











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Recent publication treats Miconia Ruiz & Pavón, Melastomataceae, as the only genus of Miconieae tribe with ca. 1900 species, and it can be considered as seventh largest genus of flowering plants that is restricted to Neotropics (Michelangeli et al., 2022). Miconia can be recognized by the same characters as the tribe, viz. terminal or axillary inflorescences, very rarely cauliflorous (then the inflorescences cymose), each flower subtended by only one pair of bracteoles, or more rarely none (but never by two pairs), anthers without pedoconnectives, leaves without acicular raphides, and fruit a berry (Michelangeli et al., 2022).
According to Michelangeli et al. (2022) the Miconia recognition as a unique genus within Miconieae requires new multidisciplinary study. Fruit and seed characters have been promising and useful in the taxonomy of eudicots (for example, the classic works of Corner, 1976 and Roth, 1977). Fruits and seeds have been investigated in Melastomataceae with a descriptive, taxonomic and evolutionary purposes (see Baumgratz, 1983-1985; Clausing et al., 2000; Cortez & Carmello-Guerreiro, 2008; Ramirez, 2011; Ocampo & Almeda, 2013; Ocampo et al., 2014; Ribeiro, 2016).
Miconieae fruits are typically berries (indehiscent fruits) usually with fleshy placentas and fused tissues which are characterized by the absence of lignified tissues, few sclereids in the pericarp, and often by an endocarp that collapses early (Clausing et al., 2000). A good example of Miconia with fruit of similar structure is shown by Cortez & Carmello-Guerreiro (2008) in M. albicans (Sw.) Triana. The taxonomic significance of seed structure has been explored most frequently in Miconieae (Groenendijk et al., 1996; Martin & Michelangeli, 2009; Ocampo & Almeda, 2013; Ocampo et al., 2014; Ocampo et al., 2022); the characters of the tribe's seeds are variable and diverse, especially in Miconia (Ocampo & Almeda, 2013).
The main purpose of this investigation was to ascertain the usefulness of fruit and seed characters as a possible criterion for separating species of Miconia. Thus, six species of Miconia belonging to four different sections were selected.
Material and methods
Plant samples
Flowers and developing fruits of six species of Miconia (Table 1) were collected from herbarium specimens and fresh specimens. The selected species of Miconia belong to the vegetation of southern Brazil, especially to the rupestrian fields, which suffer anthropic destructive action. Fresh specimens were collected in Guartelá State Park, located in the municipality of Tibagi, Brazil. Herbarium specimens were obtained from the Herbarium of the Museu Botânico Municipal de Curitiba (MBM), Brazil.
Table 1 Species investigated with information on sample type and herbarium accession number.
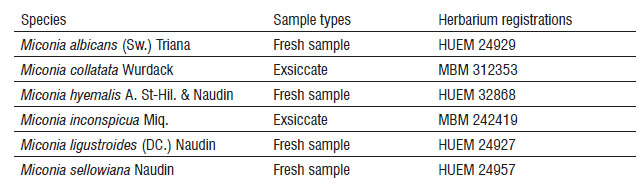
HUEM - Herbarium at the State University of Maringá; MBM - Herbarium of the Municipal Botanical Museum of Curitiba
Sample preparation
The herbarium samples were rehydrated with boiling water, immersed in 5% potassium hydroxide (KOH) and submitted to the 10%, 30%, 50% and 70% ethyl series, according to Smith & Smith (1942), with modifications (change in percentage from 2% to 5% KOH and time of exposure to the reagent). Herbarium and fresh samples were fixed in FAA 50 (formaldehyde, acetic acid and ethyl alcohol), and later dehydrated using the series of ethyl alcohol with concentrations of 80%, 90% and 100%.
Analysis of samples under a light microscope
In sequence, the developing flowers and fruits were embedded in Leica historresin, according to the manufacturer's specification. The samples were sectioned on a rotation microtome, whose sections obtained 6-8 µm in thickness were stained in Toluidine Blue in phosphate buffer (pH = 6.8) (O'Brien et al., 1964). Photomicrographs were taken with a Leica ICC50 light microscope with an attached digital camera, using the Leica Application Suite software, version 1.8.
