










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista Peruana de Biología
versão On-line ISSN 1727-9933
Rev. peru biol. vol.26 no.1 Lima jan./mar. 2019
http://dx.doi.org/10.15381/rpb.v26i1.15910
TRABAJOS ORIGINALES
Composición y fluctuación poblacional de la araneofauna en el algodonero de la Universidad Nacional Agraria La Molina, Lima, Perú
Composition and population fluctuation of the spider community in a cotton field at Universidad Nacional Agraria La Molina, Lima, Peru
Laura Cruz*;1, Diana Silva2 y Clorinda Vergara3
1Universidad Nacional Mayor de San Marcos, Lima-Perú.
2Universidad Nacional Mayor de San Marcos, Museo de Historia Natural, Lima, Perú.
3Universidad Nacional Agraria La Molina, Museo de Entomología Klaus Raven Büller, Lima-Perú.
Resumen
En el presente trabajo, se evaluó la araneofauna durante todo el periodo fenológico del cultivo de algodón, utilizando dos técnicas de muestreo (búsqueda directa y trampas de caída). Se registraron 2304 individuos, agrupados en 50 especies y 18 familias. Las arañas estuvieron presentes durante todo el periodo del cultivo, relacionándose positivamente a su desarrollo fenológico, y donde las prácticas agronómicas, la temperatura y humedad no fueron influyentes en su abundancia, excepto por la cosecha, actividad que sí influenció negativamente sus poblaciones.
Palabras clave: Arañas; Gremios de arañas; Diversidad de artrópodos; Prácticas agronómicas; Cultivo de algodón.
Abstract
In this work, the spider community was assessed during the whole phenological period of a cotton crop using two sampling techniques (direct search and pitfall traps). 2304 individuals were registered, grouped into 50 species and 18 families. The spiders were present during the whole crop’s period; positively related to its phenological development; agronomic practices, temperature and humidity were not influential in their abundance, except for harvest, which negatively influenced their population.
Keywords: Spiders; Spider guilds; Arthropods diversity; Agronomic practices; Cotton crop.
Introducción
Comúnmente los cultivos de algodón son frecuentemente infestados por plagas (Beingolea 1959), por lo que también son intensamente sometidos a plaguicidas (CCI 2007). Aunque los principales plaguicidas utilizados en el algodón son relativamente baratos y de amplio espectro, estos también son muy perjudiciales para los insectos benéficos, y además, su poder residual se convierten en un problema importante para el ambiente (CCI 2007). Pero, en Perú la naturaleza de sus valles costeros permite que la mayoría de las plagas del algodón puedan ser controladas eficientemente por sus enemigos naturales (Beingolea 1959), y se estima que de no ser así sería difícil mantener la producción de algodón (Herrera 1987).
Las arañas constituyen uno de los grupos depredadores más abundantes de los ecosistemas terrestres, considerados polífagos o generalistas (Bristowe 1971), por lo que no podrían ser eficientes controladores de una plaga determinada (Debach & Rosen 1991). Sin embargo, existen resultados discrepantes de su eficiencia en diversos cultivos, siendo utilizados exitosamente como agentes de biocontrol en dos tipos de agroecosistemas:huertos (principalmente de manzano) en Israel, Europa, Australia y Canadá, y arroz en Asia (Mansour et al. 1980, Marc 1993, Zhao 1993, Maloney et al. 2003). Marc y Canard (1997) redefinieron la importancia de las arañas en el agroecosistema, especificando que si se toma en cuenta sus estrategias de caza y localización en la vegetación, pueden considerarse como depredadoras especialistas. Así mismo indican que no todas las especies de arañas son útiles contra una plaga particular, pero considerando su diversidad podrían ser importantes para controlar diversas plagas.
En el Perú, Aguilar (1965, 1968, 1974, 1975, 1976, 1977, 1978a, 1978b, 1979a, 1979b, 1988) realizó importantes contribuciones al conocimiento de la araneofauna en cultivos de algodón a lo largo de la costa peruana, resaltando su importancia (diversidad, preferencia de localización en el cultivo y casos particulares de depredación sobre plagas), y enfatizando la necesidad de continuar estas investigaciones. Sin embargo, posteriormente no han sido desarrollados trabajos que realicen el seguimiento continuo y detallado de la araneofauna durante toda la fenología del cultivo del algodón, por lo tanto, aún no se conoce en detalle las interacciones entre el desarrollo del cultivo y las prácticas agronómicas sobre la comunidad de arañas.
Teniendo en cuenta a las arañas como un componente importante de la fauna benéfica del cultivo, y tratando de promover el interés para que se realicen más investigaciones de este grupo de artrópodos en otros agroecosistemas, el objetivo de esta investigación fue evaluar la diversidad y fluctuación de la abundancia de la comunidad de arañas en los algodonales de la Universidad Nacional Agraria La Molina en relación con las condiciones climáticas, fenológicas y prácticas agronómicas realizadas durante el desarrollo del cultivo.
Material y métodos
Área de estudio.-
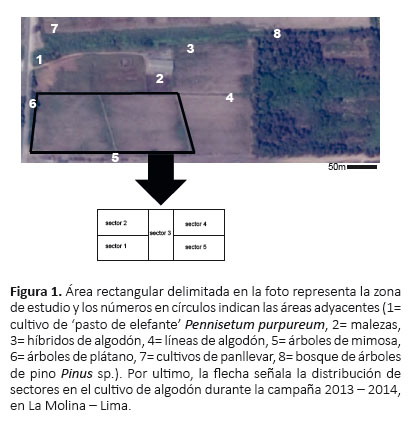
La investigación se realizó en el algodonero variedad Tangüis (cultivar UNA N°1) en la Universidad Nacional Agraria La Molina, Lima (12°05'06.5"S, 76°56'42.7"W, 238 m de altitud), en un campo con una extensión de 1.5 ha. Las áreas colindantes al cultivo de algodón fueron: un cultivo de pasto de elefante Pennisetum purpureum, un área sin cultivar cubierta de malezas, un área destinada al cultivo de híbridos y líneas de algodón, y algunos árboles de plátano (Musa paradisiaca) y mimosa (Acacia longifolia) como bordes del campo (Fig. 1); además cerca de la zona de estudio se encontraba un bosque de árboles de pino Pinus sp. y otros cultivos de panllevar.
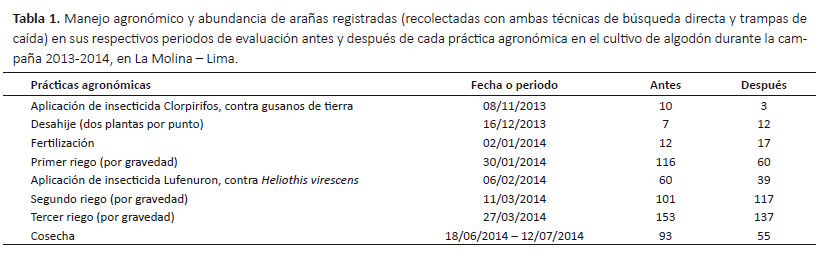
Las evaluaciones se ejecutaron con la ayuda de dos personas en un solo día y por la mañana (8:00 – 12:00 h), cada siete días, de noviembre del 2013 a julio del 2014 (nueve meses), tiempo que corresponde al periodo fenológico del cultivo. Las condiciones meteorológicas fueron registradas por la Estación Meteorológica Alexander Von Humboldt (UNALM) y el manejo agronómico se llevó a cabo de acuerdo a lo establecido por el Programa de Algodón de UNALM (Veramendi & Lam 2011) (Tabla 1).
Técnicas de muestreo.- El campo se dividió en cinco sectores de 3000 m2 cada uno (Fig. 1) y se aplicaron dos técnicas de recolecta: búsqueda directa y trampas de caída (Tabla 2). La secuencia cronológica que se siguió para evaluar cada uno de los sectores fue la siguiente: sector 5, sector 4, sector 3, sector 2 y sector 1.

- Técnica de búsqueda directa: Se efectuó de forma manual y con paraguas entomológico. De forma manual, en cada sector se evaluaron 20 plantas escogidas al azar en cada periodo de muestreo, siendo un total de 100 plantas evaluadas. La evaluación consistió en el examen minucioso de toda a planta (hojas, botones florales, flores y bellotas). Con paraguas entomológico, en cada sector se evaluaron cuatro plantas escogidas al azar en cada periodo de muestreo, teniendo un total de 20 plantas evaluadas. Ésta se realizó sacudiendo vigorosamente la planta (entre 10 a 15 batidas) sobre una manta blanca (0.75 x 1 m) extendida sobre el suelo. En total 120 plantas fueron evaluadas por la técnica de búsqueda directa.
- Técnica de trampas de caída: En cada sector se instalaron dos trampas de caída, colocadas equidistantes sobre los surcos. En total se instalaron 10 trampas, éstas consistieron en envases de plástico (9 cm de altura, 9 cm de diámetro y 450 ml de capacidad), a los cuales se les colocó 2/3 de agua, dos cucharaditas de sal común al ras para evitar la descomposición de las arañas recolectadas y shampoo inodoro e incoloro para romper la tensión superficial. Con la ayuda de una lampa pequeña las trampas fueron instaladas al ras del suelo y cambiadas cada siete días, colocando el contenido recolectado en una bolsa plástica, con sus respectivos datos de recolecta.
El material recolectado fue llevado al laboratorio del Museo de Entomología Klaus Raven Büller – UNALM, en donde se acondicionó el material para su reconocimiento y curación final. Las arañas se preservaron con alcohol al 75%.
