










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkArnaldoa
versão impressa ISSN 1815-8242versão On-line ISSN 2413-3299
Arnaldoa vol.25 no.1 Trujillo jan./abr. 2018
http://dx.doi.org/http://doi.org/10.22497/arnaldoa.251.25103
ARTÍCULOS ORIGINALES
New combinations in Belloa J. Rémy and new diagnoses for Andean Lucilia Cass. and Mniodes (A.Gray) Benth. (Gnaphalieae, Asteraceae)
Nuevas combinaciones en Belloa J. Rémy y nuevos diagnósticos de Lucilia Cass. y Mniodes (A. Gray) Benth. (Gnaphalieae, Asteraceae) de los Andes
Michael O. Dillon
The Field Museum, Integrative Research Center, 1400 South Lake Shore Drive, Chicago, IL 60605, USA. mdillon@fieldmuseum.org
Abstract
Changes in the generic boundaries, as suggested by DNA sequence data, have altered concepts of Andean Asteraceae, specifically Belloa J. Rémy, Lucilia Cass., and Mniodes (A.Gray) Benth. (Gnaphalieae, Asteraceae). Over the past 30 years, numerous changes in classification and the resulting transfers have generated confusion. This paper provides a short history of these changes and current information for these genera as they exist in Andean South America. Further, other transfers are provided as suggested by results of sequence analysis data and newly proposed generic concepts: Belloa eriophora (J. Rémy) M. O. Dillon, comb. nov., and Belloa nivea (Phil.) M. O. Dillon, comb. nov. Changes in the diagnosis of Mniodes necessitates the following transfers: Mniodes argentea (Wedd.) M. O. Dillon, comb. nov., M. spathulifolia (Sagást. & M. O. Dillon) M. O. Dillon, comb. nov. and M. turneri (Sagást. & M. O. Dillon) M. O. Dillon, comb. nov.
Keywords: Asteraceae, Gnaphalieae, Chile, Peru, Andean flora.
Resumen
Los cambios en los límites genéricos, como sugieren los datos de secuenciación de ADN, han alterado conceptos en las asteráceas andinas, específicamente en Belloa J. Rémy, Lucilia Cass. y Mniodes (A. Gray) Benth. (Gnaphalieae, Asteraceae). En los últimos 30 años, numerosos cambios en la clasificación y las transferencias resultantes han generado confusión. Este documento proporciona una breve historia de estos cambios y la información actual para estos géneros tal como existen en los Andes de América del Sur. Además, se proporcionan varias transferencias como lo sugieren los resultados de análisis de secuenciación y nuevos conceptos genéricos propuestos: Belloa eriophora (J. Rémy) M. O. Dillon, comb. nov., y Belloa nivea (Phil.) M. O. Dillon, comb. nov. Los cambios en el diagnóstico de Mniodes requieren las siguientes transferencias: Mniodes argentea (Wedd.) M. O. Dillon, comb. nov., Mniodes spathulifolia (Sagást. & M. O. Dillon) M. O. Dillon, comb. nov. y Mniodes turneri (Sagást. & M. O. Dillon) M. O. Dillon, comb. nov.
Palabras clave: Asteraceae, Gnaphalieae, Chile, Perú, flora andina.
Introduction
The Gnaphalieae (Cass.) Lecoq & Juill. (Asteraceae) is a small tribe comprising ca. 185 genera (Ward et al., 2009) and perhaps 2000 species. In South America, some 20 genera and over 100 species are recorded with highest diversity in the tropical and subtropical Andean Cordillera (Dillon & Sagástegui 1991), and some have proliferated in upper elevation or alpine habitats (Dillon, 2005). These genera overwhelmingly possess at least some members with reduced habits form small cushions of tightly clustered pubescent stems and densely canescent leaves. Some genera look remarkably similar, both in the field and as collections on herbarium sheets. Experience within these groups has suggested that inspection of capitular and floral microcharacters are necessary for generic identity to be confirmed with certitude.
Biologists often distinguish between ‘convergent’ and ‘parallel’ evolution (Arendt & Reznick, 2008). The distinction between these two usually assumes that phenotypes are the result of the underlying genetic mechanisms; when the mechanisms are different and in distantly related taxa it is convergent, but when mechanisms are similar and in more closely related taxa it is termed parallel. Convergent evolution, such as the formation of cushion plants in high-elevation habitats within several different families, for example, Azorella compacta Phil. (Apiaceae), Mniodes aretioides (Schultz-Bip.) Cuatrec. (Asteraceae), and Pycnophyllum peruvianum Muschl. (Caryophyllaceae). The formation of cushions within different clades of the subtribe Gnaphaliinae could be viewed as parallel, similar forms in Mniodes (A. Gray) Benth. (South America), Pterygopappus Hoof.f. (Tasmania), and Raoulia Hook.f. (New Zealand), with each having derived from related genetic origins (Luebert et al., 2017, Nie et al., 2013). The distinction within the tribe would is best viewed as a continuum of convergence.
Generic boundaries have been controversial and none more so than the recognition and relationships of a suite of genera in the Lucilia-group, an informal gathering first proposed by Merxmüller et al. (1977). The recognition and constitution of Belloa J. Rémy and Lucilia Cass. has been an ongoing classification saga (Dillon 1990, Luebert et al., 2017). The description of additional genera, e.g., Luciliocline Anderb. & S. E. Freire and Gamochaetopsis Anderb. & S. E. Freire, has been confusing even to the most astute student of the Asteraceae. Further, in response to sequence data, Friere et al. (2014) transferred of all Luciliocline, and some Lucilia and some Belloa into a greatly expanded Mniodes Wedd. The generic delimitation or boundaries of Belloa and Lucilia have been expanded (Cabrera, 1958; Freire, 1986; 1987a,b) and contracted (Anderberg & Freire 1991, Dillon, 2003; Freire et al., 2014). Recently, on the strength of molecular studies (Urtubey et al., 2016), Lucilia alpina (i.e., Gamochaetopsis) has been transferred to Gamochaeta Wedd.
