











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
El crecimiento de la población humana, la intensificación económica, el cambio climático y la capacidad de mantener la biodiversidad entre otros aspectos dificulta mantener el ecosistema, especialmente en países en desarrollo (Bedunah y Angerer, 2012). La demanda de alimentos para la humanidad se incrementa día a día, por lo que se requiere incrementar la producción animal y vegetal. El incremento en la producción animal provocaría un incremento en la demanda de pastizales y forrajes (Estell et al., 2012)
De acuerdo con datos del Instituto Nacional de Estadística e Informática (INEI, 2012) se estima que 86.3% de los pastizales naturales del Perú están localizados en la región Puna, ecosistema de crucial importancia para la subsistencia de la población (Bedunah y Angerer, 2012), pues es fuente de alimento para el ganado. Además, este es un recurso esencial en la provisión de servicios hidrológicos, ya que mejoran la infiltración del agua y reducen la erosión de los suelos (Mosquera et al., 2022); sin embargo, más del 40% de estos pastizales estarían en condición pobre y en proceso de deterioro por sobrepastoreo e inadecuado manejo (Zarria y Flores, 2015), lo que limita la producción animal en estos ecosistemas.
Gran parte de los pastizales naturales son pajonales dominados por especies de los géneros Festuca, Stipa y Calamagrostis, siendo la especie Festuca dolichophylla una de las gramíneas nativas más comunes en la región Puna, especie que aporta buena cantidad de forraje para la alimentación del ganado. La mejora de la condición de los pastizales puede realizarse por resiembra de especies nativas, abonamiento, clausura y manejo del pastoreo, además del mejoramiento genético de especies promisorias. Especies nativas pueden ser de gran utilidad para la revegetación de regiones áridas y también para uso como forraje (Kozub et al., 2017), pero deben ser seleccionadas en función de su potencialidad de adaptación y producción forrajera en ambientes específicos. Para ello es necesario comprender los procesos básicos de crecimiento, especialmente de las gramíneas perennes, para evaluar el potencial de crecimiento de las plantas dentro de ambientes específicos (Nelson, 2000).Asimismo, se pueden desarrollar cultivares localmente adaptados para su aprovechamiento como forraje o en procesos de revegetación.
Para un mejor aprovechamiento de Festuca dolichophylla como forraje o para su uso en procesos de revegetación es necesario conocer las potencialidades que diferentes poblaciones pueden tener bajo condiciones específicas. El objetivo de este estudio fue evaluar las tasas de sobrevivencia y patrones de crecimiento en poblaciones de Festuca dolichophylla bajo condiciones similares.
MATERIALES Y MÉTODOS
Parcela Experimental
La parcela experimental se localizó en el sector «Pucro Corral» de la Comunidad Campesina Pastales Huando (CCPH) en el distrito de Ascensión, región Huancavelica, Perú, a una altitud de 4620 msnm. Los datos de precipitación anual y temperatura media obtenidos desde la base de datos PISCO fueron de 899 mm (Aybar et al., 2020) y 8.1 °C (Huerta et al., 2018), respectivamente. La mayor precipitación se presenta desde octubre a abril y coincide con las temperaturas medias más altas; mientras que de mayo a septiembre presenta las precipitaciones más bajas y coinciden con las temperaturas más bajas.
La parcela experimental tuvo 150 m2 (10.0 x 15.0 m) dentro de un área ligeramente inclinada y con suelo uniforme. La textura del suelo era franco-arcilloso, con promedios (error estándar) de pH 5.8 (0.070) y contenido de nitrógeno, fósforo y potasio de 0.34% (0.006), 5.9 ppm (0.967) y 556.5 ppm (30.239), respectivamente. Dentro de la parcela se realizaron marcaciones con distanciamiento de 1.0 m, que fueron codificados en serpentín para la instalación de las plantas (10 filas y 15 columnas).
Poblaciones y Accesiones
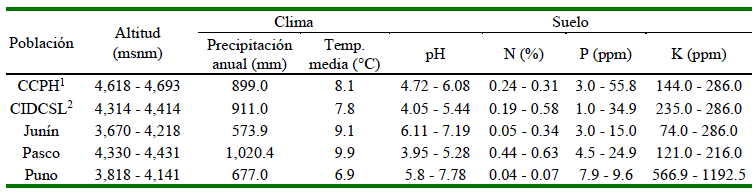
Se obtuvieron accesiones de cinco poblaciones: en Junín, Pasco y Puno, además del CCPH y del Centro de Investigación y Desarrollo de Camélidos Sudamericanos Lachocc de la Universidad Nacional de Huancavelica (CIDCSL). Los rangos de altitud, precipitación y temperatura, obtenidos desde la base de datos PISCO, así como los contenidos de nitrógeno, fósforo y potasio tuvieron gran variación entre poblaciones (Cuadro 1).
