










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de la Sociedad Química del Perú
versão impressa ISSN 1810-634X
Rev. Soc. Quím. Perú v.78 n.3 Lima jul./set. 2012
TRABAJOS ORIGINALES
Purificación de una lectina tipo C del veneno de la serpiente peruana Lachesis muta
Purification of a lectin type c from the venom of Lachesis muta peruvian snake
Mercedes Palominoa , Fanny Lazoa , Julio Delgadilloa , Ruperto Severinob , Armando Yarlequéa*
a* Laboratorio de Biología Molecular, Facultad de Ciencias Biológicas, UNMSM, Av. Venezuela Cuadra 34 s/n, Lima- 1. e-mail: ayarleque48@gmail.com
b Laboratorio de Zoología de Invertebrados, Facultad de Ciencias Biológicas - UNMSM.
RESUMEN
A partir del veneno de la serpiente Lachesis muta, que habita en la amazonía peruana, se purificó y determinó las propiedades bioquímicas de una proteína no enzimática denominada lectina Tipo C. La purificación se logró a través de un único paso cromatográfico utilizando una columna de DEAE-Sephadex A-50 equilibrada con buffer acetato de amonio 0,1 M pH 5. La lectina fue reconocida debido a su capacidad para aglutinar glóbulos rojos humanos in vitro; por lo que se trata de una hemoaglutinina. La proteína en estudio fue purificada 3,6 veces y representó el 3,5 % del contenido proteico total del veneno. El análisis por PAGE-SDS, demostró que la lectina se encontraba al estado homogéneo con una masa molecular de 27, 5 kDa y 14,3 kDa en su forma reducida. La lectina fue inhibida en su actividad por la lactosa (1,56 mM), seguida de galactosa, arabinosa, sacarosa y glucosa. En cambio, no se registró inhibición con maltosa, manosa y fructosa. La capacidad hemoaglutinante de la lectina también fue inhibida por EDTA, seguida de DTT y 2-mercaptoetanol a distintas concentraciones; la inhibición causada por EDTA (0,125 - 0,5 mM), fue anulada por los iones Ca+2, Mg+2 y Mn+2. Los resultados indican que la proteína purificada es una lectina tipo C, la cual es una potente hemoaglutinina.
Palabras clave: Hemoaglutininas, veneno, serpiente, Lachesis muta.
ABSTRACT
A lectin type C was purified and researching its biochemical properties from the venom of Lachesis muta Peruvian snake. Purification was performed through a unic chromatographic step using a DEAE-Sephadex A-50 column, equilibrated with 0,1 M ammonium acetate pH 5 buffer. A lectin was recognized for its capacity in vitro to agglutinating human red cells. This protein was purified 3,6 folds and it represents 3,5 % of the total protein into the venom. It had 27,5 kDa and 14,3 kDa of molecular weight in non reduction and reduction conditions, respectively, using a PAGE-SDS analysis. It was strongly inhibited by the lactose (1,56 mM) followed by galactose, arabinose, sacarose and glucose. While maltose, mannose and fructose were not inhibitors. Hemagglutinating activity was inhibited by EDTA, DTT and ME too using different concentrations. However inhibition produced by EDTA (0,125-0,5 mM) was abolished by Ca+2, Mg+2 and Mn+2 ions. These results showed that this purified protein is a lectin type C, with a potent hemagglutinin activity.
Key words: Hemagglutinins snake, venom, Lachesis muta.
INTRODUCCIÓN
Lachesis muta, también conocida como "Shushupe", es el vipérido de mayor longitud de América, pudiendo alcanzar de 3 a 4 m1, habita la región tropical y sub tropical desde Costa Rica hasta el sur de Brasil. En el Perú es la segunda responsable de accidentes ofídicos2. El veneno de Lachesis muta posee diferentes componentes enzimáticos que han sido aislados y caracterizados desde la década del 70. Dentro de ellos se encuentra la enzima similar a trombina, fosfolipasa A, exonucleasa, L-aminoácido oxidasa, proteasas3 y hialuronidasa4. Tales enzimas están implicadas en el proceso de envenenamiento causando edema, hemorragia, hipotensión, desórdenes en la coagulación sanguínea, y necrosis3. Sin embargo, se han encontrado componentes no enzimáticos del tipo de las lectinas, incluyendo el veneno de Lachesis muta stenophyrs de centroamérica5,6.