Materials and methods
Herbarium material was investigated from the following herbaria: including B, BM, COL, CONC, F, GH, HAO, HSP, HUT, K, M, MO, NY, P, SGO, TEX, US, and W. Dried herbarium material was used for measurements and descriptions of achene surface structures. Vouchers were listed in Luebert et al., 2017 (cf. Appendix 2). Herbarium acronyms follow Thiers (2017).
Results and discussion
A. Chilean Belloa J. Rémy (1847)
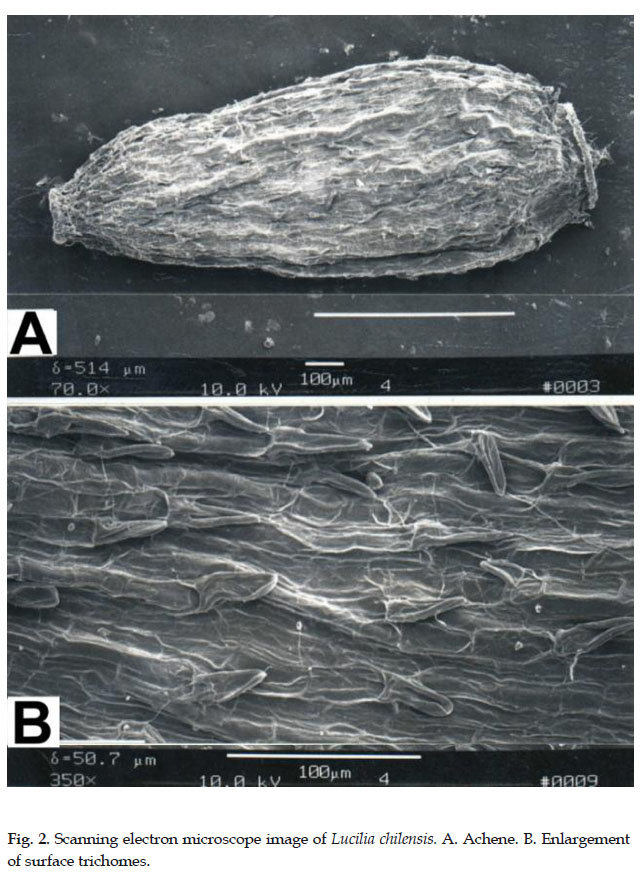
In 1835, William Joseph Hooker and George Walker Arnott published Lucilia chilensis Hook. & Arn. based upon a collection by Hugh Cumming 342 from Chile. De Candolle (1838) recognized the distinctness of Lucilia chilensis when he established Lucilia section Lucilioides based upon Lucilia chilensis. J. Rémy (1847) described the genus Belloa to contain Belloa (Lucilia) chilensis, a single species with pubescent achenes but with much smaller trichomes, only 40–50 µm long. He was aware of the genus Lucilia, also having described Lucilia eriophora J. Rémy (1847) from central Andean Chile in the same publication. The latter species has achenes pubescent with elongate twin hairs or Zwillingshaares (Hess 1938), the type and size of trichomes more similar to other Lucilia species with twin hairs > 150 µm long.
Cabrera (1958) recognized Belloa as distinct from Lucilia Cass. and transferred six species that had been in Gnaphalium L., Lucilia or Merope Wedd. Subsequently, Cabrera (1958) expanded his concept of Belloa, transferring two more species and describing two new species from Peru and Argentina. He commented that Belloa was distinct and easily distinguishable from Lucilia principally through their achenial trichomes. In addition to the strikingly different pubescence of the achenes, several other important characteristics separated these two genera, including the apices of the style branches of hermaphroditic florets and pappus characteristics. One of the species that Cabrera initially recognized as a Belloa, B. erythractis (Wedd.) Cabrera, was subsequently transferred to Gamochaeta Wedd. on the basis of its unique, sessile achenial trichomes (Cabrera 1978, Urtubey et al., 2016).
Freire (1986) provided a monograph for Lucilia and expanded its generic limits to include all taxa previously accorded to Belloa (Cabrera 1958; Sagástegui & Dillon 1985) with the statement that the type of achenial trichomes had no phylogenetic significance and provided no distinction between Belloa and Lucilia. She had based her classification system upon phenetic cladistic analysis (Freire 1984) and subsequently published another phenetic cladistic analysis (Freire 1987b) to support her expanded Lucilia. Freire (1987a) published a paper where she synonymized several of the newly described species proposed by Sagástegui & Dillon (1985). She argued for an expanded generic concept for Lucilia; a diagnosis that allowed for a genus with component species containing at least two fundamentally different types of achenial trichomes, i.e., some species possessing biseriate, capitate-glandular trichomes and others possessing elongate twin hairs or Zwillingshaares (Hess, 1938). Under the discussion, Freire maintained that the types of achenial trichomes were trivial and had no importance in the systematics of the group.
Dillon (1988, 1990) presented data suggesting that the two different types of trichomes represented in some Belloa (pro parte) and those of Lucilia (sensu stricto) were the products of quite complex ontogenetic patterns that were fundamentally different. These ontogenetic developments are not reversible ontogenetic endpoints, i.e., one type of trichomes could not revert or transform into the other. There appears to be no capacity to reverse ontological developments as illustrated by Hess (1938).