Instalación, Monitoreo y Análisis de Datos
Se obtuvieron cinco accesiones de cada población, elegidas por su mayor vigor. Cada accesión fue extraída con su raíz y fue dividida en seis plantas pequeñas (macollos) de 2.5 cm de diámetro. Se codificaron y almacenaron en bolsas de papel con parte de su propia tierra para ser transportados dentro de cajas refrigerantes a la parcela experimental.
La asignación de cada planta a los puntos de la parcela experimental, previamente marcados, se realizó de forma aleatoria. La instalación de las plantas se realizó durante las dos primeras semanas de diciembre de 2019. La instalación se realizó a una profundidad de 10.0 cm con ayuda de un pico. Además de las plantas experimentales, se instalaron dos líneas adicionales de plantas (procedentes de la CCPH) para controlar el efecto borde.
Después de 12 meses de establecimiento se realizó el corte de uniformización a una altura de 5.0 cm en todas las plantas y al mismo tiempo se registraron las tasas de sobrevivencia por inspección directa. La altura a la hoja bandera fue considerada como altura de planta y medida con un vernier digital, el número de tallos por planta y el número de hojas por planta se obtuvo por conteo directo, teniendo en cuenta tallos y hojas mayores o iguales a 2.0 cm de longitud, determinado cada 14 días a partir del corte de uniformización hasta mayo de 2021. La tasa de crecimiento (mm/día), tasa de aparición de tallos (n/día) y la tasa de aparición de hojas (n/día) se calculó con los datos obtenidos y se analizó para cada mes.
Para la tasa de sobrevivencia, obtenido en porcentaje para cada accesión, se utilizó un análisis de varianza de una sola vía incluyendo el efecto de la población como único factor. En cambio, los datos relacionados a los patrones de crecimiento fueron analizados utilizando el siguiente modelo lineal: Yijk = µ +Pi + Aj (Pi) + Bk + εijk, donde Yijk: Características de patrones de crecimiento de Festuca dolichophylla, µ: Media general, Pi: Efecto de la i-ésima población, Aj(Pi): Efecto de la j-ésima accesión dentro de la i-ésima población Bk: Efecto del k-ésimo bloque (fila), y εijk: Error experimental.
Se hizo detección de datos atípicos por medio de un análisis exploratorio y estos fueron excluidos de posteriores análisis. En todos los procesos de análisis se utilizó el software R 4.0.3 y la comparación de medias se realizó por el método de Tukey con un nivel de significación de 0.05.
RESULTADOS Y DISCUSIÓN
Tasa de Sobrevivencia
Las mayores tasas de sobrevivencia se encontraron en plantas de Festuca dolichophylla procedentes de Puno, seguida por plantas de la CCPH. En ambos casos, las tasas de sobrevivencia fueron superiores al 85%, mientras que en las otras poblaciones fueron inferiores al 70% (Cuadro 2). Además, hubo diferentes grados de variabilidad dentro de cada población, lo que concuerda con Gil et al. (2015), quienes estudiaron 13 poblaciones de Trichloris crinita, gramínea nativa de Argentina, y encontraron persistencias desde 10% hasta 95%. La variabilidad es muy importante cuando se busca plantas con gran potencial de adaptación a diferentes ambientes.
Cuadro 2. Poblaciones, accesiones y tasas de sobrevivencia (error estándar) en Festuca dolichophylla
| Población | Accesiones | Repeticiones | Plantas instaladas | Plantas vivas | Tasa de sobrevivencia (%) |