Las lectinas encontradas en venenos de serpientes pertenecen a la superfamilia de lectinas del tipo C7, las cuales son proteínas dependientes de calcio para interactuar con carbohidratos, que se encuentran extracelularmente; presentan puentes disulfuros y no poseen un ligando sacárido típico7. Sin embargo, se han purificado y caracterizado un gran número de lectinas a partir de venenos de serpientes5,6,8. Estas proteínas pertenecen a la familia de lectinas tipo C y poseen una región conservada denominada dominio de reconocimiento de carbohidratos (CDR)9. Pueden dividirse en dos grandes grupos: 1) lectinas tipo C verdaderas, aquellas que se unen a carbohidratos y son dependientes de calcio y 2) proteínas similares a lectinas tipo C, aquellas que contienen un CDR pero que no se unen a carbohidratos ni son dependientes de calcio 10.
Estas proteínas son capaces de aglutinar in vitro glóbulos rojos de diferentes orígenes incluyendo aquéllos obtenidos de humanos11; presentan actividad edemática, actividad mitogénica sobre linfocitos, y pueden inducir agregación plaquetaria; ocasionan alteraciones renales o son capaces de activar neutrófilos11.
A pesar de exhibir diferentes actividades biológicas, las lectinas tipo C verdaderas, son consideradas como proteínas no tóxicas12 y por lo tanto presentan un potencial biotecnológico que ha estado siendo evaluado en los últimos años. En ese sentido, recientemente, se purificó y caracterizó la lectina del veneno de Bothrops leucurus encontrándose que esta proteína posee una actividad antimicrobiana efectiva contra bacterias Gram-positivas: Staphylococcus aureus, Enterococcus faecalis y Bacillus subtilis 13.
Por lo tanto, el presente trabajo ha tenido como objeto la exploración y purificación de una lectina procedente del veneno de Lachesis muta del Perú para su ulterior caracterización.
PARTE EXPERIMENTAL
Material biológico
Se empleó veneno de Lachesis muta de especímenes adultos procedentes de Pucallpa-Dpto. de Ucayali y mantenidos en cautiverio en el Serpentario "Oswaldo Meneses" del Museo de Historia Natural de la UNMSM. El veneno obtenido, fue liofilizado y conservado a 4 °C.
Cuantificación proteica
Se realizó por el método de absorción ultravioleta a 280 nm y por el método de Lowry et al., modificado por Loayza et al.14, usando como proteína estándar albúmina sérica bovina.
Actividad hemoaglutinante (AH)
El método usado fue el de Ogilvie y Gartner, 198415. En cada pocillo de una placa de microtitulación con fondo "V", se colocaron 50 µL de solución TCS (Tris 0,01 M, CaCl2 2 mM, NaCl 0,5 M) a pH 7 y luego se agregó 50 µL de veneno o proteína purificada al primer pocillo de cada fila, procediéndose a efectuar diluciones sucesivas ½ para los demás pocillos de cada fila. Por último, 50 µL de solución al 3% de glóbulos rojos humanos del grupo O Rh+ fueron añadidos a cada pocillo de las placas. Las lecturas se realizaron luego de una hora de incubación a temperatura ambiente. Los resultados fueron clasificados en hemoaglutinación completa, incompleta y no hemoaglutinación. Esta prueba, así como todas las pruebas de hemoaglutinación, en adelante fueron realizadas por triplicado y el título (última dilución que exhibe hemoaglutinación completa) hallado es un promedio de los tres resultados obtenidos.
Purificación de la lectina
50 mg de veneno de L. muta fueron disueltos en buffer acetato de amonio 0,1 M pH 5,0 y luego de separar los restos insolubles por centrifugación, la muestra fue aplicada a una columna de intercambio aniónico DEAE-Sephadex A-50 (38 x 1,1 cm) previamente equilibrada con el mismo buffer. Se colectó fracciones de 1 mL con un flujo de 10 mL/hora. La elución de la proteína retenida en la columna se realizó adicionando NaCl 0,3 M. Se determinó el contenido proteico y la actividad hemoaglutinante de todas las fracciones colectadas.
Evaluación de la pureza y determinación del peso molecular
La lectina purificada fue evaluada por electroforesis en geles de poliacrilamida al 12% (PAGE-SDS) en ausencia y presencia de 2-mercaptoetanol (ME), de acuerdo al método de Laemmli (1970)16. El peso molecular se determinó usando como marcadores: albúmina sérica bovina (66 kDa), ovoalbúmina (45 kDa) y lisozima (14,3 kDa).
