










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Gastroenterología del Perú
versão impressa ISSN 1022-5129
Rev. gastroenterol. Perú vol.34 no.3 Lima jul. 2014
Reportes de Casos
Metagenomic analysis of the gastric microbiota cultivable from a patient with gastritis concomitant with Barretts esophagus
Análisis metagenómico de la microbiota gástrica cultivable a partir de un paciente con gastritis concomitante con esófago de Barrett
Andrés Julián Gutiérrez-Escobar 1,a, Martín Bayona-Rojas 1,b, Carlos Barragan-Vidal 1,c, Sebastián Rojas-Lara 1,d, Ricardo Oliveros 1,e
1 Grupo de Investigación en Ciencias Biomédicas y Genética Humana Aplicada GIBGA. Universidad de Ciencias Aplicadas y Ambientales. Bogotá, Colombia.
a Licenciado en Biología. Ms.c.; bBacteriólogo Ms.c.; cMicrobiólogo Ms.c.; dJoven investigador; eMédico Gastroenterólogo-cirujano.
ABSTRACT
Barretts esophagus is a distal metaplasia characterized by the transformation of squamous mucosa into columnar mucosa. This esophageal phenotype is a product not only of the chronic reflux of gastric acids, but also by microorganisms that colonize the oral cavity and stomach. Two classes of microbiota can be identified in Barretts esophagus; microbiota type I is associated with the normal esophagus and type II with an inflamed esophagus. The present study describes the gastric microbiota of a patient with antral gastritis concomitant with Barretts esophagus absent infection with Helicobacter pylori. Gastric biopsies were obtained following the protocol of Sydney and following ethical practices. The isolates were cultivated under microaerophilic conditions on Columbia Agar supplemented with IsoVitaleX and 7% sterile blood. Extracted DNA was sequenced using 454-GS and the results analyzed on the MG-RAST server. Gram negative isolates were found and bacteria resistant to levofloxacin, amoxicillin, tetracycline, erythromycin, and clarithromycin. The phyla Bacteroidetes, Firmicutes, Fusobacteria and Proteobacteria, the genus Bacteroides and the species group Bacteroides fragilis were most abundant. Functionally, the metabolism of carbohydrates, amino acids, and to a lesser extent, the metabolism of cofactors and vitamins were most dominant, and of which the enzymes β-glucosidase (EC 3.2.1.21), β-galactosidase (EC 3.2.1.23) and β-N-acetylhexosaminidase (EC 3.2.1.52) were most dominant. The findings of this study, because they are of only one case may probably suggest a possible pathogenic role, previously undescribed for Bacteroides fragilis, associated with human gastritis when concomitant esophageal pathology exists.
Key words: Barrett esophagus; Gastritis; Microbiota; Bacteroides fragilis (source: MeSH NLM).
RESUMEN
El esófago de Barrett es una metaplasia distal caracterizada por la transformación de la mucosa escamosa a mucosa columnar. Este fenotipo esofágico es producto no solo de la exposición crónica al reflujo de ácidos gástricos sino también a microbios colonizantes de la cavidad oral y del estómago. El esófago Barrett presenta 2 clases de microbiotas; la microbiota tipo I asociada con esófago normal y la tipo II a fenotipos esofágicos inflamatorios. En el presente estudio se describió la microbiota gástrica de una paciente con gastritis antral concomitante con esófago de Barrett sin infección por Helicobacter pylori y se obtuvieron biopsias gástricas siguiendo el protocolo de Sydney y estándares bioéticos. Los cultivos se hicieron en condiciones microaerofílicas en agar Columbia suplementados con isovitalex y sangre estéril al 7%. El ADN extraído fue sometido a secuenciación empleando 454 GS y las lecturas fueron analizadas en el servidor MG-RAST. Se obtuvieron aislamientos gram-negativos y resistentes a levofloxacina, amoxicilina, tetraciclina, eritromicina y claritromicina. Los Phylum Bacteroidetes, Firmicutes, Fusobacteria y Proteobacteria, el género Bacteroides y las especies de grupo Bacteroides fragilis fueron los más abundantes. Funcionalmente, el metabolismo de carbohidratos, aminoácidos y en menor grado el metabolismo de cofactores y vitaminas fueron los más dominantes; de los cuales las enzimas la β-glicosidasa (EC 3.2.1.21), β-galactosidasa (EC 3.2.1.23) y la β-N-acetilhexosaminidasa (EC 3.2.1.52) fueron las más dominantes. Estos resultados, por ser de un solo caso, solo podrían sugerir un posible papel patogénico no descrito para Bacterioides fragilis asociado con gastritis humana cuando existe patología esofágica concomitante.