|---|---|---|---|---|---|
| CCPH1 | 5 | 6 | 30 | 26 | 86.7 (8.2)ab |
| CIDCSL2 | 5 | 6 | 30 | 13 | 43.3 (13.5)b |
| Junín | 5 | 6 | 30 | 19 | 63.3 (16.2)ab |
| Pasco | 5 | 6 | 30 | 20 | 66.7 (11.8)ab |
| Puno | 5 | 6 | 30 | 29 | 96.7 (3.3)a |
| Total | 25 | 150 | 107 | 71.3 (6.1) |
1 Comunidad Campesina Pastales Huando, 2 Centro de Investigación y Desarrollo de Camélidos Sudamericanos Lachocc.
a,b Superíndices diferentes dentro de columnas muestran diferencias significativas (p<0.05) a la prueba de Tukey
La tasa de sobrevivencia de las plantas de Puno fue superior a las del CIDCSL, pero similares al resto de las poblaciones (Cuadro 2), a pesar de que las diferencias numéricas fueron superiores al 5.0%, debido principalmente a la elevada variabilidad. Por otro lado, Gil et al. (2015) encontraron diferencias entre poblaciones de Trichloris crinita para persistencia en un ambiente uniforme. Para otras especies nativas, como Festuca humilior y Calamagrostis macrophylla se reportan tasas de mortalidad dependientes del abonamiento del suelo (Tácuna et al., 2015), mientras que Gonzales et al. (2021) no encontraron diferencias entre especies nativas e introducidas para sobrevivencia a los tres meses de implantación, siendo superior al 78%. En el presente estudio la temperatura del suelo (datos no publicados) fue uniforme, las características químicas del suelo fueron uniformes (bajos errores estándar) y la fuente de agua fue únicamente de lluvias, de allí que la variabilidad mostrada se atribuye principalmente al componente genético de cada población. De hecho, las adaptaciones al medio ambiente de cada población, en especial los relacionados con el suelo y disponibilidad de agua habría generado una amplia variación fenotípica y genética en las poblaciones (Kozub et al. (2017).
Patrones de Crecimiento
Las poblaciones tuvieron un efecto significativo sobre la altura de planta de F. dolichophylla (p<0.05), siendo las plantas procedentes de Puno las que mostraron mayor altura con respecto a las demás poblaciones durante el estudio, excepto con las plantas del CIDCSL (Cuadro 3). En todo el periodo de evaluación se encontró elevada variabilidad, reflejado en sus altos errores estándar. Estos resultados concuerdan con el estudio de Gil et al. (2015), quienes reportaron variabilidad y efecto de poblaciones para alturas al dosel y a la panoja más alta en trece poblaciones de Trichloris crinita.
Cuadro 3. Altura de planta (mm ± error estándar) en poblaciones de Festuca dolichophylla , según el mes de evaluación
| Población | n | Diciembre | Enero | Febrero | Marzo | Abril | Mayo |
|---|---|---|---|---|---|---|---|
| CCPH1 | 25 | 91.7 (3.6)bc | 153.4 (12.3)bc | 194.3 (19.9)bc | 229.0 (25.6)bc | 252.6 (28.4)bc | 258.1 (29.0)bc |
| CIDCSL2 | 13 | 106.2 (5.6)ab | 212.5 (17.8)ab | 292.2 (31.4)ab | 349.8 (37.9)ab | 362.9 (38.7)ab | 363.3 (38.8)ab |
| Junín | 18 | 89.1 (3.7)bc | 146.0 (8.5)c | 180.8 (10.6)c | 201.2 (12.3)c | 213.4 (11.1)bc | 217.1 (11.6)bc |
| Pasco | 19 | 85.4 (3.7)c | 148.4 (9.8)bc | 174.9 (14.4)c | 188.6 (17.9)c | 198.3 (19.8)c | 199.7 (20.2)c |
| Puno | 25 | 114.6 (3.7)a | 231.7 (8.8)a | 332.0 (16.6)a | 420.5 (19.4)a | 454.1 (17.3)a | 458.5 (16.5)a |
| Total | 100 | 97.6 (2.1) | 178.4 (6.2) | 235.3 (10.6) | 279.9 (13.8) | 299.9 (14.6) | 303.4 (14.6) |
1 Comunidad Campesina Pastales Huando, 2 Centro de Investigación y Desarrollo de Camélidos Sudamericanos Lachocc.
a,b,c Superíndices diferentes dentro de columnas muestran diferencias significativas (p<0.05) a la prueba de Tukey
La tasa de crecimiento fue influenciada de manera significativa por las poblaciones en el periodo de enero a marzo (p<0.05), en tanto que no se encontró diferencia significativa en abril y mayo (p>0.05). Plantas procedentes de Puno mostraron mayores tasas de crecimiento en comparación al resto de las poblaciones, con excepción a las plantas del CIDCSL (Figura 1) en el periodo de enero a marzo. Por otro lado, se encontró altos niveles de variabilidad, que se reflejan en el error estándar. En estudios en Lolium perenne, Festuca arundinacea y Dactylis glomerata se reportó efecto de la especie sobre la extensión laminar y tasa de crecimiento expresado en mg/día (Ganderats y Hepp, 2003), así como del volumen de agua sobre la elongación de tallos y hojas en Brachiaria decumbens (de Souza et al., 2008).
