos y aplicaciones.
Cuadro 2 Matriz de característica externa y patrones cuticulares murciélagos en la región Arequipa (Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa
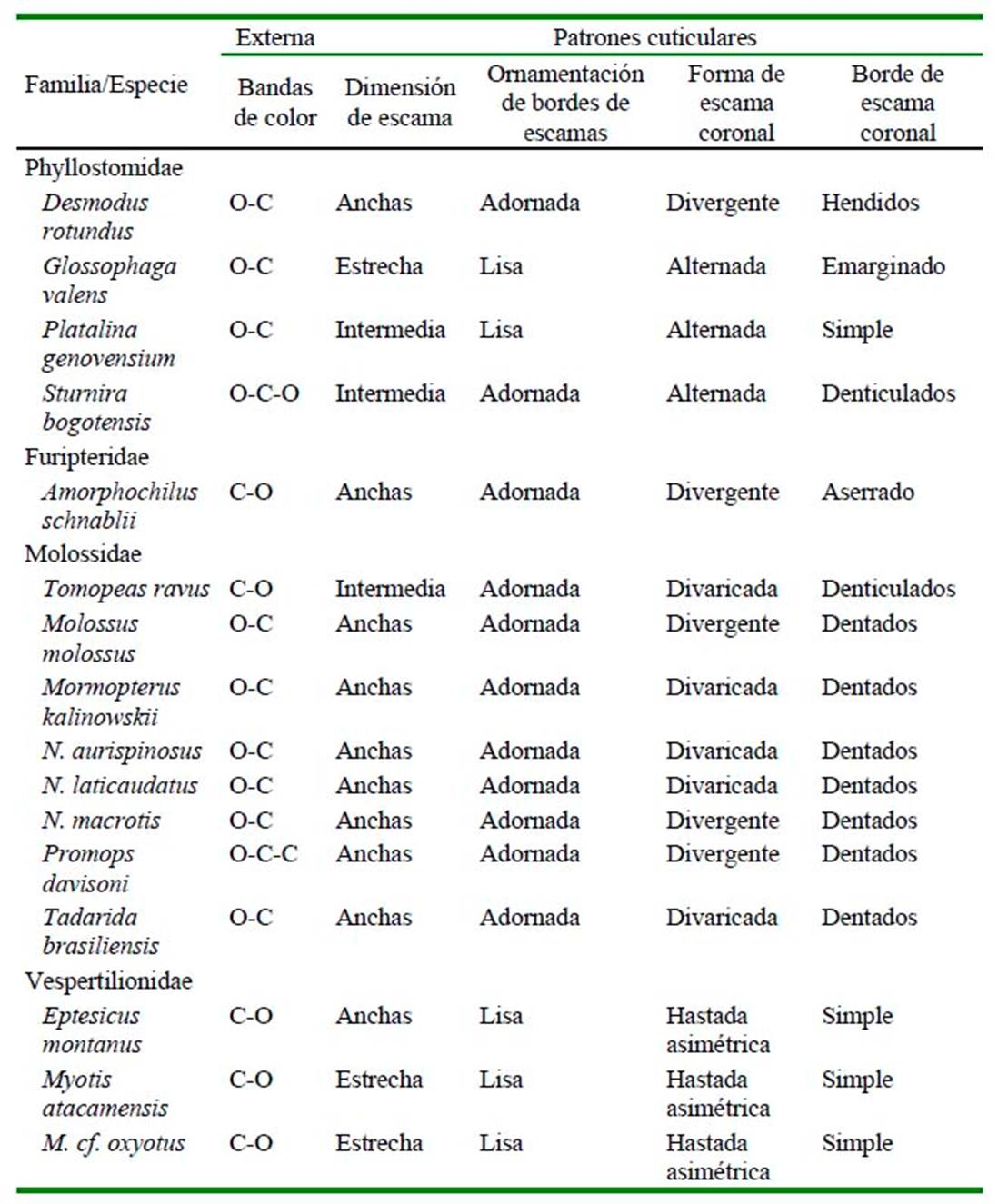
Bandas de color: O: Oscuro y C: Claro

Figura 1 Patrones cuticulares de pelos de guardia de murciélagos de las familias Phyllostomidae, Furipteridae y Vespertilionidae presentes en la Región Arequipa, Perú: A, Desmodus rotundus (MUSA 1855); B, Glossophaga valens (MUSA 16979); C, Platalina genovensium (MUSA 6425); D, Sturnira bogotensis (MUSA 11228); E, Amorphochilus schnablii (MUSA 9386); F, Eptesicus montanus (MUSA 16964); G, Myotis atacamensis (MUSA 9820); H, Myotis cf. oxyotus (MUSA 9826)

Figura 2 Patrones cuticulares de pelos de guardia de murciélagos de las familias Molossidae presentes en la Región Arequipa, Perú: A, Tomopeas ravus (MUSA 14500); B, Molossus molossus (MUSA 9687); C, Mormopterus kalinowskii (MUSA 13601); D, Nyctinomops aurispinosus (MUSA 2659); E, Nyctinomops laticaudatus (MUSA 1295); F, Nyctinomops macrotis (MUSA 11326); G, Promops davisoni (MUSA 16983); H, Tadarida brasiliensis (MUSA 13429)