Palabras clave: Esófago Barrett; Gastritis; Microbiota; Bacteroides fragilis (fuente: DeCS BIREME).
INTRODUCTION
Barretts esophagus is a distal metaplasia characterized by the transformation of squamous mucosa into columnar mucosa. Some 10% of patients with gastroesophageal reflux develop Barretts esophagus and have an increased risk of developing esophageal adenocarcinoma. It is believed that only 1 in 20 patients with esophageal cancer, whose 5 year survival rate is about 5%, have been diagnosed with this condition (1). This inflammatory phenotype of the esophageal mucosa is a result of the chronic reflux of gastric acids and bile. During reflux, both the squamous epitheliums as well as the lamina propia are exposed to microorganisms that colonize the oral cavity and the stomach (2).
Initially attempts were made to link Helicobacter pylori with the development of Barretts esophagus; however these bacteria could not be shown to increase in numbers in mucosa with this characteristic (1). However it was found that the absence of H. pylori was associated with an increased risk to suffer Barretts esophagus (3). On the other hand studies of the gastric microbiota demonstrated that not only H. pylori has the capacity to colonize the stomach, but that other microbial species can accompany the reflux as well (4,5). The 16S ribosomal RNA profile of the distal esophagus with epithelium from Barretts patients identified two dominant microbiotas as type I and type II. Type I microbiota is associated with normal esophagus, while type II with inflammatory phenotypes. It is known that the type I microbiota is dominated by Gram-positive bacteria, principally of the phylum Firmicutes and type II principally by Gram-negatives from multiple phyla such asBacteroidetes, Proteobacteria, Fusobacteria and Spirochaetes. The genus most dominant of the esophageal microbiota is Streptococcus spp. (6).
Although studies based on 16S rRNA profiles have been extensively used to characterize the taxonomic diversity of the microbiota in different anatomical regions, such as the mouth (7,8), the esophagus (9), the stomach (10), the small intestine (11), the colon (12,13) and the vagina (14,15), these types of studies are being replaced by other techniques based on latest generation sequencing, not only for the reduced cost but also because it is possible to get better information, for example, metabolic profiles, identification of signaling pathways and identification of enzymes (16).
To date, neither the taxonomic nor functional ends have been characterized in microbiota derived from chronic gastritis concomitant with Barretts esophagus without infection with Helicobacter pylori. In the present study we describe the microbiota from 5 gastric biopsies from a patient with chronic erosive antral gastritis concomitant with Barretts esophagus without infection with Helicobacter pylori using high performance pyrosequencing. We isolated, principally, Gram-negatives resistant to levofloxacin, amoxicillin, tetracycline, erythromycin and clarithromycin. The phyla identified were Bacteroidetes, Firmicutes, Fusobacteria and Proteobacteria. The genus Bacteroides and the species group Bacteroides fragilis were the most abundant among the isolates. Functionally, the metabolism of carbohydrates, amino acids, and to a lesser extent, the metabolism of cofactors and vitamins were most dominant, and the three most abundant enzymes were β-glucosidase (EC 3.2.1.21), β-galactosidase (EC 3.2.1.23) and β-N-acetylhexosaminidase (EC 3.2.1.52), all of which have clinical implications and are important in biotechnology.