During morphological investigations, Dillon and Sagástegui (1991) provided the observation that a unique type of achenial trichomes was to be found in both Mniodes and in some members of Belloa (sensu lato), i.e., not like those of the type species but rather like those eventually become Luciliocline Anderb. & S. E. Freire and more recently Mniodes (Freire et al., 2014). Dillon (2003) accepted Luciliocline, concluding that Chilean Belloa (B. chilensis) were distinct and easily separated from Luciliocline. Further, Dillon (2003) suggested that Mniodes and Luciliocline shared close relationships, as both possessed similar habits, leaves, and floral morphology as well as the achenial trichomes.
Anderberg and Freire (1991) published a morphological cladistic analysis where Lucilia was dismantled, parsing the various species into four genera, i.e., Lucilia (8 spp.), Belloa (9 spp.), Luciliocline (5 spp.), and Gamochaetopsis (1 sp.). Surprisingly, their generic diagnosis for both Belloa and Luciliocline contained clearly non-parsimonious mixtures of achenial trichome types by maintaining Lucilia species within their concept of Belloa, [e.g. Lucilia kunthiana (DC.) Zardini, L. lehmannii Hieron.]. They amended the generic description of Belloa to include a mixture of the types of achenial trichomes with six Luciliocline species considered as Belloa.
Dillon (2003) provided transfers for eight Belloa species to Luciliocline. In Dillon’s generic diagnosis, all species Luciliocline were diagnosed by achenes covered with capitate-glandular trichomes or rarely, with trichomes only near the carpopodium. He recognized B. kunthiana as a legitimate member of Lucilia and placed B. lehmannii under the synonymy of the former species; both these taxa have achenes possessing elongate, twin hairs > 150 µm long. Dillon (2003) had considered Belloa chilensis (Hook. & Arn.) J. Rémy to be monotypic and with trichomes representing greatly reduced twin hairs and not similar to the capitate-glandular achenial trichomes possessed by Luciliocline and some Mniodes. However, these two species have well-developed twin hairs.
Although, gross morphology was not unlike other Lucilia species, L. eriophora and L. nivea, are here transferred to Belloa on the strength of DNA results and biogeography (Luebert et al., 2017; Nie et al., 2013). The Chilean species previously attributed to Lucilia, i.e., L. nivea and L. eriophora are recovered in well-supported clade with Belloa chilensis. The apical cells of the trichomes of Belloa chilensis are uniformly less than 50 µm long (Dillon 2003).
These Chilean taxa are isolated from the clade containing Lucilia s.s. Freire et al. (2014) interpreted Belloa as monotypic and placed L. araucana Phil., L. eriophora and L. nivea into synonymy of an expanded Belloa. Luebert et al. (2017) maintained these morphologically distinct taxa and Lucilia eriophora and L. nivea are here transferred to join a re-circumscribed Belloa. The new diagnosis of Belloa includes the presence of two sizes of trichomes, elongate, twin hairs >150 µm long in L. eriophora and L. nivea, and smaller twin hairs uniformly <50 µm long in Belloa chilensis.
Lucilia araucana Phil. (1895) is perhaps best placed in the synonymy of L. nivea on the strength of overall comparative morphology. It may be observed that L. araucana tends to form more compact cushions, however, field observations have confirmed that growing positions can affect overall phenotypes and especially shortening or expanding internode length. These two species share elongated achenial trichomes and further studies are needed to confirm taxonomic status.
1. Belloa chilensis (Hook. & Arn.) J. Rémy, Basionym: Lucilia chilensis Hook & Arn., Comp. Bot. Mag. 1: 102. 1835. TYPE: Chile, Región V (Valparaíso), Valparaíso, H. Cuming 342 (lectotype: K000500443; isolectotypes: K00050439, P00704659). Fig. 1., 2.

2. Belloa eriophora (J. Rémy) M. O. Dillon, comb. nov., Basionym: Lucilia eriophora J. Rémy, Fl. Chil. 3 (3): 335. 1847. TYPE: Chile, Región VI (Libertador General Bernardo O’Higgins), Prov. Colchagua, Yaquil, "aprili 1839", C. Gay 243 (holotype: P , FM neg. 37818). Fig. 3.

3. Belloa nivea (Phil.) M. O. Dillon, comb. nov.,Basionym: Chevreulia nivea Phil., Anales Univ. Chile 21: 380. 1862. TYPE: Chile, Región VIII (Bio-Bio), Concepcion, Nevado de Chillan, Cerro de Azufre, R.A. Philippi s.n. (holotype: SGO#64455). Lucilia nivea (Phil.) Cabrera, Notas Mus. La Plata, Bot. 7: 118. 1942.
B. Andean Lucilia Cass. (1817)
Molecular studies (Luebert et al., 2017, Nie et al., 2013) confirm that Andean Lucilia, i.e., L. kunthiana and L. conoidea, are recovered in a clade with other members of the genus from Brazil, and Berroa, Facelis, and Micropsis. These high-elevation taxa with greatly reduced habits have been recorded from throughout the Andean Cordillera from Venezuela to Argentina and Chile. Therefore, Lucilia is a genus of nine species distributed throughout South America, but with a center of diversity in Argentina, Bolivia, southeast Brazil, Uruguay and Paraguay; two are known from Peru and a single species Lucilia kunthiana recorded from Venezuela, Colombia, Ecuador, Peru, Bolivia, Argentina, and Chile.
