











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La crianza de llamas y alpacas es una actividad económica que ocupa a más de 500 mil unidades familiares de la región Andina, principalmente de Perú y Bolivia, donde en conjunto se maneja alrededor de 3.5 millones de alpacas y 3.6 millones de llamas (FAO, 2005ab), con alimentación basada en el uso de pastizales naturales de pobre disponibilidad y calidad (Reiner et al., 1987; Genin et al., 1994), clima adverso, con descensos térmicos de hasta -25 °C (Pinto et al., 2010), obteniéndose fibra de extraordinario valor textil (Iñiguez et al., 1998) y carne de alto valor nutricional (Polidori et al., 2007), con bajo contenido de grasa intramuscular (Salvá et al., 2009), pudiendo constituirse en fuentes alternativas de alimentos para las familias que viven en la región Altoandina (Cawthorn y Hoffman, 2014).
La nutrición de los camélidos sudamericanos cuenta con escasos reportes sobre los requerimientos energéticos; entre estos, el de Engelhardt y Schneider (1977), mediante el balance de C-N en llamas alimentadas con forrajes, estimándose un requerimiento de energía metabolizable de mantenimiento (EMm) de 61.2 kcal/kg BW0.75, el de Carmean et al. (1992) por calorimetría indirecta en llamas alimentadas con heno de avena y concentrado, calculando una EMm de 84.5 kcal/kg BW0.75, y el de Nielsen et al. (2014) por un método similar en llamas alimentadas con heno, obteniendo una EMm de 77.4 kcal/kg BW0.75, siendo estos trabajos realizados en altitudes cercanas al nivel del mar.
Van Saun (2006) propuso dos modelos de requerimientos de EMm para llamas y alpacas: Modelo I, que considera el valor promedio de los dos primeros reportes (72.85 kcal/kg BW0.75), y Modelo II, utilizando el valor de 84.5 kcal/kg BW0.75 reportado por Carmean et al. (1992). Los demás requerimientos energéticos (actividad física, termorregulación y otros) los estimó por extrapolación factorial a partir de datos de requerimientos energéticos de ovinos y caprinos, sugiriendo los mismos modelos para llamas y alpacas. El National Research Council incorporó la propuesta, publicando por primera vez los requerimientos nutricionales de los camélidos sudamericanos (NRC, 2007).
Los estudios comparativos realizados en contextos andinos han mostrado amplias diferencias entre llamas y alpacas en selectividad de dieta (Bryant y Farfan, 1984), consumo de alimento (Reiner et al., 1987), peso vivo y ganancia de peso (García et al., 2002), así como en la dinámica del nitrógeno (Robinson et al., 2005). No obstante, se tiene una mayor demanda de N para alpacas que para llamas, por lo que las extrapolaciones de los requerimientos fueron puestas en duda (Davies et al., 2007). Por esto se requiere realizar estudios específicos, considerando sobre todo que, en los Andes, los camélidos pueden alcanzar pesos adultos tardíamente a los 60 meses de edad (Wurzinger et al., 2005).
La técnica de sacrificio comparativo es un protocolo diseñado para medir los cambios en la composición corporal de los animales durante un periodo de alimentación (Lawes y Gilbert, 1859). La técnica, luego de su desuso por un siglo, fue retomada para medir la retención de energía en el cuerpo animal (Garrett et al., 1959). El proceso consiste en sacrificar un grupo representativo de animales al inicio del experimento, alimentar el resto de los animales por un periodo de tiempo y sacrificarlos al final para medir la retención de energía en sus cuerpos. La técnica se apoya en la asunción de que todos los animales tienen la misma composición corporal al inicio del experimento, por lo que su éxito radica en la habilidad de predecir con precisión la composición corporal inicial, a partir de la composición corporal de animales similares de la misma población que son sacrificados al inicio del experimento (Blaxter, 1967).
Los estudios piloto realizados en condiciones de la zona altoandina (4200 msnm), han mostrado la posibilidad de utilizar la técnica de sacrificio comparativo como herramienta para determinar los requerimientos energéticos de alpacas, a través de las mediciones de la energía disponible en el alimento y la retención de energía, en combinación con el balance de energía (Roque, 2009). Esto permitiría mejorar los programas de alimentación en los camélidos sudamericanos, con menor impacto ambiental (Hume et al., 2011).