Results and discussion
The fruits of M. hyemalis originate from the inferior ovary, while the fruits of the other five species derive from the semi-inferior ovary (Figure 1A,B). The six species have a hypanthium with a free portion (perigynous hypanthium), whose length varies in different flowers, and another part that is adnate to the ovary (gynoecial hypanthium) (Figure 1A,B). The ovaries generally have 3 carpels and 3 locules, but M. hyemalis exhibits 3 or 4 carpels and locules, and M. ligustroides (Figure 1C) consists of 4 carpels and locules (Table 2). The placentation is axile and the ovules attach to the septum at different heights.
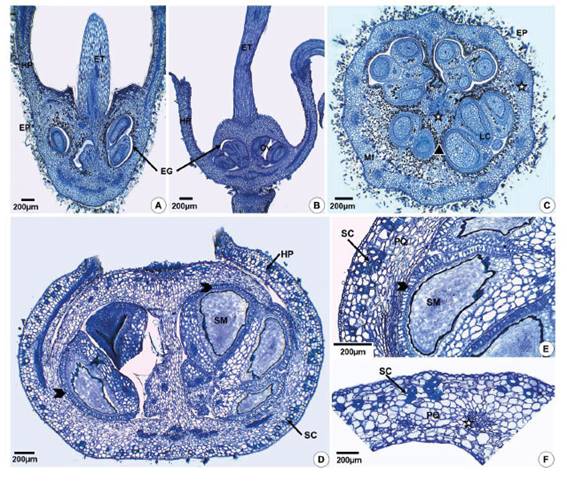
Figure 1 Ovary and fruit structure of Miconia hyemalis (A,F) and Miconia sellowiana (B-E) in longitudinal (A,B,D,E) and cross- (C,F) sections. A,B - Inferior and semi-inferior ovary. C - Ovary with 4 carpels and locules. D - Fruit originating from semi-inferior ovary showing hypanthium adnate to pericarp and free hypanthium (perigynous). E - Detail of the pericarp and seed showing sclereids and exotesta of macrosclereids (arrowhead). F - Detail of the pericarp with sclereids. (EG=glabrous inner epidermis; EP=hairy outer epidermis; ET=style; HP=hypanthium; LC=locule; Mf=mesophyll; Ov=ovule; PQ=parenchyma; SM=seed; SC=sclereids; stars indicate vascular bundles).
Table 2 Taxonomically important features that may be useful in separating the six species of Miconia.
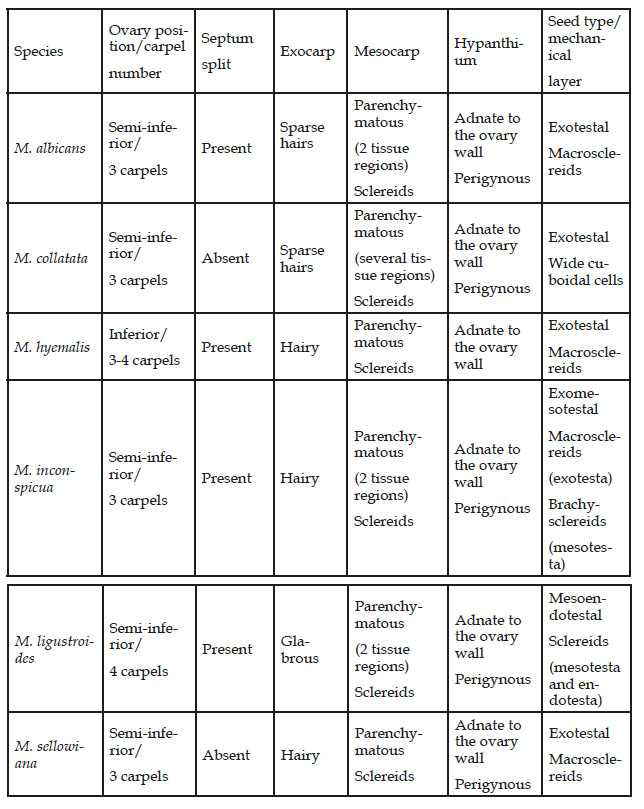
The ovary wall of all species has a parenchymatous mesophyll (Figure 1A-C), which is generally compact on the outside and spongy or lacunose on the inside. Parenchymatous mesophyll seems to be common in angiosperm ovaries (Fahn, 1990; Souza, 2022), but Gonçales (2020) reported the presence of collenchyma in the ovary of some species of Miconia. The ovary septa (Figure 1A,B) of the six Miconia species consist of epidermis and spongy parenchyma, and they can exhibit splits (Table 2), which are considered by Carr & Carr (1961) as compitum in multilocular ovaries of flowering plants. In the anatomical study of Miconia flowers, Gonçales (2020) treated the investigated species as eu-syncarpous (term proposed by Carr & Carr, 1961) with compitum characterized as splits in the septa or a single strand of transmitting tissue in the style.