Cuadro 3 Datos cuantitativos (promedio ± d.e.) de los pelos de guardia de murciélagos en la Región Arequipa (Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa)
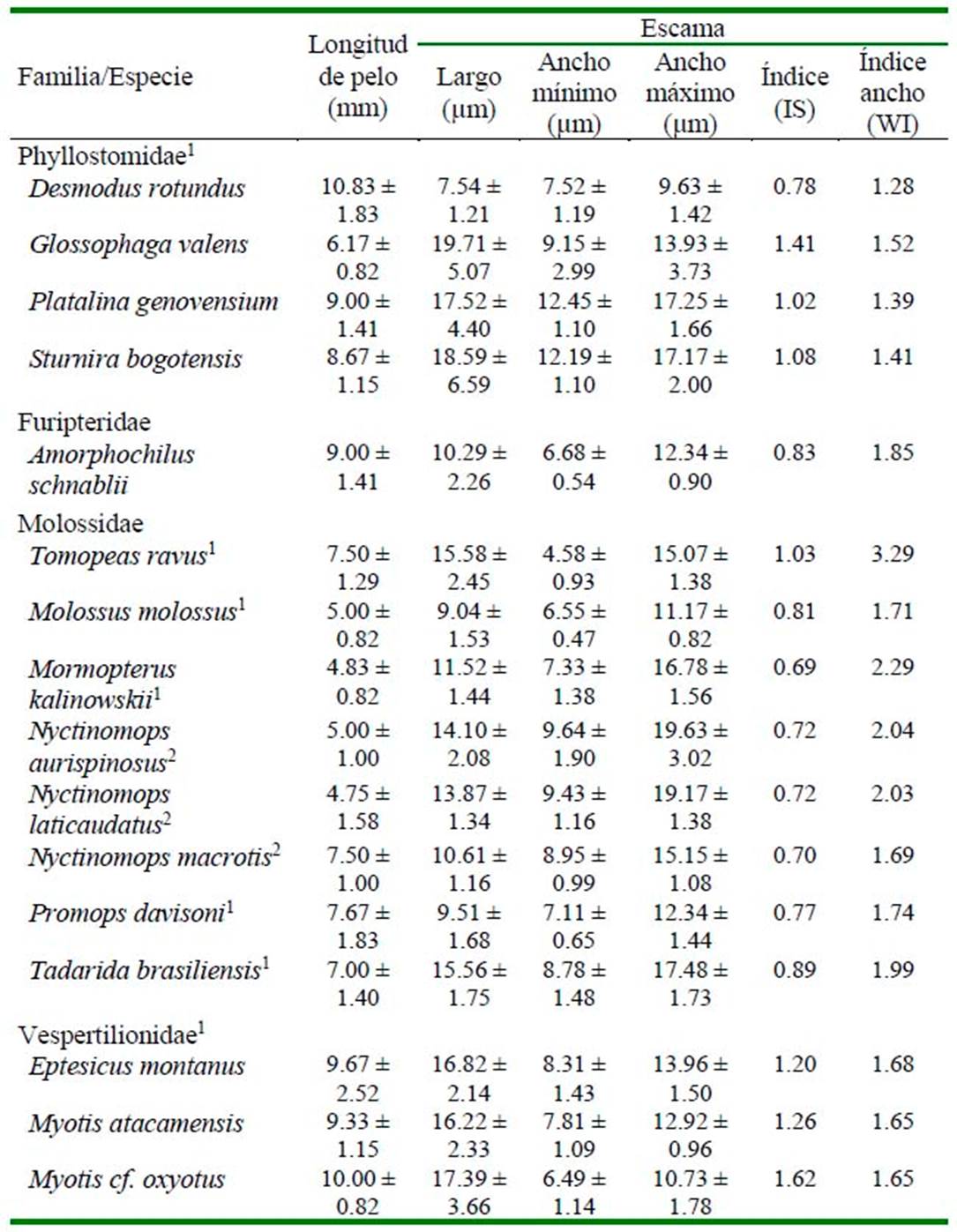
1 30 pelos por especie; 2 20 pelos por especie
Cuadro 4 Factores extraídos del análisis de componentes principales y del análisis de función discriminante para 16 especies de murciélagos del desierto costero del sur de Perú (Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa), en base a una matriz de varianza-covarianza
Información sobre la correcta identificación de los pelos de murciélagos en Sudamérica es limitada (Van Staaden y Jones, 1997). A pesar de que existen colecciones biológicas importantes de mamíferos en los museos de historia natural del Perú, sólo se cuenta con un estudio de pelos a nivel nacional (Medina et al., 2018), lo cual pone en evidencia la relevancia del presente estudio.
Los pelos observados en las cuatro familias de murciélagos estudiadas (Phyllostomidae, Furipteridae, Molossidae y Vespertilionidae) carecieron de médula, lo cual concuerda con lo observado por diversos autores (van Staaden y Jones, 1997; Baca y Sánchez-Cordero, 2004; Debelica y Thies, 2009); sin embargo, al realizar la descripción de los patrones coronales, y compararlos con descripciones previas, se observó una falta de uniformidad en las definiciones, como a continuación se detalla:
Las especies de la familia Phyllostomidae, subfamilia Desmodontinae presentaron escamas coronales adpresas hendidas, mientras que las subfamilias Glossophaginae y Sternodermatinae presentaron escamas alternadas simples y emarginadas. Benedict (1957) observó que D. rotundus tenía escamas de forma coronal corta, repanada a sinuosa con menores irregularidades marginales; sin embargo, Schaetz et al. (2009) lo describen como escamas divergentes hendidas. Respecto a las subfamilias Glossophaginae y Sternodermatinae, Baca y Sánchez-Cordero (2004) describen las escamas de G. soricina como coronal hastada asimétrica; mientras que, Schaetz et al. (2009) describen las escamas coronales como alternadas, corroborando las observaciones del presente estudio. En la familia Furipteridae, Amorphochilus schnablii presentó escamas coronales divergentes denticuladas; concordando con lo observado por Benedict (1957).