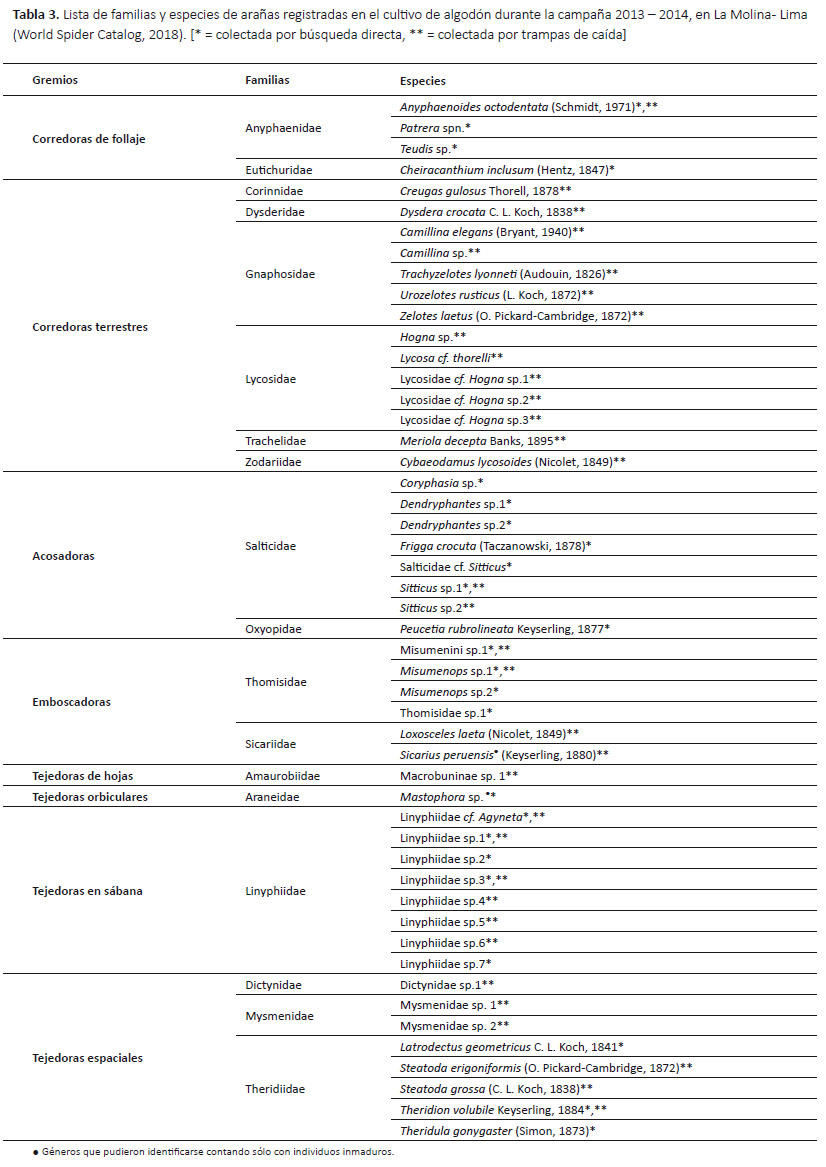
Identificación de arañas.- Con la ayuda de un microscopio estereoscopio y claves y publicaciones específicas se identificaron las arañas hasta el nivel de especie o morfoespecie. En algunos casos a los machos, se les cortó el pedipalpo izquierdo (en vista dorsal) entre la unión de la tibia y patela o de la patela y fémur (dependiendo de la presencia de alguna apófisis). En algunas hembras, se disectó el epiginio, el cual se sumergió en agua destilada para enjuagarse, y luego se colocó en una solución con proteasa (1.5 mL de agua destilada más la pastilla enzimática) de 8 a 10 horas a 37 °C (para la limpieza de las estructuras internas). Por último, cada estructura se preservó en un microvial con alcohol al 75% junto a la araña. La nomenclatura asignada sigue la última actualización de World Spider Catalog (2018).
Asignación de gremios.- Las familias registradas fueron agrupadas en gremios basados en la clasificación de Uetz et al. (1999), una clasificación específica para agroecosistemas, el cual agrupó las familias de arañas que se encuentran en diversos tipos de cultivos (incluyendo el algodón) en ocho gremios: corredoras de follaje, corredoras terrestres, emboscadoras, acosadoras, tejedoras de hojas, tejedoras en sábana, tejedoras espaciales y tejedoras orbiculares.
Análisis de datos.-Los siguientes análisis ejecutados se realizaron en base a la hipótesis de que la diversidad y abundancia de arañas están influenciadas por las condiciones climáticas, fenológicas y prácticas agronómicas durante todo el periodo del cultivo.
Se realizó un análisis cuantitativo (adultos y juveniles) determinado por la suma de las arañas recolectadas y sus distribuciones porcentuales. La sección de análisis general se realizó mediante la suma de las arañas recolectadas de todos los sectores, todas las técnicas y todas las fechas de evaluación. La fluctuación poblacional de arañas se realizó mediante la suma de las arañas recolectadas de todos los sectores, todas las técnicas y todas las fechas de evaluación por cada mes, además de mostrar las arañas registradas antes y después de cada práctica agronómica, la cual se realizó sumando las arañas recolectadas de todos los sectores y todas las técnicas evaluadas de aquella evaluación previa y posterior a cada práctica agronómica. Por último, la fluctuación poblacional de arañas a nivel de familias se realizó mediante la suma de las arañas recolectadas de todos los sectores, todas las técnicas y todas las fechas de evaluación por cada mes y cada familia de arañas (considerando solo las familias más abundantes).
La sección de análisis por sectores se realizó mediante la suma de las arañas recolectadas de todas las técnicas y fechas de evaluación por cada sector; la fluctuación poblacional de arañas se realizó mediante la suma de las arañas recolectadas de todas las técnicas y todas las fechas de evaluación por cada mes y cada sector. Y por último, la sección de análisis por técnicas de evaluación se realizó mediante la suma de las arañas recolectadas de todos los sectores y todas las fechas de evaluación por cada técnica. Además, se evaluó la correlación lineal entre la abundancia de arañas con la temperatura y humedad relativa con el programa Past 2.17c.
Se evaluó la composición de especies (adultos y juveniles) y la diversidad (adultos) tomando en conjunto las arañas recolectadas de todas las técnicas y fechas de evaluación, con los siguientes índices y estimadores: Riqueza específica, Índice de dominancia de Simpson, Índice de diversidad de Shannon-Wiener, Chao 1, Jacknife de 1° orden y Curvas de acumulación de especies. Los estimadores Chao 1, Jacknife de 1° orden y las Curvas de acumulación de especies se evaluaron para toda la zona de estudio (análisis general) y se utilizó el programa Estimates 9.1.0; y la riqueza específica, los índices de dominancia de Simpson y de diversidad de Shannon-Wiener se evaluaron por sectores (análisis por sectores) y se utilizó el programa Past 2.17c.
Por último, se realizó un Análisis de clúster jerárquico con el algoritmo de Método de Ward para evaluar la relación de abundancia de arañas entre sectores, dos Análisis de clúster jerárquico con el algoritmo de Grupo pareado con las mediciones de similaridad de Morisita y Raup-Crick para evaluar la relación de diversidad de las especies de arañas entre sectores, y un Análisis de clúster jerárquico con el algoritmo de Grupo pareado con la medición de similaridad de Morisita para evaluar la relación de diversidad de los gremios de arañas entre sectores, y se utilizó el programa Past 2.17c.
Resultados
Análisis general
Abundancia de arañas.- Se recolectaron un total de 2304 individuos durante los nueve meses de estudio, constituyendo una densidad poblacional de 0.2 arañas m-2, donde 15.06% fueron adultos y 84.94% juveniles, con una proporción de juveniles/adultos = 5.64. De los adultos recolectados, el 61.38% fueron machos y el 38.62% hembras, con una proporción de machos/hembras= 1.59. Las familias más abundantes fueron Thomisidae (40.71%), Anyphaenidae (23.35%) y Theridiidae (12.15%). Las especies más abundantes fueron Misumenops sp.1 (26.80%), Zelotes laetus (15.85%) y Theridion volubile (13.54%). Las correlaciones lineales entre la abundancia de arañas y la temperatura (R= 0.059, P=0.748), y la abundancia de arañas y la humedad relativa (R= -0.012, P= 0.948) resultaron no significativas, por lo que estos factores climáticos no guardaron relación con la abundancia de arañas.
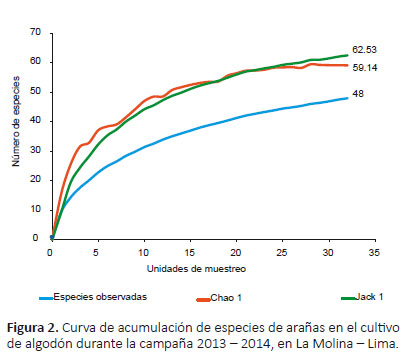
Riqueza de arañas.- Se identificaron 50 especies, agrupadas en 29 géneros y 18 familias (Tabla 3). Las familias con mayor riqueza fueron Linyphiidae (ocho especies) y Salticidae (siete especies). Diversidad de arañas. Los estimadores Chao 1 y Jack 1 obtuvieron un valor de 59 y 63 especies cada uno, e indicaron que se tiene el 81% y 76% de las especies estimadas respectivamente. La curva de acumulación de
especies
Diversidad de arañas.- Los estimadores Chao 1 y Jack 1 obtuvieron un valor de 59 y 63 especies cada uno, e indicaron que se tiene el 81% y 76% de las especies estimadas respectivamente. La curva de acumulación de especies (Fig. 2) no alcanzó la asíntota, observándose en crecimiento, por lo que el cultivo tiene una mayor riqueza esperada de arañas que la registrada.
Gremios de arañas.- Se registraron ocho gremios de arañas según la clasificación de Uetz et al. (1999) (Tabla 3). El gremio más abundante fue el de las emboscadoras (40.84%) (Tabla 4) y el gremio con mayor riqueza fue el de las corredoras terrestres (seis familias y 14 especies) (Tabla 3). En el gremio de las corredoras de follaje la familia más abundante fue Anyphaenidae (98.53%), y en el de las corredoras terrestres la familia más abundante fue Lycosidae (48.70%). En los gremios de las acosadoras y emboscadoras las familias más abundantes fueron Salticidae (95.60%) y Thomisidae (99.68%) respectivamente. En los gremios de las tejedoras espaciales la familia más abundante fue Theridiidae (98.25%). Los gremios de las tejedoras de hojas, tejedoras en sábana y tejedoras orbiculares estuvieron representadas por una familia cada una.
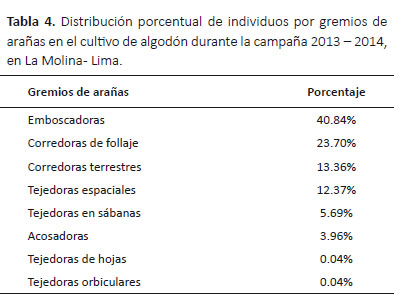
Fluctuación poblacional de arañas.- Las arañas estuvieron presentes durante todo el ciclo de desarrollo del algodonero (nueve meses) (Fig. 3, 4), y se observó que sus poblaciones disminuyeron en los periodos de aplicación de los insecticidas, sin embargo, nunca dejaron de estar presente (Tabla 1, 6).
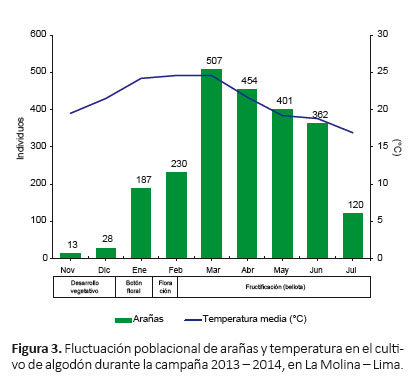
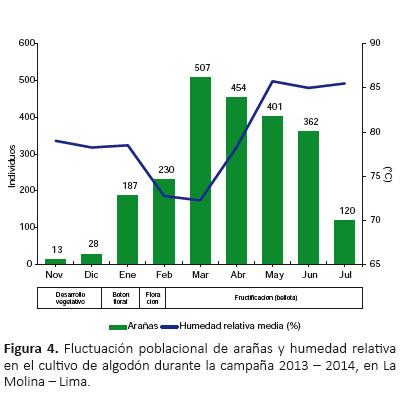
Durante la primera etapa fenológica (desarrollo vegetativo), a pesar de la colonización de la araneofauna al inicio de la campaña, la población de arañas disminuyó rápidamente luego de la primera aplicación de insecticida (Clorpirifos), registrándose en la última evaluación antes de la aplicación 10 individuos y luego de ésta solo tres individuos (Tabla 1), recolectándose en total solo 13 individuos en noviembre (Fig. 3, 4). Iniciando la segunda etapa fenológica (formación de botón floral) se realizó el desahije, encontrándose en la última evaluación antes de esta práctica siete individuos y luego de ésta 12 individuos, registrándose en total 28 individuos en diciembre (Tabla 1, Fig. 3, 4). Poco tiempo después se realizó la fertilización, donde también se observó un incremento de la población, registrándose en la última evaluación antes de esta práctica 12 individuos y luego de ésta 17 individuos, recolectándose en total 187 individuos en enero, una cifra muy superior a los meses anteriores, formándose así el primer pico de máximo crecimiento (Tabla 1, Fig. 3, 4).