La alimentación de llamas y alpacas en la zona altoandina se realiza en forma tradicional al pastoreo (San Martin y Bryant, 1989), sin considerar sus demandas nutricionales, obteniéndose niveles de producción relativamente bajos (Apaza y Quispe, 2016), con relación a otros contextos y sistemas (Riek y Gerken, 2007). Considerando los aspectos mencionados anteriormente, este estudio fue planeado a fin de determinar los requerimientos de energía metabolizable de mantenimiento y ganancia de peso vivo de llamas y alpacas machos en crecimiento, bajo condiciones de gran altitud, utilizando la técnica de sacrificio comparativo.
MATERIALES Y MÉTODOS
Localización
El trabajo se realizó en el Centro Experimental La Raya de la Universidad Nacional del Altiplano de Puno, ubicado en el distrito de Santa Rosa, provincia de Melgar, departamento de Puno, Perú, entre mayo y agosto de 2016. La zona se encuentra a 4200 msnm (Puna húmeda), con una temperatura media ambiental de 10.2 ± 2.1 °C, la misma que está dentro de la zona termoneutral de la llama (Riek et al., 2017). Los análisis químicos y el procesamiento de las muestras de carnes se realizaron en el Laboratorio de NutriciónAnimal de la Facultad de Medicina Veterinaria y Zootecnia, y en el Laboratorio de Carnes del Departamento deAgroindustria de la Facultad de Ciencias Agrarias de la Universidad Nacional del Altiplano de Puno.
Animales y Alimentación
Para el experimento de sacrificio comparativo se utilizaron 20 llamas macho (106.6 ± 7.7 kg de peso vivo, PV) y 20 alpacas macho (55.7 ± 4.9 kg PV), todos de 2 años de edad, procedentes de un sistema de alimentación en pradera nativa, dominada por especies de Festuca y Calamagrostis. Un grupo de cuatro llamas (102.2 ± 11.9 kg PV) y otro de cuatro alpacas (61.3 ± 4.5 kg PV), todos machos, acompañaron con experimentos de metabolismo convencional in vivo, para determinar el contenido de energía metabolizable de la dieta. El acostumbramiento para el manejo en confinamiento fue gradual y sistemático (Lund et al., 2012). Así, los animales consumieron la vegetación disponible en un potrero de pradera nativa durante la primera semana, heno entero de avena y alfalfa en la segunda semana, la dieta experimental ofrecida colectivamente en la tercera semana, y la dieta experimental ofrecida dentro de jaulas individuales de alimentación y jaulas metabólicas en la cuarta semana, hasta el sacrificio. El uso de animales para el estudio experimental fue autorizado por el Comité de Ética de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Nacional del Altiplano de Puno.
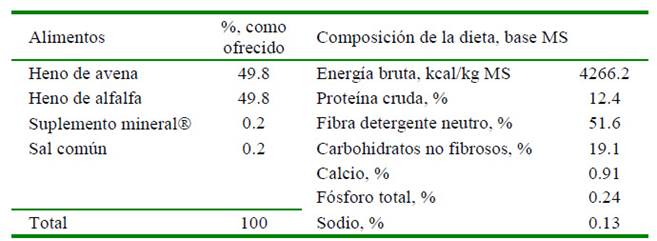
La dieta experimental estuvo conformada por una mezcla (1:1) de heno de avena (Avena sativa) y heno de alfalfa (Medicago sativa). Los henos cuales fueron procesados mecánicamente con molino-picador forrajero Trapp TRF-800, a través de una zaranda de 12 mmø. A la mezcla de forrajes se le agregó una fuente mineral comercial (0.2%) y sal común (0.2%). La dieta fue formulada para contener 12% de proteína cruda en materia seca, elaborada y ofrecida a los animales para cuatro niveles de consumo: mantenimiento, intermedio bajo, intermedio alto y ad libitum, correspondientes a 40, 50, 60 y 70 g MS/kg PV0.75, respectivamente, de la dieta tal como ofrecido, con 94.2% de materia seca (MS) (Cuadro 1). Los pesos de las dietas ofrecidas a los animales fueron registrados con una balanza digital T-Scale de 50/0.001 kg de capacidad.
Metodología
Los requerimientos de energía metabolizable de mantenimiento y ganancia se determinaron mediante la técnica de sacrificio comparativo (Lofgreen y Garrett, 1968), sujeta a la ecuación del balance de energía, EM = PC + RE, donde EM es la energía metabolizable consumida en la dieta, PC la producción de calor del animal, y RE la retención de energía en el cuerpo del animal después de un período de alimentación controlada.