The generic limits between Lucilia and Belloa have been controversial (see above). The gross appearance of some members of each genus is quite similar and especially those which occupy high elevation Andean habitats. Cassini’s original description of Lucilia described sericeous achenes; however, Freire (1986; 1987a) expanded the genus and transferred several species possessing achenes with biseriate, multicellular, capitate-glandular trichomes into Lucilia. Sagástegui & Dillon (1985) removed from Lucilia all members with glandular achenes and placed them in Belloa. Eventually, the genus Luciliocline was proposed for the majority of species in Belloa (Dillon, 2003). In the present treatment, only those taxa with consistently sericeous achenes are included in Lucilia. The extra-Andean taxa are restricted to more eastern South America and at altitudes below 3000 m; Lucilia acutifolia (Poir.) Cass., L. ferruginea Bak., L. linearifolia Bak., L. lycopodioides (Less.) S. E. Freire, L. nitens Less., and L. tomentosa Wedd.
Key to Species of Andean Lucilia
1. Habit not cespitose; stems weakly erect, 6-18 cm tall, leaves linear-oblong, 2-6 mm long, 0.7-1 mm wide, recurved; capitula 9-11 mm tall .............................. L. recurva
1’. Habit cespitose; stems in dense cushions, linear-obovate to spathulate, 5-20 mm long, 0.8-2 mm wide, blades not recurved but occasionally plane to falcate; capitula (5-)7-8 mm tall ..........2
2. Leaves spatulate, plane to falcate, 10-20 mm long, 1-2 mm wide, both surfaces tomentose ................................. L. kunthiana
2’. Leaves linear, pleated or canaliculate, 5-15 mm long, 0.8-1 mm wide, lower surface arachnoid-tomentose, glabrescent, upper surface densely silvery-tomentose ........... L. conoidea
1. Lucilia conoidea Wedd., Chlor. Andina 1: 154. 1856. TYPE: Peru, Dept. Cuzco, "Cordilleres du departement de Cuzco," C. Gay 1355 (holotype: P00704661, fragment, F). Fig. 4, 5.


Lucilia conoidea is known from southern Peru and northern Bolivia. It approaches L. kunthiana in general appearance but is distinctive in having leaves that are pubescent on the upper surface and glabrescent below, the reverse of the usual condition in Lucilia. This morphological distinction has a restricted geographic range in southern Peru and adjacent Bolivia. Lucilia conoidea was reduced to synonymy of L. kunthiana (as L. pusilla) by Freire (1986).
2. Lucilia kunthiana (DC.) Zardini, Ann. Missouri Bot. Gard. 74: 431. 1987.
Conyza kunthiana DC., Prodr. 5: 379. 1836. (nomen novum for Conyza pusilla Kunth). Gnaphalium kunthianum (DC.) Kuntze, Revis. Gen. PL, 3: 152. 1898. Mniodes kunthiana (DC.) S. E. Freire, Chemisquy, Anderb. & Urtubey, Pl. Syst. Evol. 301 (4): 1245. 2014.
Conyza pusilla Kunth, Nov. Gen. Sp. (fol.) 4: 54. 1820. TYPE: Ecuador, Prov. Pichincha, "crescit regione alpina Regni Quitensis," A. Humboldt & A. Bonpland s.n. (holotype: P00322293; isotype: P00704669, FM neg. 15126). Non Houttuyn (1779). Lucilia pusilla (Kunth) Hieron., Bot. Jahrb. Syst., 29. 29. 1900. Fig. 6.

Gnaphalium depressum Nutt., Trans. Amer. Philos. Soc., n.s. 7: 405. 1841. TYPE: Ecuador, Prov. Pichincha, W. Jameson s.n. (holotype: PH-01085444, img 00012252).
Lucilia lehmannii Hieron., Bot. Jahrb. Syst. 19: 51. 1895. TYPE: Ecuador, Prov. Azuay, Paramo del Cajas, 3600-4200 m, F. C. Lehmann 4649 (holotype: B, probably destroyed, F neg. 15126; isotype: NY00214769). Mniodes lehmannii (Hieron.) S. E. Freire, Chemisquy, Anderb. & Urtubey, Pl. Syst. Evol. 301 (4): 1245. 2014.
Lucilia affinis Wedd., Chlor. Andina 1: 230. 1857. TYPE: Bolivia, La Paz, Cordilleres du departement de la Paz, H. Mandon 168 (lectotype: K000500449; isolectotypes: G00223212, G00223213, P02533964, P02533965, P02533966, P02533958).
Lucilia violacea Wedd., Chlor. Andina 1: 155. 1855[1856]. TYPE: Bolivia, Cordillera de Sorata, Weddell s.n. [biz 3000] (holotype: P00704674).
Lucilia venezuelensis Steyer., Fieldiana, Botany 28: 644, f. 140. 1953. TYPE: Venezuela, Merida, below El Aguila and Chachopo, 3650-3965 m, 13 Mar 1944, J. A. Steyermark 55657 (holotype: F-1390433; isotypes: NY00214771, US-2046674, VEN).
Lucilia kunthiana is distributed throughout high elevation Andean habitats from Venezuela, Colombia, Ecuador, Peru, Bolivia, and extreme northern Chile; 3000-4800 m. Many regional species have described for cespitose, highly reduced plants with canescent leaves and achenes densely pubescent.
Schultz-Bip. (1856) proposed Gnaphalium evacoides based upon a plant collected from Venezuela by Moritz 1407 (P); however, the name was not validly published. Steyermark (1953) concluded that it was identical to Gnaphalium radians Benth. At the same time, Steyermark transferred Gnaphalium radians Benth. to Lucilia, placing G. evacoides Schultz-Bip. under its synonymy; Bentham’s type material was collected by K.T. Hartweg 1146 (K) in Colombia. Steyermark mentioned that G. radians had been confused with Lucilia kunthiana (as L. pusilla), a species originally collected in Ecuador by Bonpland and Humboldt and described in Conyza. De Candolle (1836) provided a new name, Conyza kunthiana due to the presence of an earlier homonym, Conyza pusilla Houtt 1779.