A partir de la tercera etapa fenológica (floración) se realizó el primer riego, después del cual la población de arañas disminuyó rápidamente, registrándose en la última evaluación antes del riego 116 individuos y luego de ésta 60 individuos. Luego, con la segunda aplicación de insecticida (Lufenuron) la población de arañas volvió a disminuir, encontrándose en la última evaluación antes de esta aplicación 60 individuos y luego de ésta 39 individuos (Tabla 1). A pesar de estos decrementos, se registraron en total 230 individuos en febrero (Fig. 3, 4). En la cuarta y última etapa fenológica (fructificación), la población de arañas incrementó nuevamente, alcanzando así el segundo pico de máximo crecimiento, recolectándose en total 507 individuos en marzo (Fig. 3, 4).
En la cuarta y última etapa fenológica (fructificación), la población de arañas incrementó nuevamente, alcanzando así el segundo pico de máximo crecimiento, recolectándose en total 507 individuos en marzo (Fig. 3, 4). En este mes se realizaron los dos últimos riegos; de los cuales, luego del segundo riego la población de arañas se incrementó, registrándose en la última evaluación antes del riego 101 individuos y luego de éste 117 individuos. En contraste, después del tercer riego la población de arañas disminuyó, encontrándose en la última evaluación antes del último riego 153 individuos y luego de éste 137 individuos (Tabla 1).
En los meses de abril y mayo no se realizaron prácticas agronómicas en el cultivo, durante este periodo la población de arañas comenzó a disminuir gradualmente, registrándose en total en abril 454 individuos y en mayo 401 individuos. Durante la cosecha, a mediados de junio y julio, la población de arañas disminuyó, recolectándose en la última evaluación antes de la cosecha 93 individuos y al término de ésta 55 individuos (Tabla 1), registrándose en total 362 individuos en junio y 120 individuos en julio (Fig. 3, 4).
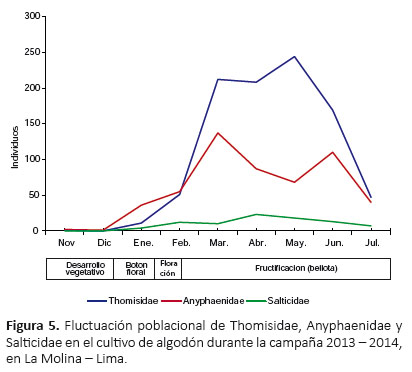
A nivel de familias, en enero se registró el primer pico de máximo crecimiento para Lycosidae; en febrero se estableció el primer y único pico de máximo crecimiento para Gnaphosidae; y en marzo se registró el primer pico de máximo crecimiento para Thomisidae, Anyphaenidae, Linyphiidae y Theridiidae (Fig. 5, 6 y 7). Además, Lycosidae, Thomisidae, Anyphaenidae y Theridiidae registraron un segundo pico de máximo crecimiento, siendo para Lycosidae en abril, Thomisidae y Theridiidae en mayo, y Anyphaenidae en junio (Fig. 5, 6 y 7). A partir de marzo y hasta el final del cultivo Thomisidae fue la familia más abundante por una marcada diferencia con las demás (Fig. 5, 6 y 7).
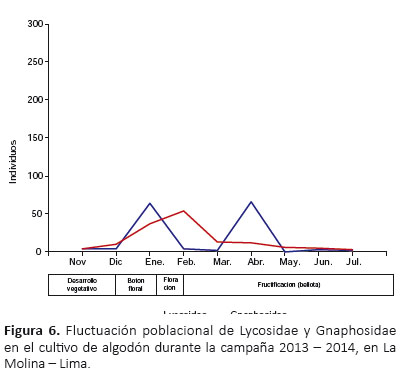
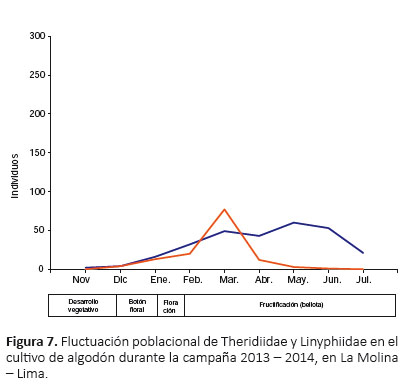
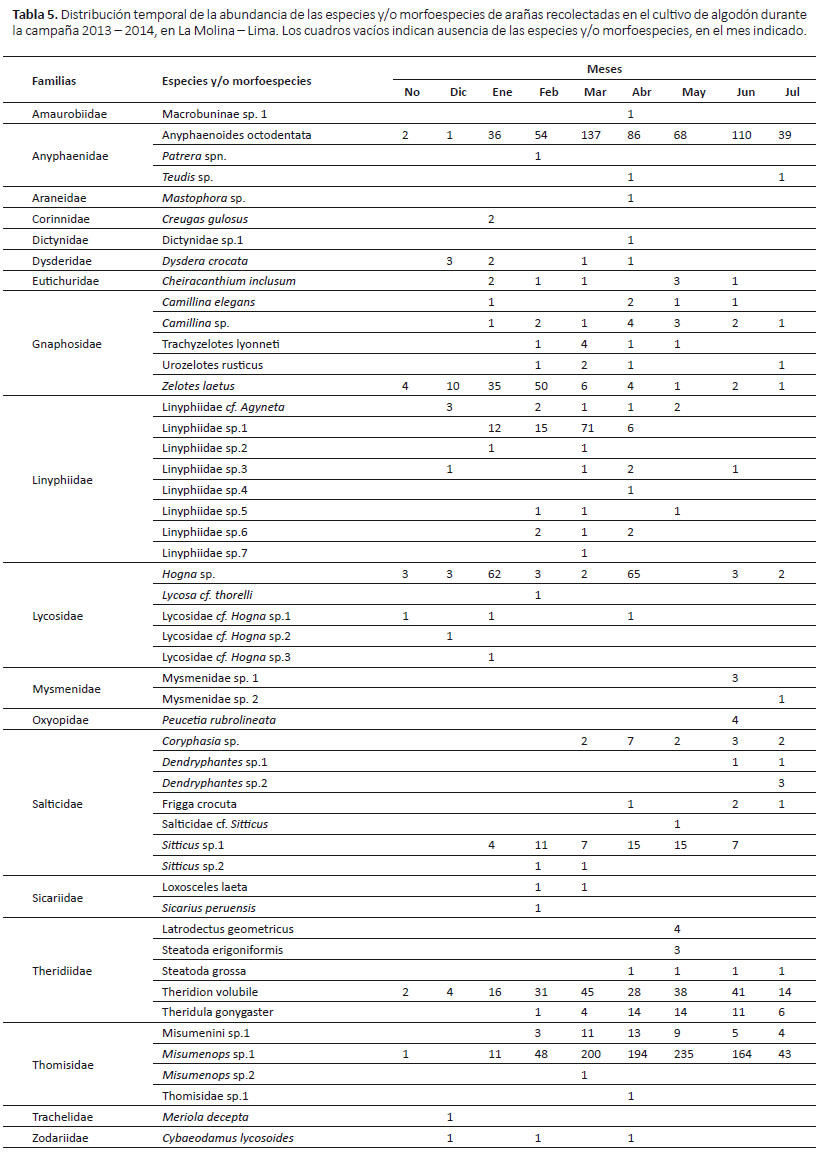
Las especies con mayor distribución temporal fueron Anyphaenoides octodentata, Zelotes laetus y Theridion volubile, las cuales estuvieron presentes en los nueve meses del periodo del cultivo; Misumenops sp. 1 y Hogna sp.estuvieron presentes en ocho de los nueve meses, estando ausentes en diciembre y mayo respectivamente; y Camillina sp. estuvo presente en siete de los nueve meses, estando ausente en noviembre y diciembre (Tabla 5).
Análisis por sectores
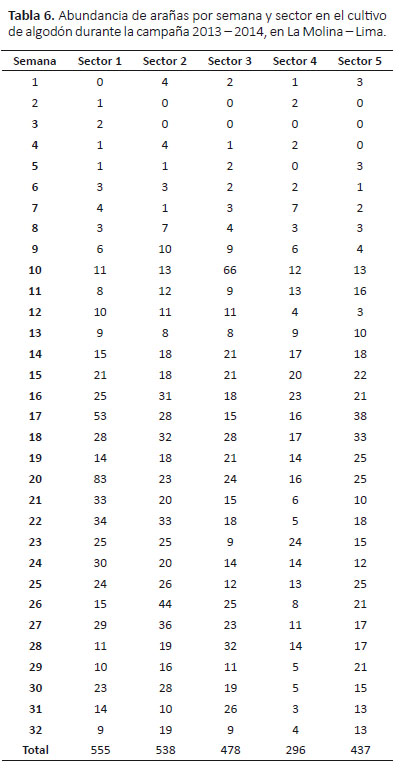
Abundancia de arañas.- La mayor abundancia se registró en el sector 1 (555 individuos) y la menor en el sector 4 (296 individuos) (Tabla 6). El dendrograma mostró dos agrupaciones en relación a su abundancia: sectores 1 y 2 y sectores 3, 4 y 5 (siendo en este último más cercanos los sectores 3 y 5) (Fig. 8).
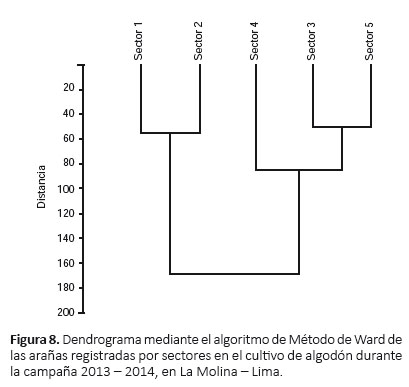
Riqueza de arañas.- La riqueza específica fue mayor en el sector 5 (30 especies), en comparación con los demás sectores, donde la riqueza fue de 25 a 26 especies (Tabla 7).
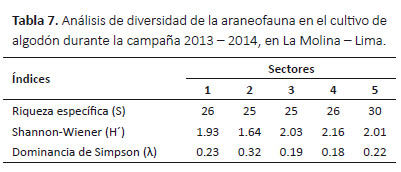
Diversidad de arañas.- El valor más alto del índice de diversidad de Shannon-Wiener fue en el sector 4 (H´= 2.16) y el menor fue en el sector 2 (H´= 1.64); en contraste al índice de dominancia de Simpson, donde el valor más alto del índice fue en el sector 2 (λ= 0.32) y el menor fue en el sector 4 (λ= 0.18) (Tabla 7).