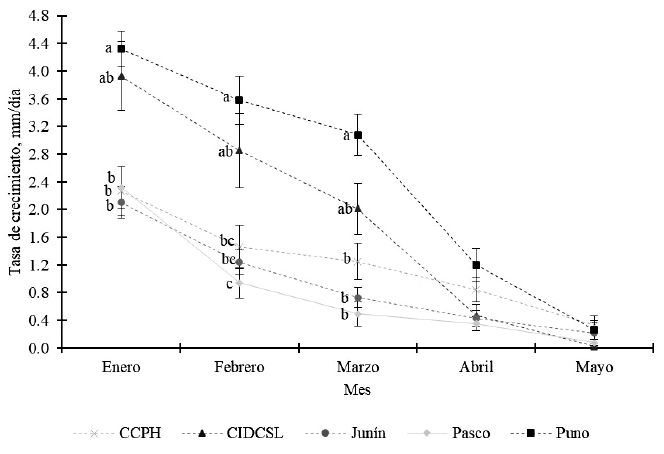
Figura 1. Tasa de crecimiento en poblaciones de Festuca dolichophylla. Letras iguales en el mismo mes indican diferencias significativas (p<0.05) a la prueba de Tukey. Las barras representan el error estándar
Los hallazgos muestran un patrón de crecimiento mayor al inicio del periodo de evaluación, coincidiendo con el incremento de las precipitaciones, para luego disminuir hasta mayo, donde el crecimiento es casi nulo, coincidiendo con el inicio de la época seca. La sequía induce la reducción en la presión de turgencia de las hojas y dan como resultado una inhibición simultánea, tanto de la expansión como de la diferenciación celular, provocando la reducción en la tasa de elongación y aumentando la duración (Coussement et al., 2021).
El número de tallos fue influenciado (p>0.05) por la población desde febrero a mayo (p<0.05), pero no en diciembre y enero. En todos los casos con altos valores de error estándar indicando alta variabilidad (Cuadro 4). Las plantas de Puno mostraron mayor número de tallos de febrero a abril con respecto al resto de las poblaciones; asimismo, Festuca dolichophylla de Puno mostró mayores valores en mayo que las otras poblaciones, pero fue similar a las plantas del CIDCSL.
Cuadro 4. Número de tallos por planta (error estándar) en poblaciones de Festuca dolichophylla según el mes de evaluación
| Población | n | Diciembre | Enero | Febrero | Marzo | Abril | Mayo |
|---|---|---|---|---|---|---|---|
| CCPH1 | 25 | 0.00 (0.00) | 0.36 (0.14) | 2.36 (0.36)b | 3.27 (0.44)b | 3.45 (0.45)b | 3.55 (0.45)b |
| CIDCSL2 | 13 | 0.08 (0.08) | 0.77 (0.23) | 2.77 (0.75)b | 3.54 (0.93)b | 3.92 (1.00)b | 4.23 (1.19)ab |
| Junín | 18 | 0.36 (0.13) | 0.86 (0.27) | 2.07 (0.32)b | 2.43 (0.37)b | 2.86 (0.39)b | 2.93 (0.38)b |
| Pasco | 19 | 0.23 (0.12) | 0.46 (0.14) | 2.23 (0.44)b | 3.00 (0.65)b | 3.08 (0.65)b | 3.08 (0.65)b |
| Puno | 25 | 0.00 (0.00) | 0.92 (0.26) | 5.84 (0.52)a | 7.44 (0.70)a | 7.92 (0.76)a | 8.32 (0.82)a |
| Total | 100 | 0.12 (0.03) | 0.66 (0.11) | 3.21 (0.28) | 4.15 (0.36) | 4.45 (0.38) | 4.63 (0.41) |
1 Comunidad Campesina Pastales Huando, 2 Centro de Investigación y Desarrollo de Camélidos Sudamericanos Lachocc.
a,b,c Superíndices diferentes dentro de columnas muestran diferencias significativas (p<0.05) a la prueba de Tukey
La tasa de aparición de tallos no mostró diferencias significativas entre poblaciones en los meses de enero, marzo, abril y mayo (p>0.05); pero se encontraron diferencias significativas en el mes de febrero (p<0.05) donde plantas de Puno tuvieron mayor tasa de aparición de tallos en comparación al resto de las poblaciones (Figura 2), lo cual muestra alta variabilidad para esta característica. Gil et al. (2015) también reportaron variabilidad y diferencias entre poblaciones de Trichloris crinita para el número de panojas.