Weddell (1855) misinterpreted Bentham’s Colombian Gnaphalium radians and Knuth’s Ecuadorian species, Conyza pusilla, combining them under one name, Merope kunthiana (DC.) Wedd. The same error was perpetrated by Hieronymus (1900) as he combined all under one name, Lucilia pusilla.
Steyermark (1953) stated that Bentham’s Gnaphalium radians and Conyza pusilla Kunth were entirely different species and recognized by their achenes, with the former being glabous and the latter being densely pubescent with elongate, biseriate trichomes (Zwillingshaares). From this material, Steyermark described Lucilia venezuelensis and related it to Lucilia kunthiana, a species with densely pubescent achenes with elongate (>150 µm) biseriate trichomes distributed from Colombia to Argentina.
2. Lucilia recurva Wedd., Chl. Andina, 1: 156, tab. 25B, 1856. TYPE: Bolivia. Dept. Chuquisaca, Prov. Tomina, "sur les rochers nus, près du sommet du mont Curi", 3500 m, H. Weddell 3742 (lectotype: P00704670; isolectotype: P00704671). Fig. 7.

Lucilia recurva is recorded from dry sites in Bolivia (Cochabamba, Chuquisaca, Tarija) and Argentina (Salta), most frequently at 2500-2800 m. As the species epithet implies, its oblong leaves are strongly recurved. It is noteworthy that unlike the close relationship between L. conoidea and L. kunthiana (sister taxa), the relationship of L. recurva apparently with Brazilian taxa, i.e., L. acutifolia and L. lycopodioides (Luebert et al., 2017, Nie et al., 2013).
C. Laennecia alpina Poepp. (1845)
Laennecia alpina was proposed by Poeppig (1845) for a small herb with obovate or suborbicular leaves and achenes with pilose pubescence collected in southern Chile (Araucanía, Los Lagos, Magallanes and Chilean Antarctica). Cabrera (1961) considered it to be congeneric with Lucilia [i.e., L. alpina (Poepp.) Cabrera]. He stated that it could be confused vegetatively with Gamochaeta nivalis (Phil.) Cabrera, but suggested that it was easily distinguished from the latter taxon by its pubescent achenes.
Anderberg and Freire (1991) based their new genus, Gamochaetopsis Anderb. & S. E. Freire, upon Laennecia alpina. In an examination of the type collection at W, it was observed that the achenial trichomes elaborated in Gamochaetopsis are essentially reduced clavate trichomes (ca. 60 µm long) and quite unlike the sessile, biseriate trichomes elaborated in all authentic Gamochaeta (< 20 µm). In the analysis of resulting from sequence data, both Urtubey et al. (2016) and Luebert et al. (2017) found Gamochaetopsis as resolved as nested in a clade also including Gamochaeta and Stuckertiella Urtubey et al. (2016) transferred L. alpina from Gamochaetopsis to Gamochaeta. In an effort to document the morphology of reproductive characters of authentic material, the holotype collection housed at W (Vienna, Austria) E. F. Poeppig 889 (W0028920, Fig. 8) was investigated. Achenes were prepared for SEM photographs, including pappus and achenial trichomes (Fig. 9). Our observations point to inconsistencies between the morphology of the type collection, the protolog for the species by Poeppig (1845), and subsequent generic description by Anderberg and Freire (1991).

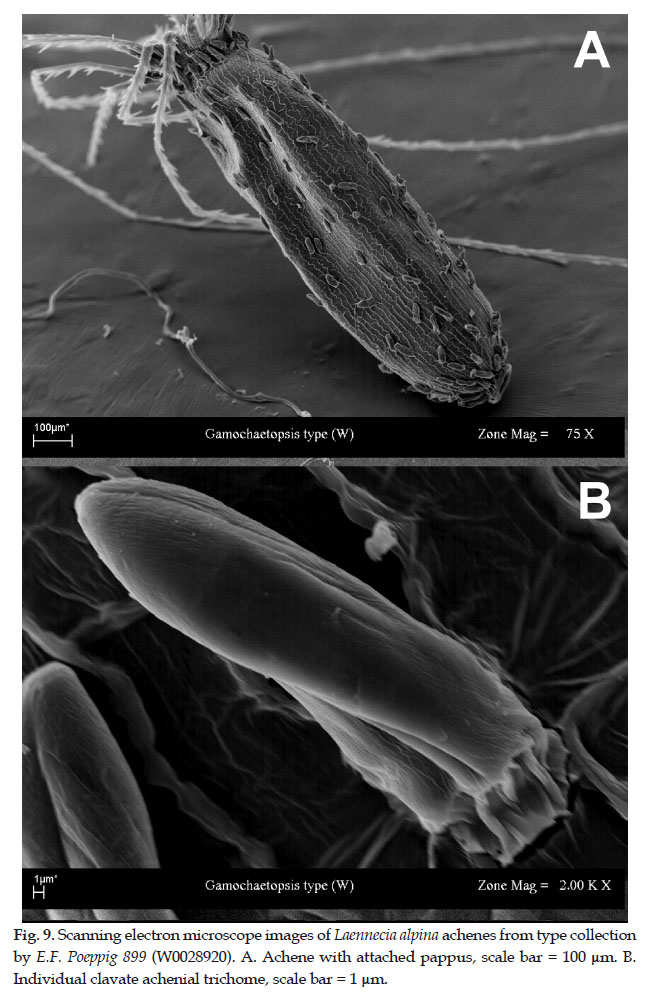
Gamochaeta alpina (Poepp.) S. E. Freire & Anderb., In: E. Urtubey, et al., 2016. New circumscription of the genus Gamochaeta (Asteraceae, Gnaphalieae) inferred from nuclear and plastid DNA sequences. Pl. Syst. Evol. 302: 1062. Basionym: Chile. "Chile australis rupium fissuris montis Pico de Pilque in Andibus Antuensium, Januario florebat", 1829, E.F. Poeppig 889 (holotype: W0028920; isotype: G-DC, G00499745).