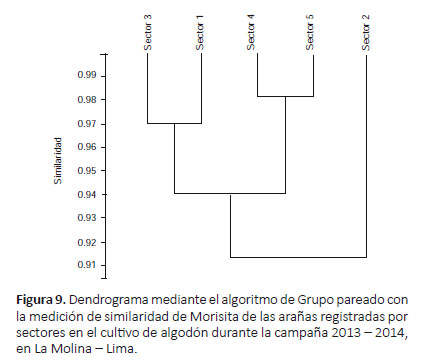
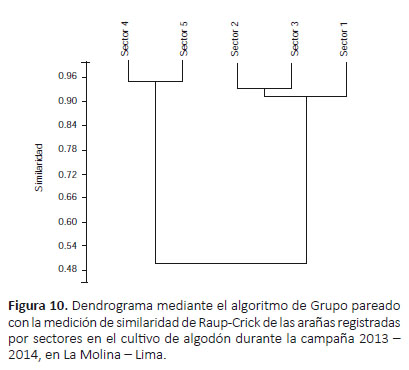
El dendrograma usando la medición de similaridad de Morisita (Fig. 9) registró dos agrupaciones en relación a su diversidad: sectores 1 y 3 y sectores 4 y 5 (donde el sector 2 se distancia desde un inicio de los demás); siendo similar al dendrograma usando la medición de similaridad de Raup-Crick (Fig. 10), que mostró 2 agrupaciones: sectores 1, 2 y 3 (siendo más cercanos los sectores 2 y 3) y sectores 4 y 5.
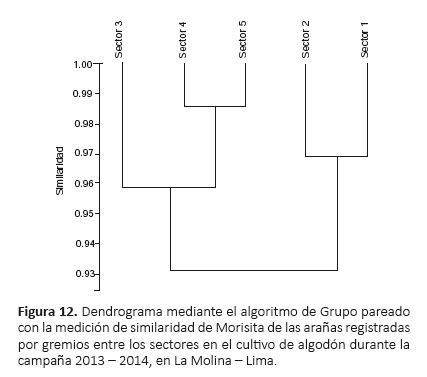
Gremios de arañas.- El gremio de las emboscadoras fue el más abundante en todos los sectores, siendo mayor en el sector 2 (54.12%) (Fig. 11). Los gremios de las tejedoras orbiculares y tejedoras de hojas registraron las menores abundancias, el primero en el sector 3 (0.21%) y el segundo en el sector 4 (0.34%), cada uno encontrado únicamente en ese sector respectivamente (Fig. 11). Asimismo, la mayor riqueza de gremios se registró en los sectores 3 y 4 (siete gremios), mientras que la menor riqueza en los sectores 1, 2 y 5 (seis gremios) (Fig. 11). El dendrograma mostró dos agrupaciones de similaridad: sectores 3, 4 y 5 (siendo más cercanos sectores 4 y 5) y sectores 1 y 2 (Fig. 12).
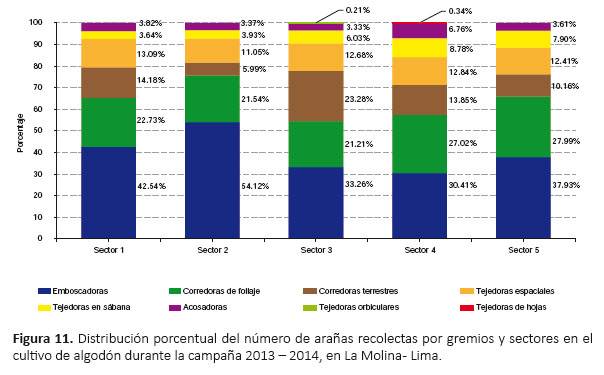
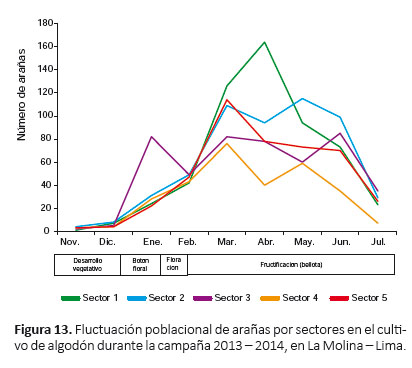
Fluctuación poblacional de arañas.- Las arañas estuvieron presentes en cada uno de los sectores durante todo el periodo fenológico del cultivo (Fig. 13). El primer pico de máximo crecimiento correspondió al sector 3 en enero; luego, el primer pico de máximo crecimiento para los demás sectores y el segundo pico de máximo crecimiento para el sector 3 se registró en marzo (Fig. 13). Los niveles más altos de la población de arañas se registraron en los sectores 1 y 2 (siendo el nivel más alto en abril en el sector 1); en cambio, los niveles más bajos fueron registrados en los sectores 4 y 5, resaltando el sector 4 por su menor abundancia (Fig. 13). En general, hasta febrero, casi todos los sectores tuvieron una población muy similar de arañas (excepto el sector 3), a partir de marzo y hasta junio se registró una diferencia más marcada de la abundancia de arañas entre los sectores (Fig. 13).
Análisis por técnicas de evaluación
Abundancia, riqueza y gremios de arañas.-En la técnica de búsqueda directa el gremio más abundante fue el de las emboscadoras (49.11%), la familia más abundante fue Thomisidae (49.11%), y la especie más abundante fue Misumenops sp.1 (46.70%); las familias con mayor número de especies fueron Salticidae y Linyphiidae (cinco especies cada una). En las trampas de caída el gremio más abundante fue el de las corredoras terrestres (68.44%), la familia más abundante fue Lycosidae (33.3%), y la especie más abundante fue Zelotes laetus (36.42%); la familia con mayor número de especies fue Linyphiidae (ocho especies).
Con la técnica de búsqueda directa se registró una mayor abundancia de arañas (80.60%) que con las trampas de caída. En contraste, la técnica de trampas de caída registró el mayor número de especies (36). Sin embargo, con los dos métodos empleados, se recolectaron el 16% de las especies y cinco gremios (Tabla 3).
Discusión
Análisis general
La densidad poblacional registrada en esta investigación fue similar al nivel mínimo estimado por Aguilar (1988), de 0.4 – 4 arañas m-2 (sacudida de planta) en cultivos de algodón con aplicación de insecticidas a lo largo de la costa peruana; también registraron resultados comunes Skinner (1974), con 0.02 – 0.1 arañas m-2 (método de colecta desconocido), y Dinkins et al. (1970), con 0.1 arañas m-2 (succión con D-vac) en cultivos de algodón con aplicación de insecticidas en Alabama y Misisipi (EEUU) respectivamente. En contraste, esta densidad fue menor a lo registrado por Dean y Sterling (1987), donde se obtuvo 0.8 arañas m-2 en cultivos de algodón sin aplicación de insecticidas (succión con D-vac) en Texas (EEUU), y a Plagens (1983), con 1.3 – 2.3 arañas m-2 en cultivos de algodón con aplicación de insecticidas (Ground cloth) en Arizona (EEUU).
El predominio de arañas juveniles concuerda con lo encontrado por Pérez-Guerrero et al. (2009) y Almada et al. (2012), en cultivos de algodón en España (91.07%) y Argentina (60%) respectivamente. Además, coinciden con otros cultivos y áreas naturales (Almada et al. 2012;Avalos et al. 2013), donde más del 60% fueron individuos juveniles. Esto podría deberse a una constante colonización de las arañas juveniles desde los márgenes del cultivo hacia éste (Almada et al. 2012). La proporción de adultos y juveniles también estaría relacionada a la técnica de recolección empleada, como las trampas de caída, eficientes para capturar arañas adultas, debido a la mayor actividad de éstas en comparación con las juveniles (Uetz & Unzicker 1976, Churchill 1993, Azevedo et al. 2002).
La mayor proporción de machos se asemeja a lo registrado por Dodson et al. (2015) en Misumenoides formosipes (2.37), y contrasta a lo evaluado por Ghavami et al. (2007) (0.17) en cultivos de algodón en Irán. La proporción de sexos también estaría relacionada a la técnica de trampas de caída, ya que los machos son más activos que las hembras, por ello, la alta proporción de machos encontrados en trampas de caída (Álvares et al. 2004), lo que explicaría la mayor abundancia de machos en este estudio. A pesar de ello, se ha señalado que existe una mayor proporción de hembras en la mayoría de arañas adultas sociales y solitarias, y desde el estado de huevo (Gunnarsson & Andersson 1996).
Las familias más abundantes registradas en este trabajo contrastan con las observaciones de Aguilar (1977,1988), quien encontró en mayor abundancia a Anyphaenidae (58%) y en segundo lugar Thomisidae (13%), familia dominante en el presente trabajo. La predominancia de estas familias podría explicarse por sus estrategias de caza y localización en el cultivo, como sugiere Aguilar(1977, 1988), ya que ambas familias al ser ‘cazadoras directas’ tienen mayor ventaja que las ‘tejedoras’. A su vez, Aguilar (1977, 1988) manifestó que Anyphaenidae tuvo una mayor abundancia por ser una familia nocturna, en comparación de Thomisidae que es diurna, entonces se encontraba protegida en sus cubiertas de seda en los órganos de la planta durante el manejo agronómico en el día, lo cual también facilitó su colecta en esta investigación.
La abundancia de las familias también estaría relacionada a las técnicas de evaluación y esfuerzos de muestreo utilizados en cada investigación, siendo cada resultado particular, como en este estudio. Por ejemplo: en Texas, Estados Unidos, Dean et al. (1982) reportaron a Oxyopidae (23%) como la familia más abundante mediante cincotécnicas de colecta: manual, succión con D-vac, redadas, trampas de feromonas y trampas de caída en campos de algodón sin aplicación de insecticidas. En Argentina, Almada et al. (2012) registraron a Thomisidae (84.04%)como la familia más abundante mediante tres técnicas de muestreo: red entomológica de arrastre, paño vertical de un metro y trampas de caída en algodón transgénico y convencional (con y sin control químico). En España, Pérez-Guerrero et al. (2009) encontraron a Miturgidae yThomisidae (21.4%) como las familias más abundantes, batiendo directamente las plantas en algodón ecológico.
Dos características importantes habrían potenciado el predominio de Thomisidae en este estudio: la preferencia de alimentación de esta familia sobre algunas plagas del cultivo como Chloridea virescens y Anomis texana (Aguilar 1977, 1988), las cuales fueron observadas en el cultivo evaluado, representando un componente importante de los enemigos naturales de las plagas en este agroecosistema (Almada et al. 2012); y a su capacidad de vivir y cazar al acecho en las flores, siendo el cultivo de algodón propicio para albergar este tipo de arañas (Pérez-Guerrero et al. 2009).
Al igual que en el presente estudio, Misumenops se encontró en cultivos de algodón en Arizona (Estados Unidos) (Plagens 1983), Sao Paulo (Brasil) (Gravena & Da Cunha 1991) y Sudáfrica (Dippenaar-Schoeman et al. 1999), y fue el más representativo (79.09%) en campos de algodón transgénico y convencional en Santa Fe (Argentina) (Almada et al. 2012). Este género también fue registrado por Aguilar (1977, 1988) a lo largo de la costa peruana, Aguilar y Lamas (1980) y Sarmiento (1992), con varias especies (M. amabilis, M. variegatus y Misumenops sp.) en el algodonero peruano, siendo uno de los más abundantes y depredadores de las principales plagas de lepidópteros en el cultivo (Aguilar 1988).
La mayor abundancia de este género podría deberse al bosque cercano al cultivo, el cual podría constituir un refugio para estas arañas, como ocurrió con arbustos de Larrea y Artemisia aledaños a campos de algodón de los valles de Mammoth y Avra (Arizona, Estados Unidos) (Plagens 1983). Estos hábitats adyacentes representarían la ‘guardería’ para Misumenops, que eventualmente colonizaría estos cultivos (Plagens 1983). El uso de un hábitat diferente en su ciclo de vida podría ser una estrategia de Misumenops, ya que requiere de 145 a 531 días a 25°C para alcanzar la madurez, lo cual es mucho más prolongado que el periodo del cultivo de algodón, ocasionando que no complete todo su ciclo de vida en este agroecosistema y necesite de un hábitat cercano estable (Plagens 1983).