As a result of an ongoing project on genomics of Helicobacter pylori we identified a patient from whom we characterized the metagenome of the cultivable gastric microbiota. Endoscopy was carried out on the high digestive tract in consultation with Gastroenterology using Olympus Eys Exera CLE-145 Score, CV-145, and a GIF Olympus Tips V Endoscope y Olympus FB-21K forceps. The endoscopes and forceps were disinfected with 3% glutaraldehyde. Clinical history was collected and informed consent signed. Biopsies were taken following the protocol of Sidney from 2009 (17). Ten biopsies were taken from the patient of whom five gastric biopsies were destined for microbiological study. The biopsies were placed in thioglycollate broth and transported at 8 °C to the laboratory of Biología Molecular de la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A. The other five biopsies were sent to pathology. The patient was protected by confidentiality and agreed to participate in this study voluntarily, offering answers to a series of questions which permitted reconstruction of clinical history, and contributing results of histological examinations in addition to the donation of gastric biopsies.
The biopsies were sterilely macerated and seeded onto Columbia agar enriched with 7% sterile blood, 2% IsoVitaleX (Labyotek) and 0.01% antimycotic (Invitrogen). The cultures were incubated for 10 days in the Gaspak system (5-10% 02; 5-10% C02; 8090% N2, 95% humidity) (BD) at 37 °C. Antibiotic susceptibility was determined using the Kirby-Bauer technique. The isolates were subcultured in Brucella broth (without antibiotics) in similar conditions and a suspension was prepared with the growth in Brucella broth adjusted to tube number 3 on the McFarland scale (approximately 1X108 CFU/ml) seeded onto supplemented Muller-Hinton agar (Oxoid). The following sensidiscs were used: levofloxacin (5 ug), amoxicillin (25 ug), tetracycline (30 ug), erythromycin (15 ug) and clarithromycin (15 ug). The cultures were incubated 4-7 days in triplicate. The zones of clearing were interpreted according to commercial standards.
DNA extraction was performed using UltraClean® kit (MoBio®). Concentration and purity were determined spectrophotometrically (Qubit® Invitrogen®). The presence of Helicobacter pylori was detected by PCR amplification of the 16S rRNA gene using primers UDCA-F (5-TCGTGTCGTGAGATGTTGGG -3) and UDCA-R (5-TAACAAACCGATAATGGCGC-3). PCR was carried out in 20µl final volume with 0.5 µM of each primer, 0.07 U/µl Taq polymerase (Biolase®, Bioline®), 2 mM MgCl2, 250 µM of each dNTP, 1X buffer, 50100 ng of DNA total with the following program: 4 min to 94 °C of initial denaturation followed by 30 cycles of 30 seconds at 94 °C, 30 seconds at 55 °C, and 30 seconds at 72 °C, with a final round of 7 minutes at 72 °C using a MyCycler® thermocycler (BioRad). The final products were visualized ina 1.5 % agarose gel.
500 ng DNA was used for the construction of a genomic library using GS FLX Titanium General Library Preparation Kit (Roche Applied Science, USA). The sample was nebulized to generate fragments of 600 to 900 base pairs and end adaptors were attached and finally amplified using emulsion PCR. Sequencing was carried out on the 454 GS FLX Titanium platform (Macrogen, Korea). The derived sequences were assembled using the program GS de novo.
The metagenome was analyzed in two phases: Predominant families, genera and species were identified through comparison with the databases Greengenes, M5RN, RefSeq and SwissProt. Afterwards, the metabolic profile of the metagenome was identified by comparison against the databases SEED, orthologous groups (COG), orthologs KEEG (KO). All of the analyses were carried out on MG-RAST (18) with an e value of 1e5, a percentage of identity of 60. Finally, we used the key words Oral, esophagus, gastro intestinal, stomach, intestine, oropharyngeal, and human microbiota in the database M5NR. The results are presented as percentages from BLAST.