En la familia Molossidae se encontraron escamas de tipo divergentes y divaricadas con bordes dentados. Benedict (1957) encontró escamas coronales dentadas con una simple y profunda incisión en V, mientras que van Staaden y Jones (1997) indican que los murciélagos tienen escamas coronales de tipo divergente o divaricada. Tadarida brasiliensis presentó escamas más divergentes que Nyctinomops y las escamas en Nyctinomops aurispinosus son menos divergentes que las de otros miembros de ese género. Amman et al. (2002) concluyeron que las escamas tipo divaricada y divergente son características de esta familia, con excepción de los murciélagos N. macrotis y T. brasiliensis que presentan escamas coronales dentadas; Pierallini et al. (2004) describen escamas coronales divaricadas, Baca y Sánchez-Cordero (2004) mencionan que las escamas coronales dentadas son propias de T. brasiliensis, Schaetz et al. (2009) observaron escamas coronales divaricadas con más de seis dientes visibles, Debelica y Thies (2009) mencionan que el género Myotis tiene escamas coronales simples y que T. brasiliensis tiene escamas coronales dentadas al igual que el género Nyctinomops. De otra parte, Sessions et al. (2014) coindice con Debelica y Thies (2009) en la descripción de los géneros Nyctinomops y Tadarida. En el presente estudio se concuerda con los autores que describen las escamas coronales como divaricadas o divergentes y denticuladas o dentadas. Esto se comprueba en la subfamilia Tomopeatinae (Tomopeas ravus) con escamas divaricadas denticuladas y en la subfamilia Molossinae (Mormopterus kalinowskii, Nyctinimops aurispinosus, N. laticaudatus, N. macrotis y Tadarida brasiliensis) con escamas divaricadas dentadas y escamas divergentes dentadas (Molossus molossus y Promops davisoni).
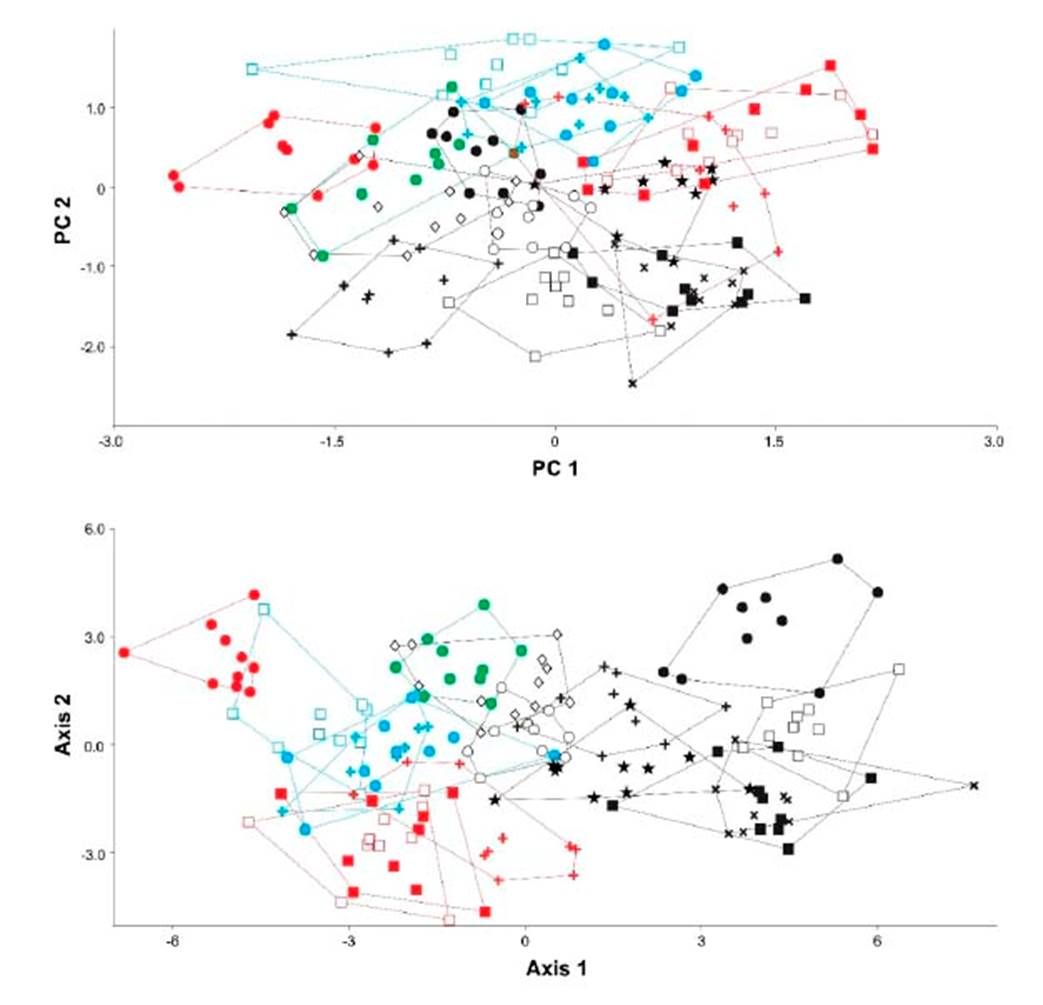
Figura 3 Resultados del análisis de componentes principales ACP (arriba) y del análisis de función discriminante AFD (abajo) de pelos de guardia de murciélagos (n=16), extraído desde cinco medidas. En color rojo, familia Phyllostomidae (Desmodus rotundus, círculos llenos; Glossophaga valens, cruces; Platalina genovensium, cuadrados vacíos; Sturnira bogotensis, cuadrados llenos). En color verde, familia Furipteridae (Amorphochilus schnablii). En color negro, familia Molossidae (Tomopeas ravus, círculos llenos; Molossus molossus, cruces; Mormopterus kalinowskii, cuadrados vacíos; Nyctinomops aurispinosus, cuadrados llenos; Nyctinomops laticaudatus, equis; Nyctinomops macrotis, círculo vacío; Promops davisoni, diamante vacío; Tadarida brasiliensis, estrellas). En color celeste, familia Vespertilionidae (Eptesicus montanus, círculos llenos; Myotis atacamensis, cruces; Myotis cf oxyotus, cuadrados vacíos)