A pesar de lo registrado en esta investigación, la temperatura y humedad son considerados influyentes en la riqueza y abundancia de las arañas, así también otros factores abióticos (altitud, estacionalidad y estabilidad ambiental) (Ferenc et al. 1999, Liljesthröm et al. 2002, Jiménez-Valverde & Lobo 2007, Rodrigues et al. 2009). No obstante, existen pocos trabajos sobre la relación de la temperatura y humedad con la araneofauna, de los cuales han mostrado resultados contrastantes: Le Sar y Unzicker (1978) registraron una mayor diversidad de arañas en el primer año de estudio, mucho más húmedo que el segundo, en cultivos de soja en Illinois (Estados Unidos); Rodrigues et al. (2009) solo hallaron relevancia entre la correlación de la temperatura y abundancia de arañas en el borde del bosque (adyacente a un cultivo de arroz), ni el cultivo ni un pastizal adyacente mostraron significancia; y Vijaykumar (2004) no encontró correlación significativa de la temperatura y humedad con las arañas en un cultivo de arroz en India.
La riqueza encontrada en el presente trabajo (50 especies y 18 familias) es similar a las 50 especies y 17 familias descritas por Aguilar (1977, 1988) en diversos valles algodoneros a lo largo de la costa peruana, y superior a las 27 especies y 19 familias citadas por Sarmiento (1992) en el algodón en Perú. Se reconocieron 37 especies y 6 familias nuevas registradas, llegando a un total de 87 especies y 24 familias de arañas para el cultivo de algodón en Perú. Este número considerable de nuevos registros en una sola campaña de evaluación podría deberse al esfuerzo de muestreo y técnicas de evaluación, ya que se evaluó cada 7 días (32 evaluaciones), desde el inicio del cultivo (emergencia de plántulas) hasta su término (cosecha) y las técnicas de evaluación fueron más completas en comparación a estudios previos, como los de Aguilar (1974, 1975, 1976, 1978a, 1978b, 1979a, 1979b), donde realizó 4 a 5 evaluaciones por campaña, y en su gran mayoría con una sola técnica de evaluación (10 plantas por campo).
La abundancia y riqueza de arañas registradas en esta investigación es alta, a pesar de tratarse de un agroecosistema, un hábitat interrumpido y fuertemente influenciado por las prácticas culturales, las cuales son consideradas uno de los factores más dañinos para la araneofauna (Ferenc et al. 1999, Marc et al. 1999, Butt & Muhammad 2012). Esto podría deberse al sistema de manejo agronómico del cultivo de algodón en Perú, ya que se realiza un manejo integrado de plagas, sin la necesidad de la aplicación tan seguida de insecticidas como en otros cultivos, con el fin de respetar la acción de la fauna benéfica en este agroecosistema (Herrera 2010). Por lo que es importante conocer el sistema de manejo agronómico en un cultivo, ya que la abundancia y riqueza de la araneofauna estaría relacionada con ello, siendo pobre en un cultivo intensivo o convencional, y con mayor riqueza (incluso mayor que un hábitat natural) en un cultivo orgánico o de bajos insumos, debido principalmente a la ausencia o disminución de agroquímicos (Ferenc et al. 1999, Butt & Muhammad 2012).
En este estudio Linyphiidae y Salticidae fueron las familias con mayor riqueza de especies (16% y 14% respectivamente); sin embargo, Aguilar (1977) y Sarmiento (1992) registraron a Salticidae como la familia con mayor número de especies con el 22% y 26% de especies cada uno. La mayor riqueza de especies de Linyphiidae podría explicarse a que esta familia teje telas pequeñas, debajo de las hojas, en diversos órganos de la planta, en pequeñas grietas o en la hojarasca del suelo (Aguilar 1988), siendo menor la probabilidad del daño de sus telas por el manejo agronómico; otro factor importante podría ser su alta capacidad de dispersión mediante ballooning, bien adaptada al patrón espacial y temporal de las alteraciones de los agroecosistemas (Halley et al. 1996, Marc et al. 1999). Además, la alta riqueza de especies de Salticidae podría deberse a su preferencia de alimentación sobre algunas plagas del cultivo de algodón (Aguilar 1988).
El inventario de arañas resultó confiable al obtener valores mayores al 70% de especies registradas (Jiménez-Valverde & Hortal 2003) en comparación con los estimadores Chao 1 y Jack 1. Esto podría deberse al continuo esfuerzo de muestreo realizado (cada siete días), ya que los valores de los estimadores de riqueza de especies dependen del esfuerzo de muestreo, donde diferentes esfuerzos producen diferentes estimaciones (Samu & Lövei 1995). En contraste, cuando la evaluación se realiza en periodos separados el esfuerzo podría diluirse y volverse una evaluación más superficial (Deza 2009). Así como en este trabajo, generalmente en estudios sobre la calidad de los inventarios de arañas las curvas de acumulación se encuentran lejos de la asíntota, sin lograr registrar todas las especies debido al elevado número de especies raras que suelen aparecer (Jiménez-Valverde & Hortal 2003). Para optimizar la recolecta de estas especies en este estudio podría ser necesario aumentar las unidades de muestreo (un mayor número de trampas de caída instaladas o de plantas evaluadas en la búsqueda directa) o la utilización de más técnicas de evaluación, por lo que el inventario de la araneofauna no es una tarea sencilla (Jiménez-Valverde & Hortal 2003).
Al igual que en el presente estudio, en cultivos de algodón transgénico y convencional en Argentina, el gremio de las emboscadoras fue el más abundante (83.91%), por la preferencia de alimentación de ésta sobre larvas de plagas importantes de lepidópteros en el cultivo (Almada et al. 2012). Los resultados de esta investigación fueron similares a lo registrado por Aguilar (1977, 1998), Uetz et al.(1999) (Arkansas-Estados Unidos) y Dippenaar-Schoeman et al. (1999) (Sudáfrica); donde clasificaron la araneofauna del algodón en dos gremios: cazadoras y tejedoras, registrando a las cazadoras mayor o igual al 60%, y englobando los ocho gremios encontrados en esta investigación las cazadoras y tejedoras constituyeron el 81.73% y las tejedoras el 18.27%. Esto podría deberse a que las arañas cazadoras no eran afectadas notablemente por las prácticas agronómicas; en cambio las arañas tejedoras sí, ya que al tejer sus telas éstas eran destruidas directamente por la manipulación de las plantas durante el manejo agronómico, y al no poder alimentarse disminuían considerablemente (Aguilar 1977, 1998).
Las características significativas para la composición de los gremios de arañas en un cultivo son la complejidad estructural del cultivo hospedero, microambientes, nivel de disturbio, las áreas adyacentes, etc. (Uetz et al. 1999). Este último factor podría haber influenciado el patrón de estructuración de los gremios en este agroecosistema, ya que actuaría como fuerza selectiva de lo que está disponible, colonizando así cultivos cercanos (Uetz et al. 1999). Como lo ocurrido en campos de alfalfa en Virginia (Howell & Pienkowski 1971) y California (Yeargan & Dondale 1974), que siendo evaluados con las mismas técnicas (redada y aspiración con D-vac), se observó una diferente predominancia de gremios en cada estudio. Howell y Pienkowski (1971), encontraron como los más dominantes a tejedoras orbiculares (42.7%) y errantes tejedoras (37.6%); en cambio, Yeargan y Dondale (1974), registraron como los más abundantes a corredoras terrestres (60%) y errantes tejedoras (33.6%).
Aunque no se midió la altura de las plantas de algodón, conforme fue avanzando el desarrollo del cultivo las arañas fueron incrementando, esto podría deberse a la relación de la araneofauna con el desarrollo o vigor de las plantas (Aguilar 1974, 1976, 1977; DippenaarSchoeman et al. 1999; Almada et al. 2012). Esto se explicaría gracias a la mayor estructuración física del hábitat, generando más microhábitats favorables, los cuales son factores importantes para la araneofauna (Rodrigues et al. 2009; Jiménez-Valverde & Lobo 2007), que potenciado a la característica propia de la planta de algodón, por ser una planta arbustiva dicotiledónea, gozaría de una buena complejidad estructural que beneficiaría aún más el desarrollo de su araneofauna (Young & Edwards 1990).
Desde la fase temprana del cultivo se registró la presencia de arañas, este resultado concuerda con Dippenaar-Schoeman et al. (1999), donde hallaron que las arañas fueron unos de los primeros artrópodos depredadores en colonizar las plantas emergentes de algodón. Esta población inicial en la investigación podría deberse a su migración desde otras zonas cercanas, ya que las arañas son conocidas por sus excelentes habilidades de dispersión y colonización (Marc et al. 1999). Estas áreas podrían tratarse de cultivos vecinos o zonas con vegetación diferenciable y generalmente menos manejadas, como el área de malezas o el bosque de pinos en este estudio, este último por tratarse de un hábitat más complejo, extenso y casi sin perturbación podría servir como refugio y sitio de hibernación para las arañas, tal como citaron Öberg & Ekbom (2006), Öberg et al. (2007) y Rodrigues et al. (2009) para áreas cercanas a los márgenes del cultivo, con vegetación diferenciable, compleja y generalmente menos manejada.
La disminución de arañas después de la aplicación del insecticida clorpirifos concuerda con la literatura encontrada, ya que se trata de un organo fosforado de amplio espectro, neurotóxico, que actúa por contacto e ingestión, muy perjudicial para las arañas (Amalin et al. 2000, Pekár 2002, Fountain et al. 2007, Pekár 2012). Sin embargo, las arañas no desaparecieron en noviembre, tal como registró Aguilar (1974, 1976, 1977), ya que a pesar de su detrimento luego de la aplicación de insecticidas arsenicales y orgánicos, su población se reestableció. Esto podría relacionarse al tiempo de vida media que posee este insecticida en la superficie del suelo, de 3 a 14 días aproximadamente (Fountain et al. 2007). A diferencia de la aplicación inicial del insecticida en esta investigación, no se recomiendan los tratamientos tempranos de plaguicidas en los campos de algodón y se consideran innecesarios, porque las arañas retardan el brote precoz de plagas en la temporada de crecimiento del cultivo (Mansour 1987). Diversos estudios también corroboran el detrimento en la riqueza y abundancia de arañas en campos aplicados con insecticidas, en comparación con las máximas poblaciones de arañas obtenidas en campos no aplicados, evaluados por Mansour (1987) en Israel, Van den Berg et al. (1990) en Sudáfrica, Rajput et al. (2012) en Pakistán, y Solanki y Kumar (2014) en la India.
No se encontraron trabajos con qué comparar los resultados de esta investigación sobre la relación del desahije y fertilización con la araneofauna, aunque Butt y Muhammad (2012) y Andrey et al. (2014) citaron que campos altamente fertilizados albergaron más detritívoros y herbívoros, y por ello más depredadores, como las arañas.