CLINICAL CASE
A 56 year old female patient presented on 6 December 2012 for gastroenterology consultation with a clinical picture characterized by burning abdominal, epigastric pain with an insidious onset and mesogastric radiation. The patient described a trigger for pain as fasting and some foods with more than one episode per week. Occasionally she took Omeprazole capsules (20 mg) with poor clinical response. She described symptoms of heartburn, bloating, and a sour taste in her mouth, moderate flatulence, nausea and hyperoxia. She denied bloody vomiting and vomiting, diarrhea, melena, hematochezia, or other symptoms, and had no difficulty breathing. This patient had been diagnosed with Barretts esophagus approximately 2 years before in Ibague, Colombia by EVDA and had not received a diagnosis, or treatment for, Helicobacter pylori. Her food was rich in protein, fats and carbohydrates and poor in fiber and vitamins. Her stools were pale yellow in color, with a frequency of daily or once every two days and a soft consistency, but she denies straining, any important family history, but reports a younger sister with gastritis.
Endoscopy of the high digestive tract (EHDT) revealed: 1. Acute erosive gastritis and chronic antral gastritis; reddened gastric mucosa characterized by petechiae and erosions with friable mucosa producing superficial lesions, with endoscopic images compatible with acute erosive gastritis. In addition enlargement and mucosal pallor at the level of the antral endoscopic images were compatible with chronic erosive gastritis. 2. Barretts esophagus was seen as Barretts epithelium in the esophagus 2 cm distal from the gastro-esophageal union, confirming the clinical diagnosis of the specialist. The histological diagnosis from the esophageal biopsy was gastric and intestinal metaplasia, and from the gastric biopsy was chronic superficial gastritis.
Microbial isolates and the metagenome
Bacilli, cocci and pleomorphs were identified in a mix of Gram positives and Gram negatives. The specific PCR analysis for Helicobacter pylori was negative. The results from the sequence analysis of the generic PCR identified at least six distinct species:Bacteroides sp. 1_1_30, Bacteroides uniformis, Fusobacterium necrophorum, Clostridium thermocellum, Bacteroides sp. 2_1_16 and Bacteroides sp. 1_1_30. Finally the results of the antibiotic susceptibility tests showed the isolate as resistant to the antibiotics used (Figure 1).

The metagenome yielded 217,596 reads (average 556 bases) for a total of 121,045,430. 6.2% did not pass quality control, and of those that passed, 1.8% was ribosomal genes and 92% non-ribosomal genes. The reads contained 46.7 ± 7% GC. The final population submitted to analysis resulted in 64,617,502 base pairs, with 204,098 reads and an average of 316 base pairs per read. The Greengenes database indicated that 42% corresponded to the phylum Bacteroidetes, 15.6% to Firmicutes, 5.6% to Proteobacteria, and Actinobacteria at 1%. The M5NR data base reported, with respect to non-ribosomal genes, that at the level of Phylum 71% corresponded to Bacteroidetes, 15.6% to Firmicutes, and 5.6% % to Proteobacteria; at the level of class 70.6% Bacteriodia, some 8% Clostridia and some 7% to Bacilli, at the level of order 70.5% Bacteriodales, 7.6% Clostridiales and 5.4% Actinomycetales; at the level of family 64.%% corresponded toBacteroidaceae, and finally at the level of genus, Bacteroides was present at some 63.4%. The dominant species were Bacteroides fragilis, Bacteroides uniformis, and Bacteroides helcogenes (Figure 2).