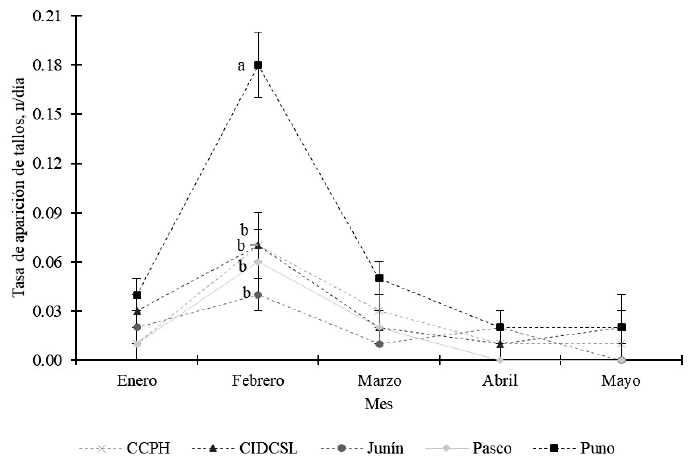
Figura 2. Tasa de aparición de tallos en poblaciones de Festuca dolichophylla. Letras iguales en el mismo mes indican diferencias significativas (p<0.05) a la prueba de Tukey. Las barras representan el error estándar
El número de hojas fue influenciado por la población (p<0.05) en los meses de diciembre y enero, mas no en febrero, marzo, abril y mayo (p>0.05), en todos los casos con elevada variabilidad (Cuadro 5). El número de hojas en diciembre fue superior en las plantas procedentes de Puno, en tanto que solo en enero fue superior a las plantas de la CCPH. En este sentido, da Silva et al. (2009) reportaron alta variabilidad para el número de hojas vivas, además de que a menor es la altura de corte en Panicum máximum el número de hojas/macollo es menor. En Urochloa brizantha y Megathyrsus maximus, Mota et al. (2021) encontraron un mayor número de hojas debidas al abona-miento con azufre y potasio.
Cuadro 5. Número de hojas por planta (error estándar) de poblaciones de Festuca dolichophylla según el mes de evaluación
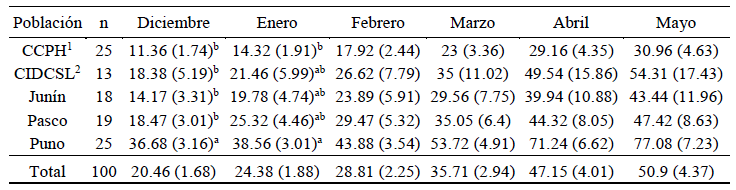
1 Comunidad Campesina Pastales Huando, 2 Centro de Investigación y Desarrollo de Camélidos Sudamericanos Lachocc.
a,b,c Superíndices diferentes dentro de columnas muestran diferencias significativas (p<0.05) a la prueba de Tukey
La tasa de aparición de hojas no mostró diferencias significativas entre poblaciones (p>0.05) durante todo el proceso de evaluación, pero se encontró alta variabilidad en las poblaciones (Figura 3). La tasa de aparición de hojas incrementa de enero a abril en todas las poblaciones, para disminuir en mayo. Así, da Silva et al. (2009) reportaron una disminución en la tasa de aparición de hojas a lo largo del periodo de crecimiento en P. maximum, una gramínea tropical, diferente a los hallazgos en el presente estudio; en cambio Mota et al. (2021) no encontraron diferencias en la tasa de aparición de hojas debido al abonamiento con azufre y potasio en Urochloa brizantha y Megathyrsus maximus.