Efecto de carbohidratos
Se evaluó D-galactosa, D- manosa, D- maltosa, L- arabinosa, D-fructosa, D-lactosa y D- sacarosa sobre la actividad hemoaglutinante. Se realizaron diluciones seriadas de los carbohidratos con concentraciones finales en el pocillo desde 200 hasta 0,05 mM. Luego se adicionó 1 µg de lectina purificada, cantidad que produce hemoaglutinación completa. Ambos componentes, azúcar más lectina, fueron incubados a 37 ºC por 20 minutos e inmediatamente se añadió glóbulos rojos al 3%. Se halló la cantidad mínima necesaria de cada azúcar para inhibir una hemoaglutinación completa. Los ensayos se realizaron por triplicado.
Efecto de agentes reductores y quelantes
Se evaluaó los efectos del 2-Mercaptoetanol (ME), ditiotreitol (DTT) y EDTA sobre la actividad hemoaglutinante de la lectina de L. muta. Se realizó diluciones seriadas de los agentes en placas de microtitulación: EDTA (0,75 - 2 mM), (ME) y DTT (0,75 – 10 mM). Se reemplazó la solución TCS (Tris 0,01M, CaCl2 2 mM, NaCl 0,5M) por solución TS (Tris 0,01 M, NaCl 0,5 M pH 7,3) en los ensayos que contenían EDTA. La lectina más el agente respectivo fueron incubados por 20 minutos a 37 ºC e inmediatamente se añadió glóbulos rojos al 3%.
Efecto de agentes químicos
De manera similar a la descrita para los agentes reductores, se realizó diluciones de iodoacetato y glutatión (0,75 – 10 mM). Enfrentándolas contra 1 µg de lectina por 20 minutos a 37 ºC.
Efecto de iones divalentes
Se preparó mezclas que contenían EDTA (0,125, 0,250 y 0,5 mM), iones divalentes bajo la forma de cloruros (0,05 – 1,5 mM) y lectina (1 µg). Se halló la concentración mínima (CMR) de cada ion capaz de reconstituir la AH anulada por EDTA.
RESULTADOS Y DISCUSION
Purificación de la lectina y peso molecular
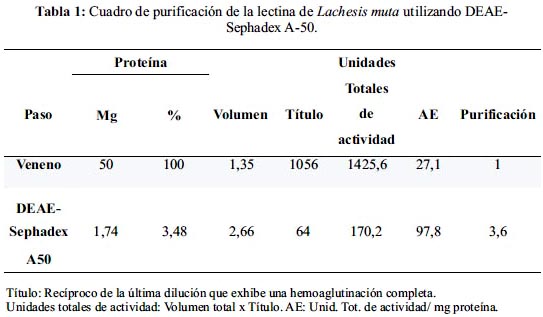
Al fraccionar el veneno de L. muta en DEAE-Sephadex A-50, se obtuvo dos picos principales de proteína, eluídos isocráticamente, lo que representó el 55,2% de la proteína total. Luego de aplicar NaCl 0,3 M, eluyeron dos picos adicionales que representaron el 44,8 %. La lectina correspondió al último pico obtenido en este paso (figura 1).
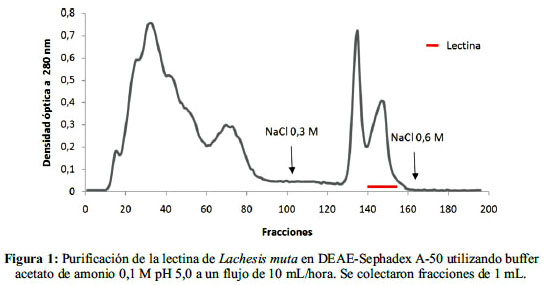
La actividad hemoaglutinante aumentó de 27,1 para la muestra inicial de veneno a 97,8 para la lectina purificada, lo que equivale a una purificación de 3,6 veces (tabla 1).Los títulos fueron: 1056 para el veneno y 64 para la lectina purificada (figura 2, tabla 1). Con esta metodología se recuperó 1,74 mg de lectina purificada que representa el 3,50%
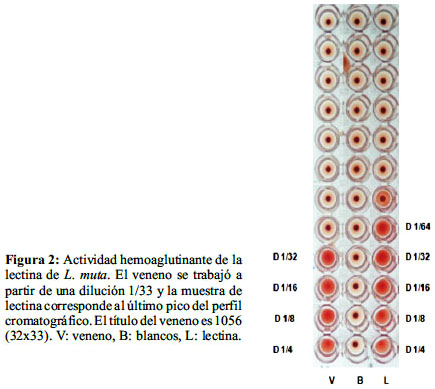
El presente trabajo es el primer reporte para el país sobre la detección y purificación de una lectina contenida en el veneno de Lachesis muta.