Energía metabolizable de la dieta
La energía metabolizable de la dieta se determinó mediante experimentos de metabolismo convencional in vivo, por deducción de la energía bruta consumida en la dieta, las pérdidas de energía en heces, orina y gases (metano), en un diseño Cuadrado Latino 4 x 4, con cuatro animales por especie, cuatro etapas y cuatro niveles de oferta de la dieta experimental seco al aire (94.2% MS): mantenimiento, intermedio bajo, intermedio alto y ad libitum (40, 50, 60 y 70 g MS/kg PV0.75 , respectivamente). La producción de metano se determinó mediante espectroscopía infrarroja por transformada de Fourier (FTIR), con el analizador de gases Gasmet DX-4030 (Lassen et al., 2012), en una cámara de acumulación de gases (Goopy et al., 2011).
Retención de energía
La retención de energía se determinó mediante la técnica de sacrificio comparativo, con 20 animales por especie, divididos al azar en cinco grupos de cuatro animales por grupo. Luego de un periodo de alimentación con la dieta experimental, un grupo representativo de cuatro llamas y otro de cuatro alpacas fueron sacrificados mediante beneficio controlado en el camal del mismo centro experimental a fin de medir su contenido de energía corporal inicial. Los restantes cuatro grupos por especie fueron alimentados en jaulas individuales durante 55 días, en los mismos cuatro niveles de oferta de la dieta experimental seco al aire (94.2% MS), suministrada dos veces por día (08:00 y 13:00 horas). El agua se suministró a libre disposición. Los residuos de alimentos se retiraron y pesaron en forma diaria, a las 07:00 horas. El peso vivo de los animales se registró cada semana, por las mañanas, antes del suministro de la dieta, con una balanza digital de plataforma metálica de la marca T-Scale de 500/0.1 kg de capacidad.
Al final de la fase de alimentación, los cuatro grupos de animales por especie fueron sacrificados, mediante beneficio controlado en el camal del mismo centro experimental, aplicando las buenas prácticas, con adecuación al Reglamento Sanitario del Faenado deAnimales deAbasto (MINAGRI, 2012). Se separó el cuerpo en sangre, piel (incluida fibra), cabeza y patas, vísceras rojas (corazón, pulmones, hígado, riñones y bazo), vísceras blancas (intestinos y estómago, vacíos) incluida la grasa del omento, y carcasa (incluidos los testículos), registrándose sus pesos en caliente. La sangre, piel, vísceras y carcasa (Longissimus dorsi) de cada animal fueron muestreadas de inmediato (en caliente) para determinar su humedad. Cada carcasa fue dividida por la línea dorsal en dos mitades y cada mitad fue empaquetada en una bolsa de polietileno, sellada y congelada a -30 °C, hasta su procesamiento.
El procesamiento consistió en separar la carcasa por disección en músculos (incluido tendones y ligamentos), grasa y huesos. Los músculos, tendones, ligamentos y grasa fueron molidos con molino Berkel (1.5 HP) y mezclados en fresco con un Cutter de 3000 rpm hasta emulsión total, obteniéndose muestras frescas de 500 g. El producto fue secado por convección a 60 °C en estufa de laboratorio Ecocell®, hasta peso constante. Los huesos también fueron secados y molidos, y estas muestras fueron mezcladas en las mismas proporciones obtenidas en la composición corporal en materia seca, determinándose su contenido de energía bruta con un calorímetro de bomba (Parr Instrument Co, USA), a una presión de 25 atmósferas de oxígeno.
La energía corporal inicial de los animales que continuaron con la fase de alimentación fue estimada por regresión lineal simple del contenido de la energía corporal total en megacalorías (Mcal) de la materia seca del cuerpo vacío, sobre el peso vivo (kg) del grupo de animales sacrificados al inicio del experimento (Wolynetz y Sibbald, 1984), obteniéndose las ecuaciones respectivas (Cuadro 2).
Cuadro 2 Ecuaciones de predicción del contenido de energía corporal inicial de llamas y alpacas que continuaron con la fase de alimentación hasta el sacrificio final
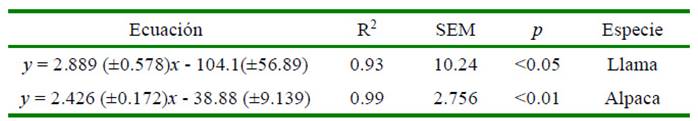
y: energía del cuerpo vacío del animal (Mcal); x: peso vivo del animal (kg)
La retención de energía (RE) corresponde a la diferencia entre el contenido inicial y final de energía del cuerpo de los animales (Zhao et al., 2016). Los valores se expresaron con relación al peso metabólico (kcal/kg PV0.75).