D. Mniodes (A.Gray) Benth. (1873)
Freire et al. (2014) found Mniodes and Luciliocline Anderb. & S. E. Freire in the same clade, and transferred an array of species into Mniodes, a genus traditionally composed of strictly dioecious species. Luebert et al. (2017) also recovered members of Mniodes and Luciliocline, a connection that had been suggested by similarity in achenial trichomes (Dillon & Sagástegui 1991). This new generic concept requires additional taxa from Luciliocline be transferred into monophyletic group circumscribed by both morphological and molecular markers. While the Lucilia-group is recovered in the same large New World clade, neither Mniodes nor Luciliocline are closely related to Lucilia or Belloa. A few highly reduced Mniodes species approach Raouliopsis S. F. Blake (Colombia) in general compact habit and reduced tightly clustered leaf arrangements. The latter genus is recorded from extreme northern South America and diagnosed with compact habits and strictly dioecious breeding systems; molecular systematic studies have not been successful.
With a monograph of Mniodes in preparation, including several new species, the following combinations are necessary as a new diagnosis of Mniodes is adopted.
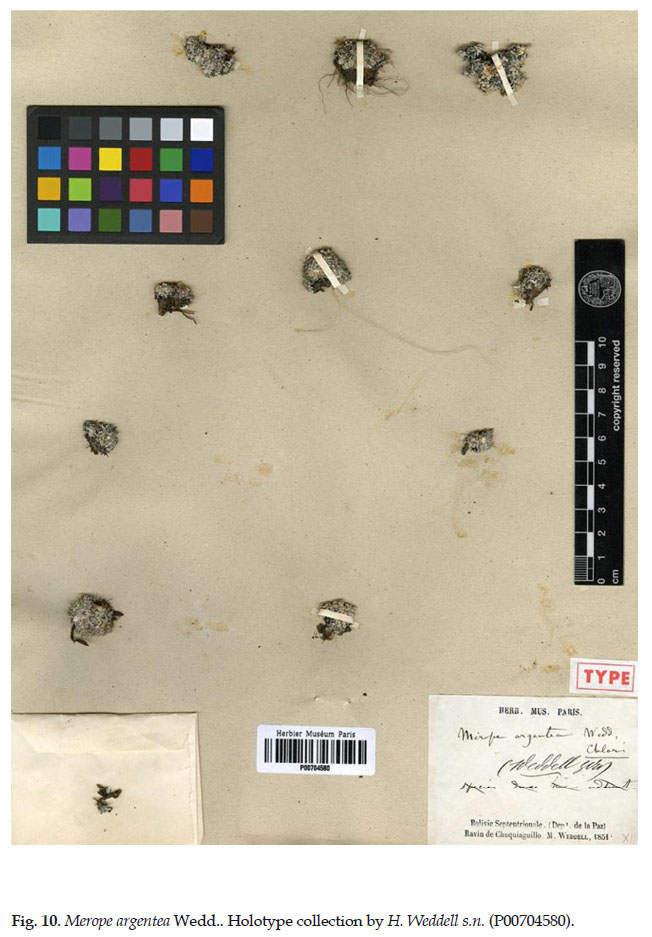
1. Mniodes argentea (Wedd.) M. O. Dillon, comb. nov. Basionym: Merope argentea Wedd., Chl. And. 1: 163. 1856. TYPE: Bolivia, Dept. La Paz, Quebrada de Chuquiaguillo, H. Weddell s.n. (holotype: P00704580). Fig. 10.
Freire (1986) placed Belloa argentea into the synonymy of Belloa schultzii with the statement that the presence of papillose achenes was not sufficient to recognize the species. Ariza (1989) accepted Belloa schultzii and distinguished it from all other Argentine species as having a cushion-form habit and glabrous achenes. While potentially related to M. schultzii, a position supported by molecular sequence data (Luebert et al., 2017), morphological differences allow discrimination of M. argentea from its sister species, M. schultzii; the former species has glabrous upper leaf surfaces.
Weddell (1855) described Merope argentea, M. schultzii Wedd., M. virescens Wedd., and M. caespititia Wedd. consecutively and in this order (p. 163–164). The first two, L. argentea and L. schultzii, are each accepted and treated as distinct; the former with smaller habit and achenes pubescent with capitate-glandular trichomes. Cabrera (1978) recognized L. argentea as distinct from L. virescens (a synonym of L. schultzii) based upon the latter’s larger leaves and pubescent achenes. In molecular studies, nrDNA data suggests these two species are sister taxa (Luebert et al., 2017).
2. Mniodes spathulifolia (Sagást. & M. O. Dillon) M. O. Dillon, comb. nov. Basionym: Belloa spathulifolia Sagást. & Dillon, Phytologia 58 (6): 394. 1985. TYPE: Peru, Dept. La Libertad, Prov. Santiago de Chuco, A. Sagástegui A., J. Mostacero L., & M. Diestra Q. 11695 (holotype: HUT; isotypes: F1947608, FM neg. 061880, MO). Fig. 11.

Freire (1987b) placed Belloa spathulifolia under the synonymy of Lucilia santanica (Cabrera) S. E. Freire, overlooking differences in habit, capitulescence, floral number, ecological preferences, and geographic distribution (Sagástegui & Dillon 1985). It is perhaps difficult for systematists to evaluate species without herbarium and field observations. Based upon data from several sources, we reject the suggestion by Freire that this distinctive taxon is a synonym of Mniodes (Luciliocline) santanica, a species with distinct morphology and distributed nearly 2500 kms to the south in northern Argentina.