Al igual que en el presente estudio, en campos de algodón en Argentina (Almada et al. 2012), Pakistán (Khuhro et al. 2012, Rajput et al. 2012) e India (Solanki y Kumar 2014), se encontraron las máximas poblaciones de arañas y con picos de crecimiento durante la floración y fructificación del cultivo, cuando la planta de algodón se mostraba exuberante en follaje. Este mayor desarrollo de la araneofauna podría deberse al aumento de sus presas de insectos que albergaría el cultivo en relación con el crecimiento de la planta de algodón (Aguilar 1977, Van den Berg 1989, Van den Berg & Dippenaar-Schoeman 1991, Dippenaar-Schoeman et al. 1999, Almada et al. 2012, Mahalakshmi & Jeyaparvathi 2014).
Los picos de máximo crecimiento observados en esta investigación podrían estar relacionados a diversos factores, uno de ellos podría ser la migración de las arañas de las cosechas de campos cercanos buscando un nuevo nicho, como lo sucedido en campos de algodón en India (Mahalakshmi & Jeyaparvathi 2014), otro podría ser las condiciones climáticas o variaciones estacionales, tal como citaron Van den Berg et al. (1990), Almada et al. (2012), y Mahalakshmi y Jeyaparvathi (2014) en campos de algodón en Sudáfrica, Argentina e India respectivamente. Por último, Almada et al. (2012) mencionaron que también podría deberse a la aplicación del regulador de crecimiento en las plantas, práctica que se realiza generalmente en esas zonas algodoneras antes de la formación de bellotas.
La disminución de arañas después de la aplicación del insecticida lufenuron, al estar precedida de la fuerte disminución de las mismas luego del primer riego, podría inferirse a que hubieron otros factores que influyeron en la disminución de las arañas; además que estudios previos indicaron que este insecticida no tuvo mucho impacto en la fauna benéfica del algodón, siendo seguro para los adultos de varias familias de arañas (Javaid et al. 1999, Sechser et al. 2001, Mansfield et al. 2006), y que en comparación con clorpirifos, mostró tener menor efecto en los depredadores (Desuky et al. 2012). Esto se debería a que clorpirifos por ser un neurotóxico tiene un amplio espectro de actividad, en cambio, lufenuron posee un buen grado de selectividad, ya que inhibe la síntesis de quitina (un constituyente importante en el exoesqueleto de artrópodos) en larvas de Lepidoptera durante el proceso de muda, su acción es por ingestión y con poca actividad por contacto (Javaid et al. 1999, Sechser et al. 2001, Desuky et al. 2012).
No se pudo reconocer en este estudio el efecto del riego sobre la población de la araneofauna, ya que hubieron disminuciones y aumentos de su población luego de los mismos, más aún existen pocos estudios sobre el tema, con resultados poco específicos y contrastantes sobre su relación con las arañas. Según Butt y Muhammad (2012) el riego afecta la distribución y la abundancia de las arañas en los cultivos. Srikanth et al. (1997) encontraron que el riego resultó negativo para la población de arañas en un cultivo de caña de azúcar (siendo más dañino el riego por gravedad que por goteo superficial). En cambio, Andrey et al. (2014) registraron que el riego tuvo un efecto positivo en la riqueza de artrópodos en un pastizal, que al aumentar los herbívoros incrementaron los depredadores. Latha (2004) comparó tres niveles de irrigación (50%, 75% y 100%) en un cultivo de algodón en Texas (Estados Unidos), y aunque el número de arañas no varió grandemente entre los tratamientos, halló que los promedios estacionales de arañas fueron mayores en el tratamiento al 100%, afirmando que condiciones más húmedas favorecen poblaciones más altas de artrópodos depredadores, aunque no es el caso para todas las especies.
La disminución progresiva de arañas en los meses siguientes después del último riego podría deberse a que las plantas comenzaron a ponerse cada vez más secas, reduciendo su follaje, como se registró en campos de soya, donde la senescencia de las plantas y la descomposición del material vegetal muerto redujeron los sitios potenciales de hibernación de las arañas, generando que se muevan fuera de los cultivos en búsqueda de hábitats adecuados para su hibernación o estancia (Turnbull 1960, Duffey 1962, Culin & Rust 1980, Ferguson et al. 1984, Liljesthrom et al. 2002). Las cosechas realizadas podrían haber intensificado la disminución de arañas en el mes de julio, debido a la interrupción y destrucción de este hábitat por parte del personal para poder retirar las motas de algodón de las plantas, como en estudios previos, donde la cosecha afectó negativamente la densidad de arañas en los campos agrícolas, obligándolas a su emigración (Marc et al. 1999, Liljesthrom et al. 2002, Butt &Muhammad 2012). Aguilar (1977) señaló que el manejo agronómico del cultivo de algodón peruano influyó negativamente a diversas familias de arañas, estando a su vez este efecto relacionado a la estrategia de caza o gremios de arañas.
La mayor abundancia de las familias Lycosidae y Gnaphosidae en la primera etapa del cultivo, en comparación de las demás familias (sobretodo Thomisidae y Anyphaenidae que fueron las más abundantes) en la segunda etapa, podría deberse a sus hábitos terrestres, ya que sus estrategias de caza podrían permitirles establecerse en el cultivo sin necesitar del mayor crecimiento de las plantas, por no depender de la estructura física de las mismas. A pesar de que después las poblaciones de estas familias disminuyeron, la distribución de tresde sus especies estuvieron presentes durante casi todo el periodo del cultivo, demostrándose así su importancia como colonizadoras y su mayor amortiguación a los cambios en el cultivo.
Análisis por sectores
La relación de la mayor abundancia de arañas entre los sectores 1 y 2 mostrado en el dendrograma podría deberse a que en estos sectores las plantas se encontraban más desarrolladas, en comparación sobretodo de los
sectores 4 y 5, donde eran más pequeñas, aunque no se midió directamente la altura de las plantas.
La mayor riqueza y diversidad de especies y gremios registrados en los sectores 3, 4 y 5, y el comportamiento asociado de mayor diversidad de especies y gremios entre los sectores 4 y 5 observados en los dendrogramas podrían estar relacionados a la mayor contigüidad de éstos a la zona de malezas y el bosque de pino, ya que las áreas cercanas a los márgenes del cultivo, con vegetación diferenciable, siendo a veces más complejas y generalmente menos manejadas, podrían servir como refugios y sitios de hibernación para las arañas (Schmidt & Tscharntke 2005, Öberg et al. 2007, Rodrigues et al. 2009). Tal como se encontró en estudios previos, como Rodrigues et al. (2009), donde registraron una mayor diversidad de arañas en el borde del bosque adyacente de un cultivo de arroz que en el mismo cultivo, y Gardiner et al. (2010), donde encontraron que los hábitats boscosos fueron importantes en la composición de araneofauna en cultivos de colza oleaginosa. Mientras que la mayor dominancia encontrada en el sector 2 podría deberse a la mayor abundancia registrada del gremio de las emboscadoras en este sector (más de la mitad de su población) en comparación con las demás.
El primer pico de máximo crecimiento de la población de arañas en enero fue debido principalmente a la abundancia de arañas en el sector 3 y por Lycosidae; a diferencia del segundo pico de máximo crecimiento de toda la población de arañas en marzo, debido a la abundancia de arañas en todos los sectores y principalmente por Thomisidae. Los diferentes niveles de abundancia de las arañas en todos los sectores a través del periodo evaluado podría deberse a los desplazamientos de las mismas de acuerdo a los requisitos necesarios para su establecimiento; ya que las arañas permanecerán en un hábitat si les satisfacen todos estos requisitos (estructura física del microhábitat, condiciones microclimáticas, competencia con otros depredadores y disponibilidad de presas); y si uno de ellos es modificado, la araña se moverá a otro lugar, mientras que otros cambios estarían relacionados a su ciclo biológico (Marc et al. 1999, Arango et al. 2000, Sunderland & Samu 2000, Quevedo & Vasconcellos-Neto 2003).
Análisis por técnicas de evaluación
La técnica de búsqueda directa registró la mayor asociación de especies, familias y gremios con preferencia de localización en el follaje por sus estrategias de caza (Ferguson et al. 1984, Ubick et al. 2005, Jocqué & Dippenaar-Schoemann 2006); mientras que las trampas de caída registraron un mayor número de especies, familias y gremios habitantes de los estratos inferiores de la vegetación y en general del suelo (Dean et al. 1982; Liljesthrom et al. 2002; Ubick et al. 2005; Jocqué & Dippenaar-Schoemann 2006), descrito como el método más eficiente para capturar arañas epigeas (Dean et al. 1982, Work et al. 2002, Álvares et al. 2004). Los resultados de cada técnica de recolecta en esta investigación corroboran la segregación que tiene la araneofauna (comunidades de arañas que habitan el suelo y el follaje) en este agroecosistema (Culin & Rust 1980)
La utilización de estas dos técnicas podría haber favorecido la alta riqueza de arañas en este estudio, ya que son reconocidas y recomendadas para la evaluación de arañas en variados hábitats (Churchill & Arthur 1999, Work et al. 2002, Ludy & Lang 2004, Álvarez et al. 2004, Wade et al. 2006, Jud & Schmidt-Entling 2008). Al mismo tiempo, la combinación de ambas técnicas fue acertada debido al bajo número de especies comunes registradas entre ambas, puesto que si dos métodos tienen un alto grado de solapamiento taxonómico se estaría desperdiciando el esfuerzo de muestreo en vez de poner más empeño en la evaluación de otros estratos (Dobyns 1997). Además, con una sola técnica evaluada hubiera habido un gran sesgo en la medición de la diversidad de la araneofauna en el cultivo, como detalló Marc et al. (1999), donde la evaluación de cada técnica empleada en una sola área de estudio enfatizó grandes diferencias en las composiciones de su araneofauna, debido a que comprende varios grupos ecológicamente distintos que habitan diferentes estratos de microhábitats, por lo que se recomienda la combinación de varias técnicas de muestreo para una evaluación apropiada de la araneofauna (Green 1999, Marc et al. 1999, Jiménez-Valverde & Hortal 2003, Álvares et al. 2004, Brennan et al. 2005).
La menor abundancia de arañas registradas en las trampas de caída podría deberse a que esta técnica evalúa especialmente la tasa de actividad de las arañas, y no refleja una real abundancia, por ello capturan en gran medida arañas adultas por su mayor tasa de movimiento, en comparación a los juveniles, a pesar que éstos ocurren en grandes números en el suelo y cerca de él (Topping & Sunderland 1992, Topping & Luff 1995, Azevedo et al. 2002, Work et al. 2002, Álvares et al. 2004). Esto también explicaría la mayor riqueza de arañas registradas con esta técnica, ya que al ser más eficientes en capturar arañas adultas, es precisamente en este estado de desarrollo donde se pueden identificar las arañas al nivel de especie o morfoespecie. Estos resultados concuerdan con lo descrito por Canard (1982), Merrett y Snazell (1983), y Churchill y Arhtur (1999) en la evaluación de las arañas en brezales, donde las trampas de caída resultaron en más taxones de arañas que otros métodos cuyo objetivo fue el estrato de follaje. A diferencia de Álvarez et al. (2004), donde encontraron que usando la técnica de golpeo proporcionó una mayor riqueza y abundancia de arañas que las trampas de caída en una reserva de bosque urbano.