Estimación de la producción de calor
La producción de calor (PC) se estimó por deducción de la energía metabolizable (EM) consumida en la dieta y la retención de energía (RE) en el cuerpo animal, sujeta a la ecuación del balance de energía (PC = EM RE), expresada por unidad de peso metabólico (kcal/kg PV0.75). El requerimiento de energía metabolizable para mantenimiento se estimó por regresión lineal simple de la retención de energía sobre el consumo de energía metabolizable, como la producción de calor en retención cero de energía, dado por el intercepto x de la regresión (Marcondes et al., 2013). El requerimiento de energía metabolizable para ganancia de peso vivo (EMg) se estimó por regresión lineal simple de la energía metabolizable destinada para la ganancia de peso vivo (kcal/día) sobre la ganancia de peso corporal (g/día); para lo cual, la energía metabolizable consumida en la dieta (EMc, kcal/día) se fraccionó en energía metabolizable destinada al mantenimiento (EMm) y energía metabolizable destinada a la ganancia de peso vivo (EMg). La pendiente de la ecuación corresponde al requerimiento de energía metabolizable de ganancia de peso vivo (Luo et al., 2004).
Análisis Estadístico
Los datos de la energía metabolizable de la dieta se analizaron mediante el análisis de varianza en un diseño Cuadrado Latino 4 x 4 por especie, utilizando el paquete estadístico VassarStats.net de acceso libre (Senthilkumar, 2006), según el siguiente modelo aditivo lineal fijo Yijk=m+ri+gj+tk+eijk; donde Yijk es la variable dependiente; m, media general; ri, variación entre etapas (filas); gj, variación entre animales (columnas); tk, variación entre niveles de consumo (tratamientos); eijk, variación entre observaciones (error).
La producción de calor de los animales en retención cero de energía se estimó por regresión lineal simple y su análisis de varianza (Kumari y Yadav, 2018), cuyos parámetros se analizaron mediante la prueba t, sujeto al siguiente modelo: Y= b0+ b1X + eij; donde Yij, variable dependiente; X, variable independiente; b0, intercepto de la regresión; b1, pendiente de la regresión; y eij, error de la regresión. Todos los análisis se realizaron a un nivel de significancia de 5% (α=0.05).
RESULTADOS Y DISCUSIÓN
Energía Metabolizable de la Dieta
El consumo de materia seca de la dieta incrementó en relación directa con el nivel de consumo, tanto en llamas como en alpacas (p<0.01). Los consumos máximos de materia seca registrados en llamas y alpacas alimentadas en el nivel ad libitum fueron de 1.968 kg/día y 1.211 kg/día (Cuadro 3), los mismos que corresponden a 1.87 y 1.92% del peso vivo, respectivamente. Ninguna llama ni alpaca logró consumir la cantidad prevista de alimento para consumo ad libitum. Estos resultados están en acuerdo con los reportes de consumo para llamas y alpacas alimentados con forrajes (San Martin y Bryant, 1989).
Cuadro 3 Energía metabolizable de la dieta en llamas y alpacas macho de dos años, en experimento de metabolismo convencional in vivo, con cuatro niveles de oferta de dieta (g MS/kg PV0.75)
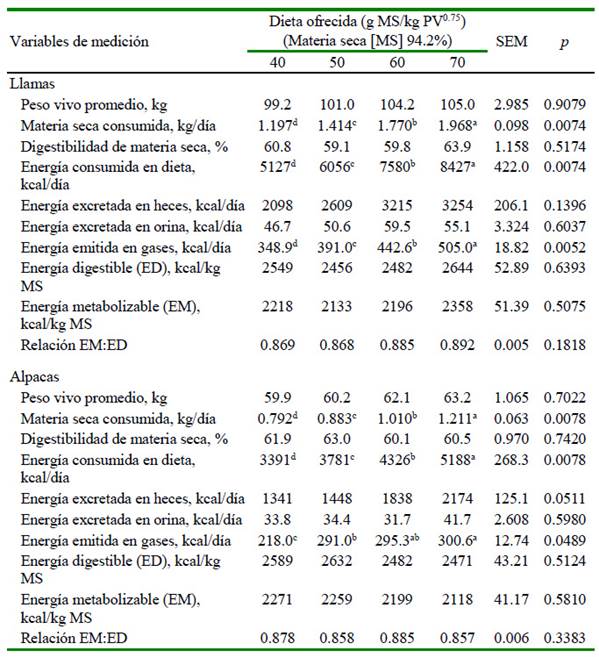
a,b,c,d Letras diferentes dentro de filas indican diferencia significativa (p<0.05)
La digestibilidad de la materia seca del alimento fue similar entre niveles de consumo y entre especies, con un promedio de 60.9 ± 4.6% en llamas y 61.4 ± 3.9% en alpacas (Cuadro 3), los mismos que están en el rango de valores reportados para los distintos forrajes evaluados en llamas y alpacas (San Martin y Bryant, 1989).