While discussing Mniodes santanica, it is noteworthy that Anderberg and Freire (1991) also listed Gnaphalium depressum Nutt. and Gnaphalium sarmentosum Klatt as synonyms of M. santanica. An examination of type material shows these taxa are essentially unrelated to Mniodes (Luciliocline) santanica or any member of Mniodes. Gnaphalium depressum (W. Jameson s.n., PH00012252) from Andean Ecuador, is a synonym of Lucilia kunthiana (DC.) Zardini; it possesses achenes with elongate trichomes typical for Lucilia, i.e. 250–850 µm long. Gray (1862) also commented that the Ecuadorian Gnaphalium depressum Nutt. was not synonymous with Gnaphalium radians (i.e., Mniodes radians (Benth.) S. E. Freire, Chemisquy, Anderb. & Urtubey, 2014), and Gray suggested it best related to Lucilia kunthiana or L. conoidea Wedd.
Freire annotated an isotype of Gnaphalium sarmentosum Klatt (C. B. Pringle 4249, S-G-2896) as Luciliocline santanica. An examination of isotype material of Pringle 4249 (MEXU01216408) from Mexico shows it to be the type of Mexerion Nesom (1990). Its achenes possess reduced twin hairs with enlarged adaxial basal cells and short, blunt, apical cells ca. 50 µ long, very different from the capitate-glandular achenial trichomes found in Mniodes santanica and most Mniodes species. Not only does the Mexican material not correspond to the morphology of M. santanica, the two populations would exhibit an amphitropical disjunction of over 6000 kms. The published results of Nie et al. (2013) and Luebert et al. (2017) both recover this taxon in a clade with Gnaphaliothamnus Kirp., far removed from Mniodes (Dillon & Luebert 2015).
3. Mniodes turneri (Sagást. & M. O. Dillon) M. O. Dillon, comb. nov. Basionym: Belloa turneri Sagást. & Dillon, Phytologia 58 (6): 392. 1985. TYPE: Peru. Dept. Cajamarca, Prov. Contumazá, A. Sagástegui A., E. García A., S. López M., & J. Mostacero L. 10087 (holotype: HUT; isotypes: F1906100, FM neg 061881, MO, TEX). Fig. 12.

Freire (1987b) placed B. turneri under the synonymy of B. lopezmirandae with the statement that nothing separated them except the length of the corollas, number of phyllaries, and number of florets per capitula. Freire mentioned that these characters were of no significance and likely represented populational variability. I have examined these species in detail, both in the herbarium and in the field, and I continue to accept our Belloa turneri as distinct species, just as when it was originally published (Sagástegui & Dillon 1985). It is a species that represents a morphologically and ecologically distinct entity and, as with several other of her efforts at synonymy, we reject Freire’s contention that these species are conspecific. It is moved yet again to satisfy current generic concepts.
Acknowledgements
I thank curators and collection management staff for the loan of materials and hosting herbarium visits to consult collections, including B, BM, COL, F, GH, HAO, HSP, HUT, K, M, MO, NY, P, SGO, US, and W. I thank Federico Luebert for sharing information that suggested the taxonomic changes presented here and his SEM photographs of Poeppig 889 from W. I thank Daniel Le for providing the digital images of Field Museum collections. I thank Daniel Montesinos for the use of the photograph of Lucilia conoidea (Figure 5) taken in the field near Abra Málaga, Department of Cusco.
Contribution of the authors
M.D.: Redaction of the text, evaluation methodology, fieldwork, taxonomic determination of the species, review and approval of the final text.
Conflicts of interest
The author declares not to have conflicts of interest.
Literature cited
Anderberg, A. A. & S. E. Freire. 1991. A cladistic and biogeographic analysis of the Lucilia group (Asteraceae, Gnaphalieae). Bot. J. Linn. Soc. 106: 173–198. [ Links ]
Arendt, J. & D. Reznick. 2008. Convergence and parallelism reconsidered: what have we learned about the genetics of adaptation? Trends in Ecology & Evolution, 23: 26-32. [ Links ]
Cabrera, A. L. 1958. El Género "Belloa" Remy, Bol. Soc. Argent. Bot. 7: 79-85. [ Links ]
Cabrera, A. L. 1978. Compositae. Flora de la Provincia de Jujuy. Colecc. Cient., Inst. Natl. Teen. Agropec. 13: 288-293. [ Links ]
Cabrera, A. L. 1961. Observaciónes sobre las Inuleae–Gnaphalineae (Compositae) de América del Sur. Boletín de la Sociedad Argentina de Botánica 9: 359–386. [ Links ]
Cassini, H. 1817. Aperçu des Genres nouveaux formés par M. Henri dans la famille des Synanthérées. Bull. Sci. Soc. Phil Paris 3: 32. [ Links ]
De Candolle, A. P. 1838. Lucilia: 46. In: Prodromus systematis naturalis regni vegetabilis, 7. Paris. [ Links ]
Dillon, M. O. 1990. A tale of two genera: the implication of character choices and outgroup selection in cladistic analysis. American Journal of Botany Abstracts, 77: 129. [ Links ]