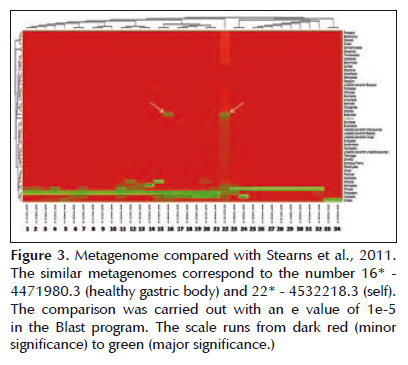
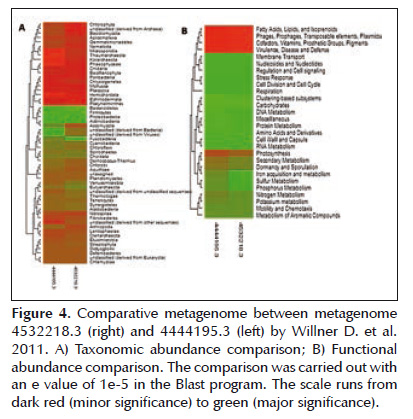
The metagenome appeared simultaneously with the project entitled Bacterial biogeography of the human digestive tract (19) that can be found at http:// metagenomics.anl.gov/metagenomics.cgi?page= MetagenomeProject&project=1982 . It presents 46 metagenomes covering the microbiota of the mouth, stomach, duodenum, colon and feces. On a general level all of the metagenomes are similar for Phylum Actinobacteria, Proteobacteria, Firmicutes and Fusobacteria. But both the metagenome of the gastric body like the present study are similar with respect to Bacteroidetes, in fact present in these two metagenomes. However the metagenome presented here has a unique taxonomic distribution pattern, for example reporting Euryarchaeota, Synergiestes and Verrucomicrobia for the first time in this type of human sample (Figure 3). Another homologous project was Human Oropharyngeal Metagenome (20). This project has two metagenomes and its objective was to identify species of bacteriophages in the human oral cavity. The project can be found at http://metagenomics.anl. gov/linkin.cgi?project=32 . The metagenome studied demonstrated a distribution very similar with respect to the dominant Phylum once again for Bacteroidetes. (Figure 4A).
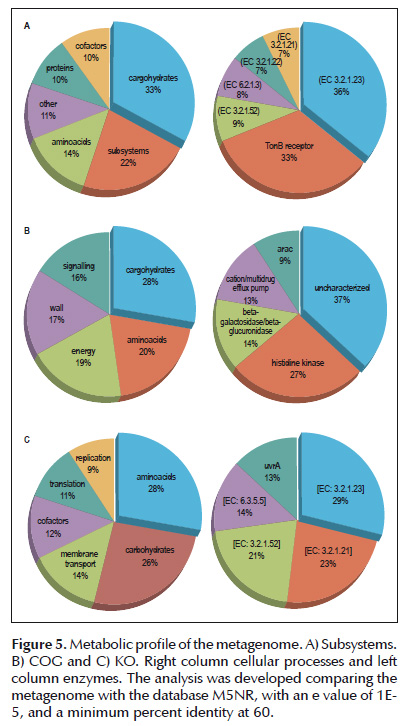
Metabolic mapping identified dominance towards carbohydrate and amino acid metabolism and in lesser proportion for cofactors and vitamins. Functional abundance highlights the presence of the enzymes β-glucosidase (EC 3.2.1.21), β-galactosidase (EC 3.2.1.23) and β-N-acetylhexosaminidase (EC 3.2.1.52) for all of the data bases. Specifically, subsystem classification identified a marked dominance of the protein dependent receptor TonB and the protein Glutarate-CoA ligase (EC 6.2.1.6); for COG the most abundant enzyme is the environmentally stimulated histidine kinase and the cationic efflux pump of antibiotics among other proteins of carbohydrate metabolism. Finally in KO the enzyme carbamoylphosphate synthase (EC 6.3.5.5) is the most abundant of enzymes among the DNA repair pathways. (Figure 5).
The functional comparison with the metagenome code 4444195.3 of the project Human Oropharyngeal Metagenome (20) permitted identification of great metabolic similarity covering processes from transport to DNA repair (Figure 4B).
DISCUSSION
The healthy oral cavity, esophagus and stomach contain microbiotas in equilibrium (21) but when this is lost, the loss is associated with illness. For example, the type I microbiota of the healthy esophagus and stomach contain similar taxonomies, however curiously, Firmicutes and the genus Streptococcus – dominant forms of the type I microbiota – are the most abundant in antral gastritis without infection with H. pylori (2,9,6,15).