El consumo de energía bruta (EB) en la dieta estuvo en relación directa con el nivel de consumo de materia seca (p<0.01). La excreción de energía en las heces mantuvo la misma tendencia que el consumo de EB en la dieta. La excreción de energía en la orina fue similar entre los niveles de consumo, en una proporción <1% de la EB consumida en la dieta, en ambas especies, concordante con los reportes para llamas y alpacas (Robinson et al., 2005; Davies et al., 2007).
Las pérdidas absolutas de energía en gas metano incrementaron en relación directa con el nivel de consumo, en ambas especies (p<0.05), donde las llamas eliminaron más metano que las alpacas (p<0.01); sin embargo, las pérdidas relativas fueron similares entre niveles de consumo y entre especies, con un valor promedio de 6.36 ± 0.99% de la energía bruta consumida en llamas y 6.88 ± 1.55% en alpacas. Estos valores están en el rango de emisiones de metano reportado para alpacas (Pinares-Patiño et al., 2003), alpacas, llamas y camellos (Dittmann et al., 2014), así como vacunos, ovinos y caprinos (Johnson y Ward, 1996).
La energía metabolizable de la dieta fue prácticamente similar entre llamas y alpacas, con un promedio de 2226 ± 206 kcal/kg y 2212 ± 165 kcal/kg de materia seca, respectivamente. Estos resultados son similares al valor reportado para llamas alimentadas con dieta de heno de avena y concentrado peletizado (Carmean et al., 1992).
La relación entre la energía metabolizable y la energía digestible (EM:ED) fue similar entre los niveles de consumo de la dieta y entre especies de animales (Cuadro 3), con un valor promedio de 0.878 ± 0.018 para llamas y 0.870 ± 0.026 para alpacas, las mismas que son mayores al valor reportado para vacunos (Sundstøl, 1993) y ovinos (Armstrong, 1964), pero similares a los valores para cabras (Teixeira et al., 2019).
Requerimientos de Energía Metabolizable de Mantenimiento
Retención de energía
Los animales iniciaron la fase de alimentación del experimento de sacrificio comparativo con pesos similares entre grupos, tanto en llamas como en alpacas, pero finalizaron con pesos diferentes (p<0.05), con ganancias de peso en relación directa con el nivel de consumo de la dieta. Algunos animales alimentados en el nivel de mantenimiento perdieron peso, mientras que todos los animales de los demás niveles ganaron peso, con la mayor ganancia en el nivel ad libitum (Cuadro 4). La retención de energía mantuvo la misma relación con el nivel de consumo de la dieta (p<0.01).
Cuadro 4 Retención de energía en llamas y alpacas machos de dos años, en experimento de sacrificio comparativo, con cuatro niveles de oferta de dieta (g MS/kg PV0.75). Periodo de alimentación de 55 días (n = 4 animales por grupo)
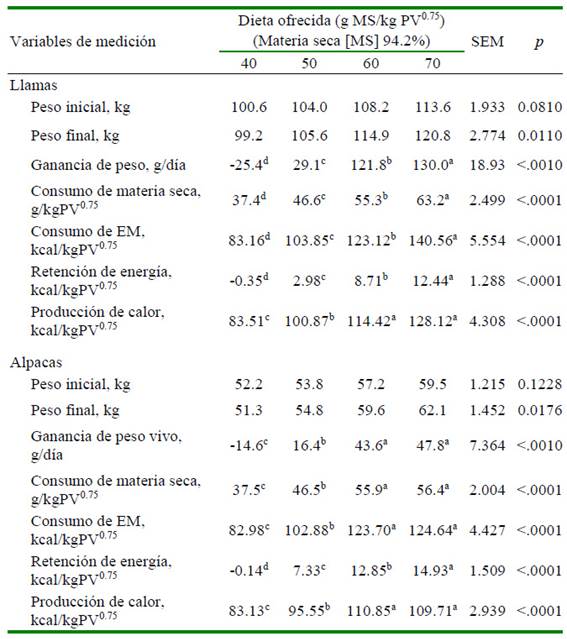
a,b,c,d Letras diferentes dentro de filas indican diferencia significativa.