Dillon, M. O. 2003. New combinations in Luciliocline with notes on South American Gnaphalieae (Asteraceae). Arnaldoa, 10: 45–60. [ Links ]
Dillon, M. O. 2005. Familia Asteraceae. In: Flora Genérica de los Páramos: Guía Ilustrada de las Plantas Vasculares. P. Sklenár, J. L. Luteyn, C. Ulloa-U, P. M. Jørgensen, & M. O. Dillon (Eds), Memoirs New York Botanical Garden, 92: 96–180. [ Links ]
Dillon, M. O. & F. Luebert. 2015. Gnaphaliothamnus nesomii (Asteraceae, Gnaphalieae), A New Species from Guatemala and Nomenclatorial Changes in the Gnaphaliinae. J. Bot. Res. Inst. Texas, 9 (1): 63-73. [ Links ]
Dillon, M. O. & A. Sagástegui. 1991. Sinopsis de los géneros de Gnaphaliinae (Asteraceae-Inuleae) de Sudamerica. Arnaldoa, 1: 5–91. [ Links ]
Freire, S. E. 1984. Fenética del género Lucilia (Compositae: Inuleae). C.I.P.F.E. - C.E.D. Orione Cont. Biol. 11: 26-32, Montevideo. [ Links ]
Freire, S. E. 1986. Revision del género Lucilia (Compositae, Inuleae). Darwiniana, 27: 431-490. [ Links ]
Freire, S. E. 1987a. A cladistic analysis of Lucilia Cass. (Compositae, Inuleae). Cladistics, 3: 254–272. [ Links ]
Freire, S. E. 1987b. Nuevos sinonimos y una nueva combinación en el género Lucilia (Compositae, Inuleae). Darwiniana 28 (1-4): 409-411. [ Links ]
Freire, S. E.; M. A. Chemisquy; A. A. Anderberg; S. G. Beck; R. I. Meneses; B. Loeuille & E. Urtubey. 2014. The Lucilia group (Asteraceae, Gnaphalieae): Phylogenetic and taxonomic considerations based on molecular and morphological evidence. Pl. Syst. Evol. 301: 1227–1248. [ Links ]
Gray, A. 1862. Characters of some Compositae in the collection of the United States South Pacific Exploring Expedition under Captain Wilkes, with observations, &c. Proceedings of the American Academy of Arts and Sciences 5: 114–146. [ Links ]
Hieronymus, G. H. E. W. 1900. Compositae. Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 29: 1–85. [ Links ]
Luebert, F.; A. Moreira-Muñoz; K. Wilke & M. O. Dillon. 2017. Phylogeny and evolution of achenial trichomes in the Lucilia-group (Gnaphalieae, Asteraceae) and their systematic significance. Taxon 66(5): 1184-1199. [ Links ]
Merxmüller, H.; P. Leins & H. Roessler. 1977. Volume 1, Chapter 21. Inuleae - systematic review. Pp. 577-602. In: V.H. Heywood, J.B. Harborne, & B.L. Turner (eds.). The Biology and Chemistry of Compositae. Academic Press, London. [ Links ]
Nesom, G. L. 1990. Mexerion (Asteraceae: Inuleae), a new genus from Mexico. Phytologia, 68: 247–254. [ Links ]
Nie, Z. L.; V. Funk; H. Sun, H.; T. Deng; Y. Meng, & J. Wen. 2013. Molecular phylogeny of Anaphalis (Asteraceae, Gnaphalieae) with biogeographic implications in the Northern Hemisphere. J. Plant Res. 126: 17–32. [ Links ]
Philippi, R. A. 1862. Viaje a los baños i al nuevo volcán de Chillán. Anales Univ. Chile 21: 377-386. [ Links ]
Philippi, R. A. 1895. Plantas nuevas chilenas de las familias que corresponden al tomo IV de la obra de Gay: Anales Univ. Chile 90: 5-44. [ Links ]
Poeppig, E. F. 1845. Nova Genera Species plantarum, quas in regno chilensi peruviano et terra amazonica, 3: 1- 91. [ Links ]
Rémy, J. 1847. Compositae. In: Gay C, Historia Física y Política de Chile, Botánica 3: 257–482. [ Links ]
Sagástegui, A. & M. O. Dillon. 1985. New species and combinations in Belloa (Inuleae-Asteraceae). Phytologia, 58: 392- 400. [ Links ]
Schultz-Bipontinus, C. H. 1856. Lechler’s neueste Sammiungen aus Peru und Chile. Bonplandia. Zeitschrift für die gesammte Botanik. 4: 50-54.
Steyermark, J. 1953. Compositae. In: J. A. Steyermark & collaborators, Contributions to the Flora of Venezuela. Botanical Exploration in Venezuela—III. Fieldiana Botany 28 (3): 620–678. [ Links ]
Thiers, B. 2017. [continuamente actualizada] Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. Disponible en: http://sweetgum.nybg.org/ih/
Urtubey, E.; A. López; A. Chemisquy; A. A. Anderberg; C. M. Baeza; N. D. Bayón; L. P. Deble; A. Moreira-Muñoz; G. L. Nesom; M. H. Alford; L. Salomón & S. E. Freire. 2016. New circumscription of the genus Gamochaeta (Asteraceae, Gnaphalieae) inferred from nuclear and plastid DNA sequences. Plant Syst. Evol. 302: 1047-1066. [ Links ]
Ward, J.; R. Bayer; I. Breitwieser; R. Smissen; M. Galbany-Casals & M. Unwin. 2009. Gnaphalieae. Pp. 537–585 In: V. A. Funk, A. Susanna, T. F. Stuessy, & R. J. Bayer (Eds.), Systematics, evolution, and biogeography of Compositae. Vienna: International Association for Plant Taxonomy, University of Vienna. [ Links ]
Zardini, E. 1987. A new combination in Lucilia (Compositae-Inuleae). Ann. Missouri Bot. Gard. 74: 431. [ Links ]
Recibido 2-XII-2017; aceptado: 20-II-2018; publicado online: 15-III-2018; publicado impreso: 30-IV-2018