As a result, one might suggest that the type II microbiota and the one described here could be associated: Bacteroidetes and Firmicutes being the most abundant, although it is not unusual to find them together (19), if it is significant that Firmicutes is present due to its linkage with gastritis. As the information on public databases has increased, Bacteroidetes, Firmicutes, Proteobacteria and Actinobacteria have been found as dominant phyla in environments as varied as baths (22) and fruits (23). A taxonomic linkage could be established between the gastric pathology with the external microbial ecosystem when Helicobacter pylori is not present.
Although taxonomic similarity was identified with the healthy gastric body and the oropharyngeal cavity (19,20) consequent with the above, as well other Phyla were identified such as Verrucomicrobia (24) that has been uniquely described in this metagenome. In fact a taxonomic difference is the dominance of the phylum Bacteroidetes, the genus Bacteroides and the species group Bacteroides fragilis that has been reported for the first time in the cultivable microbiota of isolates derived from human gastritis concomitant with Barretts esophagus. Normally, B. fragilis maintains a beneficial relationship with the host (25-28) although it is considered a principle anaerobic pathogenic bacterium (29-31). Initially this bacterium was only isolated within feces (32-34) but actually it can be isolated in diverse niches (35-40). Although the object of the current study was not to determine a cause-effect relationship between Bacteroides fragilis and antral gastritis concomitant with Barretts esophagus, there is published evidence that Bacteroides spp. could generate antral gastritis in an animal model (41). More study however is necessary. It should be noted that the isolates identified here are multiresistant to antibiotics that are recommended in practice, for example against Helicobacter pylori (Figure 1). This is, at least, significant for public health purposes in Colombia for Barret´s disease (42).
Functionally, dominance was seen for the metabolism of carbohydrates, amino acids, cofactors and vitamins. Although in the metagenomic study of the oropharynx the homologous reads of prokaryotes and eukaryotes were eliminated, we kept the reads that permitted complete comparison of the metagenomes. The functional similarity covered the synthesis of macromolecules, degradation and production of energy, cell wall, membrane transport, and the metabolism of ions. This demonstrated a dynamic functional activity that implied that similarity not only occurred at the taxonomic level, but also at the functional level, and that could reflect response to the surrounding ecosystem. Unfortunately, advanced studies concerning the biogeography of human microbiota are limited only to ribosomal profiles and it is not currently possible to realize a functional comparison. The most abundant enzymes are β-glucosidase [EC 3.2.1.21], important for the liberation of units of glucose from complete polysaccharides and as well in the development of biotechnological processes (43). The presence of this enzyme could indicate the existence of metabolic pathways for sucrose and the biosynthesis of tyrosine, phenylalanine and tryptophan. Another is β-galactosidase [EC 3.2.1.23]; important for the liberation of monosaccharides from lactose and as well could indicate pathways related to the synthesis of amino sugars and nucleotides, of glycine, serine and threonine. Finally, the enzyme β-N-acetylhexosaminidase [EC 3.2.1.52] participates as well in the synthesis pathway of amino sugars and nucleotides.
Subsystem classification identified two other enzymes. The first was the receptor TonB. In Escherichia coli the TonB protein interacts with its receptor assisting energy dependent uptake (44). The other enzyme is Glutarate-CoA ligase [EC 6.2.1.6] that indicates pathways for the degradation of fatty acids. On the other hand, COG identified a cationic efflux pump with code COG0841, possibly AcrB associated with resistance to antibiotics and another with code COG4585 corresponding with a histidine kinase implicated in signal transduction. Finally, the database KO identified the enzyme carbamoylphosphate synthase [EC 6.3.5.5] that indicates that the metagenome could present metabolic pathways of pyrimidine nucleotides, alanine, aspartate and glutamate.
Acknowledgements: We thank Diana Castiblanco and Julián Pineda for the contribution in transporting the samples.