EM: energía metabolizable
La regresión lineal simple de la retención de energía (kcal/kg PV0.75) sobre el consumo de energía metabolizable (kcal/kg PV0.75), generó las siguientes ecuaciones (Figuras 1 y 2): RE = 0.2264 (± 0.0132) EM -19.564 (± 1.520), R2 0.9543 (Ec.1) llamas RE = 0.3378 (± 0.0121) EM -27.921 (± 1.328); R2 0.9824 (Ec.2) alpacas
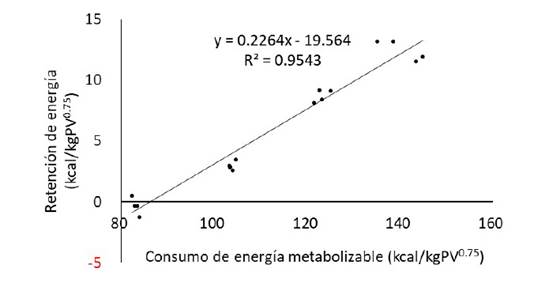
Figura 1 Regresión de la retención de energía (RE) sobre el consumo de energía metabolizable (EM) en llamas macho de dos años (período de alimentación: 55 días; n = 4 animales por grupo)
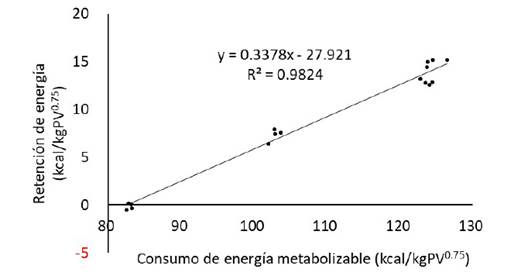
Figura 2 Regresión de la retención de energía (RE) sobre el consumo de energía metabolizable (EM) en alpacas macho de dos años (período de alimentación: 55 días; n = 4 animales por grupo)
Las regresiones fueron significativas (p<0.01), tanto para llamas como para alpacas, con elevadas bondades de ajuste (R2>0.95). Las pendientes de las regresiones corresponden a las eficiencias de uso de la energía metabolizable para la retención o ganancia, siendo kg=0.2264para llamasykg = 0.3378 para alpacas, las mismas que son relativamente bajas con relación a las de otras especies de rumiantes (Regadas et al., 2011). Las diferencias, km = 0.7736 para llamas y km = 0.6622 para alpacas, expresan las eficiencias parciales de uso de la energía metabolizable para la producción de calor por encima del mantenimiento, las mismas que están en los rangos de eficiencia de uso de EM para mantenimiento y ganancia reportados para experimentos de sacrificio comparativo (Johnson et al., 2003).
Producción de calor
La producción de calor varió también en relación directa con el consumo de energía metabolizable (p<0.01), desde 83.51 kcal/ kg PV0.75 en el nivel de mantenimiento hasta 128.12 kcal/kg PV0.75 en el nivel ad libitum en llamas; y desde 83.13 kcal/kg PV0.75 hasta 109.71 kcal/kg PV0.75, respectivamente, en alpacas (Cuadro 4). Estos resultados evidencian que las llamas y las alpacas producen calor en forma activa para mantener su temperatura corporal relativamente estable a fin de hacer frente a la adversidad térmica de la gran altitud (Riek et al., 2019). La cobertura de fibra es una ventaja que posibilita a estas especies realizar la termorregulación y mantener la homeotermia; sin embargo, esa capacidad puede verse alterada por la práctica de la esquila, incrementando su demanda de energía para la producción de calor (Gerken, 2009).
Al respecto, los reportes sobre la termorregulación en camélidos que viven en las grandes altitudes muestran datos sorprendentes sobre sus habilidades para el manejo de las condiciones térmicas adversas del medio ambiente. Los camélidos tienen ventanas térmicas que les permiten modular la disipación de calor (Gerken, 2009), por lo que los animales amanecen con temperaturas bajas y atardecen con temperaturas altas, con un gasto de energía notablemente bajo con relación a otros herbívoros (Riek et al., 2019). Es posible que la llama se comporte como el camello dromedario, especie que alterna periodos de homeotermia y poiquilotermia (Bouâouda et al., 2014), con una temperatura corporal que puede variar desde 34 °C al amanecer hasta 41 °C al atardecer (Gaughan, 2011), haciéndolo diferente al ovino (daSilva y Minomo, 1995) y al bovino (Gaughan et al., 2010), cuando las condiciones térmicas son adversas. Existe limitada información sobre estos aspectos relacionados a la termorregulación y a la actividad física en camélidos sudamericanos, las mismas que son los componentes adicionales necesarios para realizar los ajustes de los requerimientos energéticos de mantenimiento.