Al analizar la lectina por PAGE-SDS, se encontró una única banda proteica tanto en condiciones reductoras y no reductoras. En cuanto al peso molecular, corresponde a 14,3 kDa en condiciones reductoras y 27,5 kDa en condiciones no reductoras.
En 1980, Gartner et al17, aislaron una proteína del veneno de Bothrops atrox capaz de aglutinar glóbulos rojos de diferentes orígenes. Ellos la denominaron trombolectina, proteína clasificada dentro de la gran familia de lectinas debido a que su actividad aglutinante se inhibía en presencia de carbohidratos. Luego de este primer reporte, se han purificado y caracterizado varias lectinas a partir de venenos de vipéridos, crotálidos y elápidos 5.
En el presente trabajo, se ha purificado una lectina a partir de veneno liofilizado de Lachesis muta. Se trata de una proteína ácida, capaz de adherirse al DEAE-Sephadex A-50 y con un punto isoeléctrico menor a 5. Estos resultados son comparables a los obtenidos para la lectina de Lachesis muta aislada a partir de un veneno comercial.
Las lectinas tipo C verdaderas de venenos de serpientes, suelen purificarse empleando columnas de cromatografía de afinidad que contengan lactosa o galactosa como grupo de unión9,11,12; sin embargo, en esta investigación se ha logrado la purificación de una lectina verdadera empleando DEAE-Sephadex A-50 y eluyendo la proteína atrapada en el sistema con una solución de NaCl 0,3M. Hasta la fecha, ninguna lectina ofídica había sido purificada al estado homogéneo, mediante un solo paso cromatográfico; ya que más bien este tipo de metodología es común al trabajar con lectinas vegetales; tal es el caso de la lectina aislada mediante una columna de DEAE-Sephadex a partir de semillas de Eugenia uniflora L.18.
La lectina de Lachesis muta aislada en la presente investigación, es un homodímero de 27,5 kDa donde cada subunidad presenta un peso de 14,3 kDa. Los monómeros se disociaron en presencia del agente reductor 2-mercaptoetanol lo que sugiere la presencia de al menos un puente disulfuro intercatenario. Este comportamiento es usual en las lectinas ofídicas estando los pesos en su forma dimérica entre 28 kDa, como la trombolectina 17, y 32 kDa, como la lectina de Bothrops insularis 19. A pesar de que las lectinas tipo C verdaderas suelen ser diméricas, la estructura cristalográfica de la lectina de Crotalus atrox (RSL) muestra una estructura decamérica, con dos pentámeros arreglados simétricamente20.
Efecto de mono y disacáridos en la actividad hemoaglutinante
Al evaluar el efecto de carbohidratos sobre la actividad hemoaglutinante de la lectina en estudio se encontró que cinco de los carbohidratos probados (lactosa, galactosa, arabinosa, sacarosa y glucosa) son capaces de inhibir la hemoaglutinación y por ende son reconocidos por la lectina. En cambio, no se observó efecto alguno en los ensayos con maltosa, manosa y fructosa, empleando concentraciones de hasta 200 mM (tabla 2).

Las lectinas tipo C verdaderas son capaces de reconocer carbohidratos y se les considera calcio dependientes por presentar un dominio conservado para el ion9. Poseen un peso molecular de 30 kDa en promedio y está formada por dos subunidades idénticas unidas por un puente disulfuro. Cada una de estas subunidades presenta un dominio de reconocimiento de carbohidrato CRD, de aproximadamente 115-120 aminoácidos con 32 posiciones conservadas distribuidas a intervalos fijos, sobresaliendo cuatro cisteínas que forman dos puentes disulfuros intracatenarios.
Los experimentos presentados, en esta investigación muestran que la lactosa, a una concentración de 1,56 mM, es capaz de inhibir totalmente la actividad hemoaglutinante y si comparamos este efecto con el de la sacarosa, esta última sólo produce este efecto inhibidor a la concentración de 50 mm. Nótese que los valores de inhibición producida por la galactosa (3,1 mM) y la glucosa (100 mM) -que son los componentes de la lactosa- son distantes entre sí y el que más se acerca a la lactosa es el de la galactosa, por lo que puede deducirse que este monosacárido juega el rol más importante en la inhibición al integrarse en la molécula de la lactosa. En cambio, el otro disacárido estudiado, maltosa, no causa ningún efecto en la actividad hemoaglutinante.