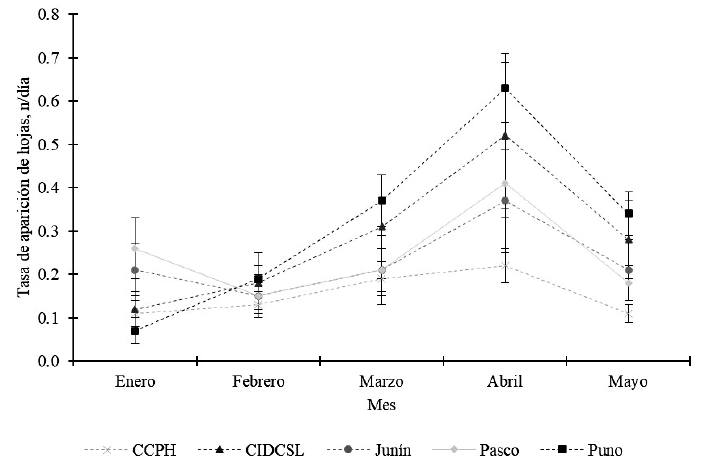
Figura 3. Tasa de aparición de hojas en poblaciones de Festuca dolichophylla. Letras iguales en el mismo mes indican diferencias significativas (p<0.05) a la prueba de Tukey. Las barras representan el error estándar
La altura de planta, número de tallos por planta, número de hojas por planta, así como la tasa de crecimiento, tasa de aparición de tallos y tasa de aparición de hojas tuvieron alta variabilidad. Resultados similares fueron obtenidos por Gil et al. (2015) en poblaciones de Trichloris crinita con relación a la altura de plantas, número de hojas, número de panículas y número de macollos, en tanto que Kozub et al. (2017), para dicha especie encontraron alta variabilidad para características relacionadas con la revegetación y uso como forraje. Esta variabilidad habría sido generada por los diferentes ambientes de origen a los cuales se adaptaron cada población, especialmente a los relacionados con las características del suelo, disponibilidad de agua, estrés abiótico (sequía, salinidad) y la competencia con otras especias nativas (Kozub et al. (2017).
En el presente estudio, las plantas procedentes de las diferentes poblaciones fueron evaluadas en un ambiente uniforme; por lo que la variación y diferencias encontradas se deberían principalmente a la diferente constitución genética de cada población. Esta variabilidad es muy importante para futuros programas de mejoramiento genético en esta especie con fines de producción forrajera o revegetación; sin embargo, previamente se requiere realizar una caracterización morfológica y genética como lo sugieren Oliveira et al. (2016) a fin de poder identificar poblaciones potenciales con propósitos específicos.
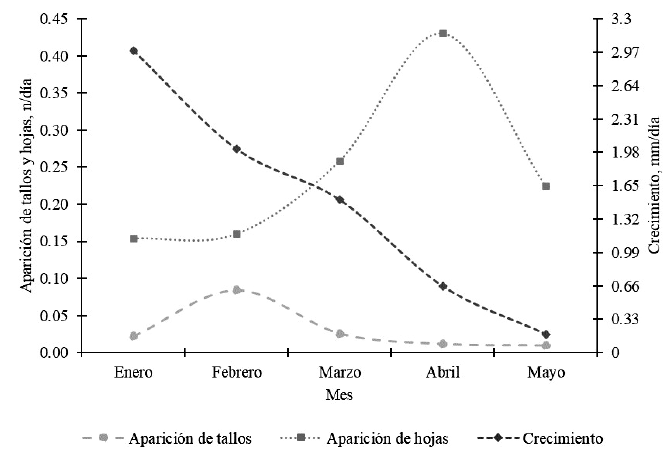
Por último, se encontró un patrón de crecimiento ordenado de las partes de la planta (Figura 4). F. dolichophylla inicia con crecimiento en tamaño (altura de planta) en el mes de enero para luego descender gradualmente; en seguida se incrementa el número de tallos, mostrando la mayor tasa de aparición de tallos en febrero, para finalmente incrementar el número de hojas, especialmente en abril. Matthew (1996) encontró una activación secuencial de la emergencia de tallos, formación de raíces y crecimiento de hojas en Lolium perenne; patrón que sería similar al del presente estudio para tallos y hojas.
CONCLUSIONES
Existe alta variabilidad y diferencias entre poblaciones de Festuca dolichophylla para tasa de sobrevivencia, altura de planta, tasa de crecimiento, número de tallos, tasa de aparición de tallos y número de hojas; a excepción de la tasa de aparición de hojas.
Festuca dolichophylla muestra un patrón de crecimiento ordenado; primero crece en tamaño, luego incrementa el número de tallos y finalmente aumenta el número de hojas.