En conclusión, sí se registró una alta riqueza y abundancia de arañas en este agroecosistema, donde su población fluctuó durante todo el periodo del cultivo, encontrándose una mayor abundancia durante su etapa media, infiriendo este resultado a que las plantas en ese momento se presentaron más grandes y más exuberantes en follaje. A pesar de aquellas disminuciones luego de la aplicación de los insecticidas y de las demás prácticas agronómicas, las arañas siempre estuvieron presentes durante todo el cultivo, y su abundancia en cada mes no se vio afectada, excepto en el último mes, ya que se registró una fuerte disminución de arañas, por lo que se concluye que la cosecha sí influyó negativamente en su población, pero las demás prácticas agronómicas y la temperatura y humedad no se encontraron influyentes en la araneofauna de este estudio.
Agradecimientos:
Nuestro agradecimiento especial a Manuel Andía y Bryan Portuguez por su ayuda en la identificación de algunas especies de arañas; a Javier Fernández y Pamela Cardich por su colaboración con el trabajo de campo; a Nicolás Tarmeño y Alfredo Giraldo por su ayuda en los análisis estadísticos; y a Marco Antonio Desales-Lara por la revisión y sugerencias para mejorar el manuscrito.
Información sobre los autores:
LC, DS, CV: realizaron el diseño experimental; LC: los experimentos; LC, DS, CV: analizaron los datos, redactaron, revisaron y aprobaron el manuscrito.
Conflicto de intereses:
Los autores no incurren en conflictos de intereses.
Fuentes de financiamiento:
El presente trabajo se realizó con fondos propios.
Aspectos éticos / legales:
Este trabajo no incurrio en ningun problema ético ni legal.
Literatura citada
Aguilar P. 1965. Notas sobre las arañas en el campo cultivado. Revista peruana de Entomología. 8(1): 80-83. [ Links ]
Aguilar P. 1968. Araneida frecuentes en el algodonero. Revista peruana de Entomología. 11(1): 92-95. [ Links ]
Aguilar P. 1974. Arañas del campo cultivado. I: Población de araneidos en algodonales de Cañete, Huaura y Rímac. Revista peruana de Entomología. 17(1): 21-27. [ Links ]
Aguilar P. 1975. Arañas del campo cultivado. II: Fluctuación de las familias de araneidos en algodonales de la costa central. Revista peruana de Entomología. 18(1): 25-28. [ Links ]
Aguilar P. 1976. Arañas del campo cultivado. III: Araneidos en algodonales del valle de Lurín. Revista peruana de Entomología. 19(1): 71-72. [ Links ]
Aguilar P. 1977. Las arañas en el agroecosistema algodonero de la costa peruana. Anales científicos Universidad Nacional Agraria La Molina. 15(1-4): 108-121. [ Links ]
Aguilar P. 1978. Arañas del campo cultivado. IV: Población de araneidos del algodonero en Chillón y Chancay – Hua ral. Revista peruana de Entomología. 21(1): 39-41. [ Links ]
Aguilar P. 1979a. Arañas del campo cultivado. VI: Observaciones en algodonales de la costa norte del Perú. Revista peruana de Entomología. 22(1): 71-73. [ Links ]
Aguilar P. 1979b. Artrópodos epígeos del campo cultivado. I: Estudio preliminar en el algodonero de Cañete. Revista peruana de Entomología. 22(1): 87-90. [ Links ]
Aguilar P. 1988. Las arañas como controladores de plagas insectiles en la agricultura peruana. Revista peruana de Entomología. 31: 1-8. [ Links ]
Aguilar P. & G. Guérovich. 1978. Arañas del campo cultivado. V:Frecuencia de araneidos en algodonales de Mala, Asia yChilca. Revista peruana de Entomología. 21(1): 42-45. [ Links ]
Aguilar P. & J. Lamas. 1980. El cultivo del algodonero. Revista peruana de Entomología. 23(1): 91-97. [ Links ]
Almada M., M. Sosa & S. González. 2012. Araneofauna (Arachnida: Araneae) en cultivos de algodón (Gossypium hirsutum) transgénicos y convencionales en el norte deSanta Fe, Argentina. Revista de Biología Tropical. 60(2):611-623. https://doi.org/10.15517/rbt.v60i2.3946 [ Links ]
Álvares E., E. Machado, C. Azevedo & M. De-Maria. 2004. Composition of the spider assemblage in an urban forest reserve in southeastern Brazil and evaluation of a two sampling method protocols of species richness estimates. Revista Ibérica de Aracnología. 10: 185-194. [ Links ]
Amalin D., J. Pena, S. Yu & R. McSorley. 2000. Selective toxicity of some pesticides to Hibana velox (Araneae: Anyphaenidae), a predator of citrus leafminer. Florida Entomologist. 83: 254-262. https://doi.org/10.2307/3496343 [ Links ]
Andrey A., J. Humbert, C. Pernollet & R. Arlettaz. 2014. Experimental evidence for the immediate impact of fertilization and irrigation upon the plant and invertebrate communities of mountain grasslands. Ecology and Evolution. 4(12): 2610-2623. https://doi.org/10.1002/ece3.1118 [ Links ]
Arango A., V. Rico-Gray & V. Parra-Tabla. 2000. Population structure, seasonality, and habitat use by the green lynx spider Peucetia viridans (Oxiopidae) inhabiting Cnidoscolus aconitifolius (Euphorbiaceae). Journal of Arachnology. 28:185-194. https://doi.org/10.1636/0161-8202(2000)028[0185:PSSAHU]2.0.CO;2 [ Links ]
Avalos G., M. Bar, E. Oscherov & A. González. 2013. Diversidad de Araneae en cultivos de Citrus sinensis (Rutaceae) de la Provincia de Corrientes, Argentina. Revista de Biología Tropical. 61(3): 1243-1260. https://doi.org/10.15517/rbt.v61i3.11938 [ Links ]
Azevedo C., E. Machado, E. Álvares & M. Demaria. 2002. Comparison of spider soil communities in six differing habitats in the ecological station of Universida de Federal de Minas Gerais, Brazil. Bios. 10: 47-53. [ Links ]
Beingolea O. 1959. Notas sobre la bionómica de ara-as e insectos benéficos que ocurren en el cultivo de Algodón. Revista peruana de Entomología. 2(1): 36-44. [ Links ]
Brennan K., J. Majer & M. Moir. 2005. Refining sampling protocols for inventorying invertebrate biodiversity: influence of drift-fence length and pitfall trap diameter on spiders. Journal of Arachnology. 33(3): 681-702. https://doi.org/10.1636/M01-105.1 [ Links ]
Bristowe W. 1971. The world of spider. Collins New naturalist, London. 304 [ Links ]
Butt A. & S. Muhammad. 2012. Effect of different agricultural practices on spiders and their prey populations in small wheat fields. Acta Agriculturae Scandinavica Section B - Soil and Plant Science. 62: 374-382. https://doi.org/10.1080/09064710.2011.624544 [ Links ]
Canard A. 1982. Utilisation comparee de quelques methodes d'echantillonnage pour l'etude de la distribution des araignees en landes. Atti della Società Toscana di Scienze Naturali. 88: 84–94. [ Links ]
CCI (Centro de Comercio Internacional). 2007. (en línea). Guía del exportador de algodón.<http://www.intracen.org/guia-del-algodon-home/>. Acceso 26/05/2013. [ Links ]
Churchill T. 1993. Effects of sampling method on composition of a Tasmanian coastal heathland spider assemblage. Memoirs of the Queensland Museum. 33 (2): 475-481. [ Links ]
Churchill T. & M. Arthur. 1999. Measuring spider richness: effects of different sampling methods and spatial and temporal scales. Journal of Insect Conservation. 3: 287-295. https://doi.org/10.1023/A:1009638706789 [ Links ]
Culin J. & R. Rust. 1980. Comparison of the ground surface and foliage dwelling spider communities in a soybean habitat. Environmental Entomology. 9(5): 577-582. https://doi.org/10.1093/ee/9.5.577 [ Links ]
Dean D. & W. Sterling. 1987. Distribution and abundance patterns of spiders inhabiting cotton in Texas. Texas Agricultural Experimental Station Bulletin N° B-1566. College Station, TX, 1-8 pp. [ Links ]
Dean D., W. Sterling & N. Horner. 1982. Spiders in eastern Texas cotton fields. Journal of Arachnology. 10: 251-260. [ Links ]
Debach P. & D. Rosen. 1991. Biological control by natural enemies. Cambridge University Press. Cambridge. 2nd ed. 440 pp. [ Links ]
Desuky W., E. El-Khayat, M. Azab & M. Khedr. 2012. The influence of some insect growth regulators and bio-insecticides against cotton leafworm and some associated predators under field conditions. Egyptian Journal of Agricultural Research. 90(1): 31-53. [ Links ]
Deza M. 2009. Variación de la araneofauna en las épocas seca y lluviosa en tres tipos de hábitat en la cuenca del río Los Amigos - Madre de Dios. Tesis para optar el título de Biólogo. Universidad Nacional Agraria La Molina. Lima-Perú. 110 pp. [ Links ]
Dinkins R., J. Brazzel & C. Wilson. 1970. Seasonal incidence of major predaceous arthropods in Mississippi cotton fields. Journal of Economic Entomology 63: 814-817. https://doi.org/10.1093/jee/63.3.814 [ Links ]
Dippenaar-Schoeman A., A.M. Van Den Berg & A. Van Den Berg. 1999. Spiders in south african cotton fields: species diversity and abundance (Arachnida: Araneae). African Plant Protection. 5(2): 93-103. [ Links ]
Dobyns J. 1997. Effects of sampling intensity on the collection of spider (Araneae) species and the estimation of species richness. Pest Management and Sampling. 26(2): 150-162. https://doi.org/10.1093/ee/26.2.150 [ Links ]
Dodson G., A. Anderson. & L. Stellwag. 2015. Movement, sex ratio and population density in a dwarf male spider species, Misumenoides formosipes (Araneae: Thomisidae). Journal of Arachnology. 43: 388-393. https://doi.org/10.1636/arac-43-03-388-393 [ Links ]
Duffey E. 1962. A population study of spiders in Limestone Grassland. Journal of Animal Ecology. 31: 571-599. https://doi.org/10.2307/2054 [ Links ]
Ferenc S., K. Sunderland & C. Szinetár. 1999. Scale-dependent dispersal and distributtion patterns of spiders in agricultural systems: a review. Journal of Arachnolgy. 27: 325-332. [ Links ]
Ferguson H., R. McPherson & W. Allen. 1984. Ground- and foliage-dwelling spiders in four soybean cropping systems. Environmental Entomology. 13(4): 975-980. https://doi.org/10.1093/ee/13.4.975 [ Links ]
Fountain M., V. Brown, A. Gange, W. Symondson & P. Murray. 2007. The effects of the insecticide chlorpyrifos on spider and Collembola communities. Pedobiologia. 51: 147-158. https://doi.org/10.1016/j.pedobi.2007.03.001 [ Links ]
Gardiner M., D. Landis, C. Gratton, N. Schmidt, M. O' Neal, E. Mueller, J. Chacon & G. Heimpel. 2010. Landscape composition influences the activity density of Carabidae and Arachnida in soybean fields. Biological Control. 55: 11-19. https://doi.org/10.1016/j.biocontrol.2010.06.008 [ Links ]