A pesar de que la lectina en estudio puede reconocer más de un carbohidrato, exhibe mayor afinidad por la lactosa. El reconocimiento de carbohidratos recaería en los residuos de aminoácidos específicos dentro del CRD; por ejemplo, cuando la triada Glu185-Pro186-Asn187 (EPN) ubicada en la lectina de unión a la manosa de la rata es reemplazada por Gln185-Pro186- Asp187 (QPD), la proteína cambia su preferencia por manosa y se convierte en una proteína que une galactosa21. Este experimento es concordante con lo encontrado en las secuencias de lectinas tipo C de venenos ofídicos, donde la triada QPD se relaciona con proteínas que reconocen la galactosa.
Efecto de quelantes, iones divalentes y otros agentes químicos
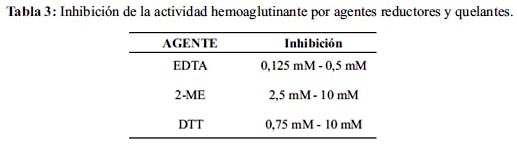
La AH causada por 1 µg de la lectina de L. muta es fuertemente inhibida por el EDTA, ME y DTT (tabla 3). El rango de EDTA capaz de inhibir completamente la AH es 0,125 - 0,5 mM. El ME causó inhibición desde 2,5 a 10 mM; mientras que el DTT inhibe la AH a concentraciones menores a 0,75 mM. Por otro lado, agentes como yodoacetato y glutation tuvieron un efecto negativo sobre los glóbulos rojos humanos a las concentraciones de 5 y 10 mM. Concentraciones inferiores no mostraron efecto alguno sobre la AH.
Los ensayos para reconstituir la AH inhibida por EDTA mostraron que los iones calcio, magnesio y manganeso son capaces de reconstituir dicha actividad con valores diferentes de CMR (tabla 4).El menor valor para reconstituir la actividad hemoaglutinante se obtuvo con el ion calcio (0,1 mM) mientras que con manganeso se requirió 1 mM para restablecer la HA.

La actividad hemoaglutinante de la proteína purificada fue inhibida por EDTA,DTT y ME; así como por cinco carbohidratos diferentes. Además, la AH previamente anulada por EDTA fue reconstituida por los iones calcio, magnesio y manganeso. Estos resultados muestran una dependencia de iones divalentes, especialmente calcio, para el reconocimiento del carbohidrato involucrado en la actividad hemoaglutinante.
Como ya se ha mencionado, las lectinas tipo C verdaderas tienen como característica resaltante el ser dependientes de iones calcio; he aquí el origen de la "C" que llevan en su nominación. Por muchos años los experimentos se limitaron a probar la reconstitución de la actividad hemoaglutinante por iones calcio, sin evaluar ningún otro ion divalente. Sin embargo, la actividad hemoaglutinante de la lectina de Bothrops jararacussu es reconstituida no sólo en presencia de iones calcio, sino también al emplear magnesio y manganeso11. Esta situación es idéntica a la encontrada para la lectina de Lachesis muta en esta investigación, siendo por tanto estas dos lectinas capaces de actuar en presencia de otros iones que no sean calcio.
Respecto a este punto, sería interesante realizar estudios cristalográficos con lectinas en presencia de iones divalentes diferentes a calcio para entender lo que sucede en estos nuevos escenarios.
CONCLUSIONES
-
Los resultados de este trabajo muestran en primer lugar, la presencia de proteínas no enzimáticas como la lectina tipo C purificada, la cual es una proteína ácida, homodimérica, dependiente de calcio y con un peso molecular de 27,5 kDa en condiciones no reductoras. La lactosa es el principal disacárido inhibidor de la AH y por tanto el de mayor afinidad por el sitio de reconocimiento de la lectina. Su dependencia de calcio fue establecida usando agentes quelantes y su reconstitución por este ion divalente, mostrándose, además, que otros iones como magnesio y manganeso son capaces de restablecer la actividad a concentraciones mayores.