A partir de las ecuaciones de regresión (Ec. 1 y 2), en retención cero de energía (RE = 0), la energía metabolizable (EM) resulta del cociente entre el intercepto y la pendiente de la ecuación, 0 corresponde al intercepto X de la ecuación, cuyo valor es 86.4 para llamas y 82.6 para alpacas. La ecuación del balance de energía indica también que en retención cero de energía, la energía metabolizable es igual a la producción de calor, EM = PC + 0 (Derno et al., 2005), siendo por tanto este valor el requerimiento de energía metabolizable de mantenimiento (EMm), expresado por las siguientes ecuaciones:
EM m, kcal/día = 86.4 kcal/kg PV0.75 (Ec. 3) llamas
EM m, kcal/día = 82.6 kcal/kg PV0.75 (Ec. 4) alpacas
Los resultados son similares al valor reportado para llamas (Carmean et al., 1992), pero mayores que los reportados por Engelhardt y Schneider (1977) y Nielsen et al. (2014); y menores en aproximadamente 27% a los requerimientos de mantenimiento de las otras especies de rumiantes, tales como vacunos (Valente et al., 2013); ovinos (Zhao et al., 2016) y caprinos (Resende et al., 2018), evidenciando que las llamas y las alpacas tienen tasas metabólicas basales mucho más bajas y requerimientos de energía metabolizable de mantenimiento excepcionalmente bajos (Riek et al., 2019).
Las llamas alimentadas en nivel ad libitum alcanzaron hasta 120 kg de peso vivo, mientras que las alpacas hasta 60 kg de peso vivo.A partir de los valores encontrados (Ec. 3 y 4), sus demandas de energía metabolizable corresponden a 3133 kcal/día y 1781 kcal/ día, respectivamente. Dado que el contenido de energía metabolizable de la dieta fue de 2.226 kcal/g de materia seca para llamas y 2.212 kcal/g para alpacas, las cantidades mínimas estimadas de materia seca necesarias para mantener la vida de estos animales son de 38.8 g/kgPV0.75 y 37.3 g/kgPV0.75, respectivamente, considerando una dieta conformada por una mezcla de heno de avena y heno de alfalfa, procesados mecánicamente. Las cantidades pueden variar con un cambio en la dieta, puesto que la especie de planta en la dieta tiene efecto sobre la composición química y el valor energético de la dieta (Uslu et al., 2018).
La otra forma de energía para mantenimiento corresponde a la producción de calor en consumo cero de alimento, conocida como producción de calor en ayuno (PCA), la misma que expresa el requerimiento de energía neta de mantenimiento (ENm). El valor depende de la eficiencia (km) de uso de la energía metabolizable para la producción de calor, PCA = km x EM m (Garrett, 1980). Las ecuaciones (Ec. 1 y 2) evidencian un valor km, 0.7736 para llamas y 0.6622 para alpacas, por lo que ENm está dada por las siguientes ecuaciones:
EN m, kcal/día = 66.8 kcal/kgPV0.75 (Ec. 5) llamas
EN m, kcal/día = 54.7 kcal/kgPV0.75(Ec. 6) alpacas
Los valores son similares a los obtenidos para llamas por Carmean et al. (1992) y Nielsen et al. (2014); sin embargo, son relativamente más bajos que los reportados para las otras especies de rumiantes, tales como vacunos (Valente et al., 2013), ovinos (Zhao et al., 2016) y caprinos (Resende et al., 2018), evidenciando la menor demanda de energía de estos camélidos, quizá como un mecanismo de evolución y adaptación a las condiciones térmicas adversas de la gran altitud (Wu et al., 2014).
Requerimientos de Energía Metabolizable de Ganancia de Peso
La ganancia de peso vivo de los animales fue diferente entre niveles de alimentación, tanto en llamas como en alpacas (p<0.01), incrementando en una relación directa con el nivel de consumo. Los grupos alimentadosennivel próximo al mantenimiento perdieron peso, mientras que los alimentados en niveles mayores ganaron peso, con la mayor ganancia en el nivel ad libitum. La retención de energía mostró la misma tendencia (Cuadro 4).Apartir de la ecuación del balance de energía, la diferencia entre la energía consumida enel alimentoylaenergía utilizadapara el mantenimiento corresponde a la energía destinada para la ganancia de peso.