Ghavami S., M. Taghizadeh, G. Amin & Z. Karimian. 2007. Spider (Order Araneae) fauna of cotton fields in Iran. Journal of Applied Biological Sciences. 1(2): 07-11. [ Links ]
Gravena S. & H. Dacunha. 1991. Predation of cotton leafworm first instar larvae, Alabama argillacea (Lep: Noctuidae). Entomophaga. 36(4): 481-491. http://dx.doi.org/10.1007/BF02374430 [ Links ]
Green J. 1999. Sampling method and time determines composition of spider collections. Journal of Arachnology. 27:176-182. Gunnarsson B. & A. Andersson. 1996. Sex ratio variation in sheet-web spiders: options for female control? Proceedings of the Royal Society of London B. 263: 1177-1182. https://doi.org/10.1098/rspb.1996.0172 [ Links ]
Halley J., C. Thomas & P. Jepson. 1996. A model for the spatial dynamics of Linyphiid spiders in farmland. Journal of Applied Ecology. 33: 471-492. https://doi.org/10.2307/2404978 [ Links ]
Herrera, J. 1987. Importancia del control biológico en el cultivo del algodonero. Revista Peruana de Entomología. 30:25-28. [ Links ]
Herrera, J. 2010. Primera experiencia a nivel mundial del Manejo Integrado de Plagas: el caso del algodonero en el Perú. Revista Peruana de Entomología. 46(1): 1-8. [ Links ]
Howell J. & Pienkowski, R. 1971. Spider populations in alfalfa, with notes on spider prey and effect of harvest. Journal of Economic Entomology. 64:163-168. https://doi.org/10.1093/jee/64.1.163 [ Links ]
Javaid I., R. Uaine & J. Massua. 1999. The use of insect growth regulators for the control of insect pests of cotton. International Journal of Pest Management 45(4):245-247. http://dx.doi.org/10.1080/096708799227635 [ Links ]
Jiménez-Valverde A. & J. Hortal. 2003. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología 8:151-161. [ Links ]
Jiménez-Valverde A. & J. Lobo. 2007. Determinants of local spider (Araneidae and Thomisidae) species richness on a regional scale: climate and altitude vs. Habitat structure. Ecological Entomology. 32:113-122. https://doi.org/10.1111/j.1365-2311.2006.00848.x [ Links ]
Jocqué R. & A. Dippenaar-Schoeman. 2006. Spider families of the world. Royal Museum for Central Africa. 336 p. [ Links ]
Jud P. & M. Schmidt-Entling. 2008. Fluid type, dilution, and bitter agent influence spider preservation in pitfall traps.Entomologia Experimentalis et Applicata 129: 356-359.https://doi.org/10.1111/j.1570-7458.2008.00773.x [ Links ]
Khuhro R., A. Ghafoor, A. Mahmood, M. Khan, S. Andleeb, M. Bukhari, I. Maqsood, M. Shahjahan & N. Baloch. 2012. Impact of temperature and relative humidity of population abundance of predatory spiders in cotton fields. Journal of Animal and Plant Sciences. 22(3): 649-652. [ Links ]
Latha P. 2004. Effect of irrigation regimes on crop phenology andarthropod populations in Texas high plains cotton. Tesis para optar el Grado de Maestría en Ciencias. Universidad de Texas Tech. Texas-Estados Unidos. 96 pp. [ Links ]
Le Sar C. & J. Unzicker. 1978. Soybean spiders: species composition, population densities, and vertical distribution. Illinois Natural History Surveys - Biol. Notes 107: 3-14. [ Links ]
Liljesthröm G., E. Minervino, D. Castro & A. Gonzalez. 2002. La comunidad de ara-as para el cultivo de soja en la Provincia de Buenos Aires, Argentina. Neotropical Entomology. 31(2): 197-210. http://dx.doi.org/10.1590/S1519-566X2002000200005 [ Links ]
Ludy C. & A. Lang. 2004. How to catch foliage-dwelling spiders (Araneae) in maize fields and their margins: a comparison of two sampling methods. Journal of Entomology and Nematology 128(7):501-509. https://doi.org/10.1111/j.1439-0418.2004.00880.x [ Links ]
Mahalakshmi R. & S. Jeyaparvathi. 2014. Diversity of spider fauna in the cotton field of Thailakulam, Virudhunagar district, Tamil Nadu, India. The Journal of Zoology Studies 1(1):12-18. [ Links ]
Maloney D., F. Drummord & R. Alford. 2003. Spider predation in agroecosystems; can spiders effectively control pest populations? The University of Maine. Technical Bulletin 190, 32 p. [ Links ]
Mansfield S., M. Dillon. & M. Whitehouse. 2006. Are arthropod communities in cotton really disrupted? An assessment of insecticide regimes and evaluation of the beneficial disruption index. Agriculture, Ecosystem and Environment 113:326-335. https://doi.org/10.1016/j.agee.2005.10.012 [ Links ]
Mansour F. 1987. Spiders in sprayed and unsprayed cotton fields in Israel, their interactions with cotton pests and their importance as predators of the Egyptian cotton leaf worm, Spodoptera littoralis. Phytoparasitica. 15:31-41. https://doi.org/10.1007/BF02980517 [ Links ]
Mansour F., D. Rosen & A. Shulov. 1980. A survey of spider populations (Araneae) in sprayed and unsprayed apple orchards in Israel and their ability to feed on larvae of Spodoptera littoralis (Boisd.). Oecologia Applicata. 1(2): 189-197. [ Links ]
Marc, P. 1993. Intraspecific predation in Clubiona corticalis (Walckenaer, 1802) (Araneae, Clubionidae): a spider bred for its interest in biological control. Memoirs of the Queensland Museum. 33(2): 607-614. [ Links ]
Marc P. & A. Canard. 1997. Maintaining spider biodiversity in agroecosystems as a tool in pest control. Agriculture, Ecosysistems and Environment. 62: 229-235. https://doi.org/10.1016/S0167-8809(96)01133-4 [ Links ]
Marc P., A. Canard & F. Ysnel. 1999. Spiders (Araneae) useful for pest limitation and bioindication. Agriculture, Ecosy sistems and Environment. 74: 229-273. https://doi.org/10.1016/S0167-8809(99)00038-9 [ Links ]
Merrett P. & R. Snazell. 1983. A comparison of pitfall sampling and vacuum sampling for assessing spider faunas on heathland at Ashdown Forest, south east England. Bulletin of British Arachnology Society. 6: 1-13. [ Links ]
Moreno C. 2001. Métodos para medir la biodiversidad. M&T-Manuales y Tesis SEA. Zaragoza. Volumen 1. 84 pp. [ Links ]
Öberg S. & B. Ekbom. 2006. Recolonisation and distribution of spiders and carabids in cereal fields after spring sowing. Annals of Applied Biology. 149: 203-211. https://doi.org/10.1111/j.1744-7348.2006.00088.x [ Links ]
Öberg S., B. Ekbom & R. Bommarco. 2007. Influence of habitat type and surrounding landscape on spider diversity in Swedish agroecosystems. Agriculture, Ecosystems and Environment. 122: 211-219. https://doi.org/10.1016/j.agee.2006.12.034 [ Links ]
Pekár S. 2002. Susceptibility of the spider Theridion impressum to 17 insecticides. Journal of Pest Science. 75: 51-55. https://doi.org/10.1046/j.1439-0280.2002.02016.x [ Links ]
Pekár S. 2012. Spiders (Araneae) in the pesticide world: an ecotoxicological review. Pest Management Science. 68(11): 1438-46. https://doi.org/10.1002/ps.3397 [ Links ]
Pérez-Guerrero S., R. Tamajón, H. Aldebis & E. Vargas-Osuna. 2009. Comunidad de ara-as en cultivos de algodón ecológico en el sur de Espa-a. Revista colombiana de Entomología. 35(2): 168-172. [ Links ]
Plagens M. 1983. Population of Misumenops (Araneae, Thomisidae) in two Arizona cotton fields. Environmental Entomology. 12(2): 572-575.https://doi.org/10.1093/ee/12.2.572 [ Links ]
Quevedo G. & J. Vasconcellos-Neto. 2003. Natural history of Misumenops argenteus (Thomisidae): seasonality and diet on Trichogoniopsis adenantha (Asteraceae). Journal of Arachnology. 31(2): 297-304. https://doi.org/10.1636/02-19 [ Links ]
Rajput S., R. Dino, S. Ali, T. Ahmed & M. Shahid. 2012. Effects of pesticide on population reduction of spiders and sucking pests in cotton crop. Pakistan Journal of Entomology Karachi. 27(2): 99-104. [ Links ]
Rodrigues E., S. Mendonça & R. Ott. 2009. Spider diversity in a rice agroecosystem and adjacent areas in southern Brazil. Revista colombiana de Entomología. 35(1): 89-97. [ Links ]
Samu F. & G. Lövei. 1995. Species richness of a spider community (Araneae): extrapolation from simulated increasing sampling effort. European Journal of Entomoly 92:633-638. [ Links ]
Sarmiento M. 1992. Plagas del cultivo del algodonero. Universidad Nacional Agraria La Molina. Departamento de Entomología. Lima-Perú. 238 p. [ Links ]
Schmidt M. & T. Tscharntke. 2005. The rol of perennial habitats for Central European farmland spiders. Agriculture, Ecosystems and Environment 105: 235-242. https://doi.org/10.1016/j.agee.2004.03.009 [ Links ]
Sechser B., S. Ayoub & N. Monuir. 2001. Selectivity of Lufenuron (Match), Profenofos and mixtures of both versus cotton predators. Pesticides and Beneficial Organisms 24(4): 121-137. [ Links ]
Skinner R., B. 1974. The relative and seasonal abundance of spiders from the herb-shrub stratum of cotton fields and the influence of peripheral habitat on spider populations. Tesis para optar el Grado de Magister. Universidad Auburn. Alabama-Estados Unidos. [ Links ]
Solanki R. & D. Kumar. 2014. Effect of pesticides on spider population in cotton agro-system of Vadodara (Gujarat). Journal of Sol-Gel Science and Technology 3(1): 48-52. [ Links ]
Srikanth J., S. Easwaramoorthy, N. Kurup & G. Santhalakshmi. 1997. Spider abundance in sugarcane: impact of cultural practices, irrigation and post-harvest trash burning. Biological Agriculture & Horticulture 14: 343-
356. http://dx.doi.org/10.1080/01448765.1997.975-5169 [ Links ]
Sunderland K. & F. Samu. 2000. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: a review. Entomologia Experimentalis et Applicata 95: 1-13. https://doi.org/10.1046/j.1570-7458.2000.00635.x [ Links ]
Topping C. & M. Luff. 1995. Three factors affecting the pitfall trap catch of linyphiid spiders (Araneae: Linyphiidae). Bulletin of British Arachnology Society 10: 35-38. [ Links ]
Topping C. & K. Sunderland. 1992. Limitations to the use of pitfall traps in ecological studies exemplified by a study of spiders in a field of winter wheat. Journal of Applied Ecology 29(2): 485-491. https://doi.org/10.2307/2404516 [ Links ]
Correspondencia:
1 Universidad Nacional Mayor de San Marcos, Lima-Perú.
2 Universidad Nacional Mayor de San Marcos, Museo de Historia Natural, UNMSM, Apartado Postal 14-0434, Lima 14, Perú.
3 Universidad Nacional Agraria La Molina, Museo de Entomología "Klaus Raven Büller", Departamento de Entomología, Apartado 12-056, Lima-Perú.
*Autor para correspondencia
Email LC: laucruzmendoza@gmail.com
Email DS: diana.silva@unmsm.edu.pe
Email CV: cvc@lamolina.edu.pe
Presentado: 27/03/2018
Aceptado: 19/11/2018
Publicado online: 18/03/2019