The fruit wall in the region that has adnate hypanthium (gynoecial hypanthium) (Figure 1D-F) consists of epidermal exocarp and endocarp, and parenchymatous mesocarp. The mesocarp may exhibit different regions of parenchyma (Table 2). In the mesocarp with two parenchyma regions the outermost part is composed of more or less rounded cells with sclereids, and the innermost portion exhibits tangentially elongated cells. In fruits from semi-inferior ovaries (Figure 1D), the upper portion of the fruit wall is essentially carpellary and is surrounded by the perigynous hypanthium. Here the wall is constituted by the papillose exocarp, and the mesocarp consists of more homogeneous parenchyma with few sclereids (Figure 1D).
All Miconia fruits investigated here have sclereids in the mesocarp (Figure 1D-F; Table 2), which are sclerenchymatous cells that can occur singly or in clusters in the fleshy pericarp of berries (Roth, 1977; Souza, 2022). Cortez & Carmello-Guerreiro (2008) also found sclereids in the mesocarp of Miconia albicans and suggested that they must have originated from sclerification of parenchyma cells and not from sclereid primordia.
Miconia fruits were classified as type pomaceous subtype berry based on the fruit classification proposed by Souza (2022) and Souza et al. (2022). Pomaceous fruit for these authors originates from an inferior or semi-inferior ovary, and exhibits several subtypes. Clausing et al. (2000) considered the fruit of Miconieae, particularly a species of Clidemia D. Don, as a soft berry that is characterized by the absence of lignified tissues and a dearth of sclereids in the fruit walls. Cortez & Carmello-Guerreiro (2008) analyzed the ontogeny of the fruit of Miconia albicans, and also considered its fruit as a berry, originating from a semi-inferior ovary. One of the characters considered diagnostic for the tribe Miconieae and the genus Miconia by Michelangeli et al. (2022) is the type of fruit like berry. Berry definition for Souza (2022) and Souza et al. (2022) is a simple fruit that must originate from the superior ovary.
Miconia seeds originate from anatropous, bitegmic and crassinucellate ovules. Seeds have non-multiplicative testa and the tegmen usually collapses. Seeds are exotestal (Figure 1D,E) in M. albicans, M. collatata, M. hyemalis and M. sellowiana, exomesotestal in M. inconspicua and mesoendotestal in M. ligustroides (Table 2). In exotestal and exomesotestal seeds the outer epidermis (exotesta) consists of macrosclereids. Outer epidermal cells of M. ligustroides are prismatic, but with thin walls.
Several studies have shown that seed morphology in Miconieae has taxonomic value, but almost all investigations describe seeds under scanning electron microscopy (Groenendijk et al., 1996; Martin & Michelangeli, 2009; Ocampo & Almeda, 2013; Ocampo et al., 2022). The anatomical analysis of the Miconieae seed is still little explored. In general, the seed coat characteristics as testa not multiplicative with outer epidermis that is made up of cuboid or shortly radially elongated lignified cells, and tegmen as pellicle no lignified are typical of Miconia (Corner, 1976). Miconia albicans seeds were also described by Cortez & Carmello-Guerreiro (2008) as having the sclerenchymatous palisade exotesta, and tegmen absent. In the case of the Miconia species studied here, we verified that the mechanical layer of the testa can occur in the outer epidermis (exotesta) or in the mesophyll (mesotesta) (Table 2).
It is not surprising, therefore, that the o fruit and seed ontogeny possesses many features of potential taxonomic significance. Ontogeny of fruits and seeds of six Miconia species provide many important features which are of taxonomic value at the specific level (Table 2). Features such as the position of the ovary (inferior or semi-inferior), carpel number (3 or 4 carpels), axile placentation (ovules attaching to the base, middle or apical region of the septum), exocarp indumentum, mesocarp structure (tissue region), type of pomaceous fruit (with hypanthium entirely adnate to the fruit wall or fruit with perigynous hypanthium), type of seed (exotestal, exomesotestal, mesoendotestal), and structure of the mechanical cell layer when it occurs in the testa (macroclereid or cuboidal sclerenchyma cells) should be highlighted.