La regresión lineal de la energía retenida (kcal/día) sobre la ganancia de peso (g/ día), generó las siguientes ecuaciones:
y = 9.2x + 339, (R2 0.763) (Ec.7) para llamas y = 11.4x + 277, R2 0.768) (Ec. 8) para alpacas donde: y, energía metabolizable requerida para la ganancia de peso (kcal/día); x, ganancia de peso de los animales (g/día).
Las regresiones fueron significativas (p<0.01), con elevadas bondades de ajuste (R2>0.75). La pendiente de la regresión expresa la contribución de la variable independiente x en el cambio de la variable dependiente y; es decir, el incremento que se produce en la variable y cuando la variable x aumenta en una unidad (Schneider et al., 2010). En el presente caso, se requieren 9.2 kcal de energía metabolizable por cada gramo de ganancia de peso vivo en llamas, y 11.4 kcal en alpacas. Estos resultados son mucho mayores a los valores reportados para ovinos, caprinos y vacunos (Salah et al., 2014), indicando que las llamas y alpacas alimentadas con dietas de forrajes utilizan mayores cantidades de energía metabolizable por cada unidad de ganancia de peso vivo, evidenciando su menor eficiencia para convertir la energía del alimento en carne, con relación al ovino productor de carne (Zhao et al., 2016).
Es posible que cualquier cambio en la alimentación o el manejo de los animales pueda hacer variar los valores de la pendiente de la regresión, puesto que el tipo de alimento y el sistema de manejo juegan roles importantes en la ganancia de peso de los animales rumiantes (Xu et al., 2017).Así por ejemplo, a 4200 m de altitud, donde la temperatura del aire varía desde -5 °C (junio) hasta 14 °C (noviembre), las mayores ganancias de peso se observan en llamas de dos años en la época lluviosa y con pastos cultivados (García et al., 2002).
La condición ambiental es el otro factor que puede alterar también la pendiente de las ecuaciones, dado que está estrechamente relacionada con el mantenimiento. Los modelos de crecimiento para llamas en los distintos contextos evidencian amplias diferencias en peso al nacimiento y peso adulto de los animales. Los datos extraídos para efectos comparativos, desde el nacimiento hasta los siete meses de edad, muestran pesos menores para animales de puna seca (Canaza-Cayo et al., 2015), mayores para los de puna húmeda (Apaza y Quispe, 2016), y mucho mayores para animales de altitudes cercanas al nivel del mar (Smith et al., 1992), evidenciando los efectos que tiene la relación entre las demandas de energía y el consumo de alimentos, siendo este último el factor clave para el rendimiento animal (Zereu, 2016), debido a que define la retención de energía (Murray, 1991), cuyas consecuencias se manifiestan en el balance de energía y la condición corporal, con estados de caquexia en ámbitos de gran altitud y obesidad en espacios de baja altitud (Van Saun, 2006).
La actividad física también puede alterar la referida pendiente de las ecuaciones, puesto que esta función consume buena parte de la energía metabolizable, pudiendo ser de 25 a 100% mayor que en animales en confinamiento (Shinde y Karim, 2007); sin embargo, este aspecto no fue considerado en el análisis, puesto que los animales estuvieron en confinamiento, con mínima actividad física.
Finalmente, el contraste de las regresiones de la retención de energía sobre el consumo de energía metabolizable entre llamas y alpacas fue significativamente diferente (p<0.01), indicando que los requerimientos energéticos derivados de estas ecuaciones son independientes.
CONCLUSIONES
El requerimiento de energía metabolizable de mantenimiento (EMm) fue diferente entre especies, 86.4 kcal/kg PV0.75 y 82.6 kcal/kg PV0.75, para llamas y alpacas, respectivamente, con sus correspondientes eficiencias parciales (km) de 0.77 y 0.66. El requerimiento de energía metabolizable de ganancia de peso (EMg) fue de 9.2 y 11.4 kcal/g de ganancia de peso vivo, con sus eficiencias parciales (kg) de 0.23 y 0.34, respectivamente. Se concluye que los requerimientos de EMm de llamas y alpacas en gran altitud son independientes entre sí, y menores en aproximadamente 27% con relación a los reportados para rumiantes.