AGRADECIMIENTO
Los autores agradecen al Consejo Superior de Investigaciones (CSI) del Vicerrectorado de Investigación (VRI) de la UNMSM por el apoyo financiero brindado. Uno de los autores (Palomino, M.) obtuvo su Título Profesional de Bióloga Genetista Biotecnóloga con parte de este estudio.
BIBLIOGRAFÍA
1. Meneses O. Los animales venenosos y sus peligros. Lima-Perú: Instituto de Salud Pública 1974; (2):3-4.
2. Zavaleta A. Mordedura de serpiente (ofidismo): Un problema de salud en el Perú. Rev Med Hered 2004;15(2):61-63.
3. Yarlequé A. Las serpientes peruanas y sus venenos. Lima Perú: Fondo Editorial UNMSM; 2000.
4. Hurtado L, Lerma R, Rodríguez E, Yarlequé A. Aislamiento y algunas propiedades bioquímicas de una hialuronato glicanohidrolasa del veneno de la serpiente Lachesis muta "Shushupe". Rev Soc Quim Perú 2007; 73 (4): 226-234.
5. Aragón-Ortiz F, Brenes-Brenes JR, Gubensek F. Characterization of a lectin-like protein isolated from Lachesis muta snake venom. Rev Biol Trop 1989; 37(1): 79-83.
6. Aragón-Ortiz F, Mentele R, Auerswald EA. Amino acid sequence of a lectin-like protein from Lachesis muta stenophyrs venom. Toxicon 1996; 34(7):763-769.
7. Gartner TK, Ogilvie ML. Isolation and characterization of three Ca2+ dependent beta- galactoside specific lectins from snake venoms. Biochem J. 1984; 224:301-307.
8. Gómez Leiva MA, Aragón-Ortiz F. Purification and some properties of a hemagglutinating protein mutina from bushmaster Lachesis muta snake venom. Rev Biol Trop 1986; 34(1): 49-53
9. Drickamer K. Two Distinct Classes of Carbohydrate-recognition Domains in Animal Lectins. J Biol Chem 1988; 263: 9557-9560.
10. Lu Q, Navdaev A, Clemetson JM, Clemetson KJ. Snake venom C-type lectins interacting with platelet receptors. Structure-function relationships and effects on haemostasis. Toxicon 2005; 45: 1089-1098.
11. Carvalho DD, Marangoni S, Oliveira B, Novello JC. Isolation and characterization of a new lectin from the venom of the snake Bothrops jararacussu. IUBMB Life 1998; 44:933–938.
12. Lomonte B, Rojas G, Gutierrez JM, Ramirez G. Isolation of a galactose-binding lectin from the venom of the snake Bothrops godmani (Godmann's pit viper). Toxicon 1990; 28;75-81.
13. Nunes ED, de Souza MA, Vaz AF, Santana GM, Gomes FS, Coelho LC. et al. Purification of a lectin with antibacterial activity from Bothrops leucurus snake venom. Comp Biochem Physiol B 2011; 159: 57-63.
14. Loayza S, Morante Y, Campos S, Yarlequé A. Enzimas proteolíticas en el veneno de la serpiente peruana Lachesis muta y Bothrops atrox. Rev Soc Quím Perú 1985; 52(3):151-163.
15. Ogilvie ML, Gartner TK. Identification of lectins in snake venoms. J Herpetol 1984; 18 (3): 285-290.
16. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970; 227(5259):680-685.
17. Gartner TK, Stocker K, Williams DC. Thrombolectin: a lectin isolated from Bothrops atrox venom. FEBS Lett 1980; 117: 13-16.
18. Oliveira MD, Andrade CA, Santos-Magalhães NS, Coelho LC, Teixeira JA, Carneiro-da- Cunha MG. et al. Purification of a lectin from Eugenia uniflora L. seeds and its potential antibacterial activity. Lett Appl Microbiol 2008; 46(3):371-376.
19. Guimarães-Gomes V, Oliveira-Carvalho AL, Junqueira-de-Azevedo IL, S Dutra DL, Pujol-Luz M, Castro HC. et al. Cloning, characterization, and structural analysis of a C- type lectin from Bothrops insularis (BiL) venom. Arch Biochem Biophys 2004; 432(1):1-11.
20. Walker JR, Nagar B, Young NM, Hirama T, Rini JM. X-ray crystal structure of a galactose-specific C-type lectin possessing a novel decameric quaternary structure. Biochemistry 2004; 43(13):3783-92.
21. Drickamer K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992; 360(6400):183-6.
Recibido el 24-07-2012
Aprobado el 16-08-2012