Cuadro 5 Clave dicotómica para determinar las especies de murciélagos del Desierto costero del sur de Perú

En la familia Vespertilionidae se encontró que las escamas son de tipo hastada asimétrica. Benedict (1957) observó escamas coronales hastadas, donde las
escamas distales pueden ser sinuadas, enteras o lobadas, pudiendo diferenciarse algunos géneros. Albayrak y Coban (1997) describen las escamas del género Myotis con forma de vaso, y las del género Tadarida con forma de espiga; Amman et al. (2002) indican que los murciélagos vespertilionidos presentan formas de escama que van desde coronal lobado amplio a coronal hastado, pero nunca corona dentada; Pierallini et al. (2004) observaron escamas coronales divergentes; y Schaetz et al. (2009) describen escamas imbricadas con forma de lengüeta y escamas imbricadas truncas. Los resultados de este estudio concuerdan con Benedict (1957) y Schaetz et al. (2009) ya que en esta familia (Eptesicus montanus, Myotis atacamensis y M. cf. oxyotus) se observaron las escamas coronales de tipo alternada asimétrica.
Dichas variaciones en las descripciones son aparentemente producto de las limitaciones tecnológicas de la época; por consiguiente se espera que los resultados aquí presentados sirvan de un marco de referencia contemporáneo.
A nivel ecológico y anatómico se han planteado algunas hipótesis para explicar las modificaciones de la morfología de la cutícula en los pelos de murciélagos. Así, Howell y Hodking (1976) mencionan que los murciélagos polinizadores presentan escamas muy divergentes y divididas que les permitiría colectar una mayor cantidad de polen de plantas, mientras que los murciélagos no asociados a flores presentan escamas más apretadas. Así mismo, Meyer et al. (1995) sostienen que ciertas formas de cutícula facilitan el vuelo debido a que generan una menor turbulencia. No obstante, es necesario más estudios que permitan explicar la variada morfología cuticular y la función que cumplen en las diversas familias de murciélagos que ocurren en Perú.
CONCLUSIÓN
Se registraron cinco tipos de patrones cuticulares coronales y seis configuraciones del borde de las escamas, los cuales junto con datos morfométricos permiten diferenciar los especímenes hasta el nivel de especie.
Se documenta, por primera vez, una descripción de la cutícula de pelos de guardia para Glossophaga valens, Platalina genovensium, Sturnira bogotensis, Amorphochilus schnablii, Tomopeas ravus, Molossus molossus, Mormopterus kalinowskii, Nyctinomops aurispinosus, Nyctinomops laticaudatus, Nyctinomops macrotis, Promops davisoni, Tadarida brasiliensis, Eptesicus montanus, Myotis atacamensis y Myotis cf. oxyotus

