











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
En Ecuador existe una vasta población de bovinos de razas con aptitud lechera que se han establecido bajo diversas modalidades de sistemas de producción (intensivo, semiintensivo o al sogueo). Esto ha conducido a que las razas de bovinos nativos del altiplano del sur de Ecuador, cuya población se ha reducido considerablemente hasta el riego de extinción, se haya establecido como un sistema tradicional de crianza de la población indígena rural (Aguirre et al., 2011). En este sentido, Aguirre et al. (2014) hicieron una caracterización genética de los bovinos criollos de la región sur del Ecuador, determinando que tienen un gran distanciamiento con las razas cebuinas, y que sus troncos ancestrales están mayormente vinculados con las poblaciones ibéricas.
La conservación de estos grupos de bovinos autóctonos es de suma importancia para preservar esta fuente de biodiversidad genética, lo cual pudiera lograse mediante la aplicación de biotecnologías reproductivas. La literatura indica que no existen programas reproductivos que involucren la conservación de gametos (espermatozoides y ovocitos) o embriones provenientes de grupos bovinos nativos en Ecuador. En el Laboratorio de Biotecnología de la Reproducción Animal de la Universidad de Cuenca se han desarrollado protocolos de criopreservación de gametos y embriones de bovinos criollos con el fin de establecer bancos de germoplasma (Alvarado et al., 2018; Argudo et al., 2019; Méndez et al., 2019), lo cual constituye un punto de partida para la preservación de este valioso patrimonio genético.
Existen dos procedimientos para la recolección de ovocitos bovinos, uno post mortem, de ovarios de matadero, y otro in vivo, mediante la aspiración folicular guiada por ultrasonografía (ovum pick-up, OPU). El primero ha sido usado ampliamente como fuente de ovocitos para optimizar las diferentes etapas del proceso de producción in vitro de embriones (Manjunatha et al., 2008; Alvares et al., 2016), así como modelo experimental para estudiar aspectos biológicos relacionados con la producción in vitro de embriones en mamíferos (Nagai et al., 2001). El segundo ha sido aplicado para obtener material genético de animales superiores con el fin de mejorar la productividad de las operaciones comerciales bovinas (Bizarro et al., 2016). Esta técnica tiene un amplio campo de aplicación para colectar germoplasma de especies silvestres y de grupos genéticos en riesgo de extinción (Li et al., 2007; Berlinguer et al., 2008), incluyendo el bovino criollo latinoamericano en permanente estado de amenaza.
En consecuencia, y al contar con un número reducido de bovinos criollos, la técnica de aspiración in vivo de ovocitos podría ser una práctica biotecnológica para conservar el material genético e incrementar el número de animales. Sin embargo, procedimientos básicos sobre la calidad, tamaño y funcionalidad (que incluye integridad de la membrana plasmática y proceso de maduración) de los ovocitos requiere ser investigado previo a los procedimientos de fertilización in vitro (FIV) y producción de embriones. Debido a esto, el estudio tuvo por objetivo evaluar las características morfométricas y funcionales de ovocitos de vacas criollas ecuatorianas y compararlos con ovocitos obtenidos de ovarios de matadero.
MATERIALES Y MÉTODOS
Localización
La investigación se realizó en el laboratorio de Biotecnologías Reproductivas de la Facultad de Ciencias Agropecuarias, Universidad de Cuenca, ubicado en la Granja Irquis, parroquia Victoria del Portete en la provincia del Azuay, a una altura de 2648 msnm.
Fuente de Ovocitos
Diez vaquillas (24-36 meses) con rasgos morfológicos y fanerópticos de animales nativos de las alturas de los Andes ecuatorianos (>3000 msnm), no lactantes y clínicamente sanas fueron sometidas a repetidas aspiraciones foliculares por un periodo de tres meses. Las vaquillas fueron alojadas en la granja bajo pastoreo rotacional [Ryegrass (Lolium perenne) y kikuyo (Pennisetum clandestinum)] y sanidad controlada. La manipulación de los animales y la aplicación de los procedimientos biotecnológicos fueron desarrollados de acuerdo con las normas de bioseguridad y bienestar animal.
El procedimiento de OPU fue realizado una vez por semana en las instalaciones de la granja (10 sesiones con un total 100 aspiraciones), y la búsqueda de ovocitos se hizo inmediatamente después de la aspiración folicular. Asimismo, y con fines comparativos, se colectaron ovarios de vacas de un matadero local una vez por semana, coincidiendo con el procedimiento de OPU. El tiempo entre el sacrificio de los animales y la llegada de los ovarios al laboratorio en envases térmicos con solución fisiológica a 30 ºC fue en un máximo de tres horas. Las vacas de matadero pertenecían a un grupo aleatorio de vacas multíparas lecheras (Holstein, Pardo Suizo, mestizas y criollas) no gestantes, sacrificadas por descarte.
Se llegó a recolectar 1157 complejos cúmulo-ovocito (CCOs) bovinos. De estos 217 del grupo OPU y 886 del grupo de matadero mediante recuperación de ovarios post mortem (PM).
Aspiración Folicular Transvaginal
La aspiración folicular transvaginal por OPU fue realizada previa sedación de las vacas criollas con xilacina al 2% (0.02 mg/kg). Las vacas fueron inmovilizadas en un brete, vaciado el contenido rectal, y aplicada una dosis de anestésico epidural con Bupivacaina al 0.5% (0.3 mg/kg). El procedimiento de aspiración se llevó a cabo con un ecógrafo Aloka (Prosound 2, Tokio, Japón) provisto de un transductor convexo de 5 MHz acoplado a una guía de aspiración. Este dispositivo estaba conformado por una aguja de 18G x 75 mm conectada a una tubería plástica de 1 mm de diámetro y 2 m de longitud, y a una bomba de vacío (WTA BV 003D ®, Cravinhos, Brasil) regulada a 90 mmHg, que condujo el líquido folicular por succión a un tubo estéril de 50 ml.
Los CCOs fueron colectados en una solución buffer fosfato salino (PBS, Sigma, P-4417) a 38.5 °C, suplementada con 1% de suero fetal bovino y 10 UI/ml de heparina. El líquido colectado fue llevado al laboratorio, filtrado y lavado con PBS atemperado a 38.5 °C. El líquido filtrado fue vertido en una caja de búsqueda de 95 mm, y los CCOs fueron identificados bajo un estereoscopio (Nikon SMZ 745T, Tokio, Japón). La selección de los CCOs se hizo al lavar las estructuras tres veces en fluido oviductal sintético buferado con Hepes (hSOF). Luego, los CCOs fueron categorizados en calidad A, B o C según los criterios descritos por Hawk y Wall (1994), quienes consideraron características del citoplasma, zona pelúcida y células del cúmulo.
Aspiración Folicular de Ovarios de Matadero
Los ovarios obtenidos en el matadero fueron lavados sucesivamente en solución fisiológica estéril temperada a 37 ºC, y el tejido circundante fue removido. Se aspiraron los folículos con diámetros entre 2 y 8 mm con el mismo equipo, la misma presión de vacío y línea de aspiración que la usada con el procedimiento OPU. Asimismo, el lavado, filtrado, búsqueda y clasificación de los COCs fue similar al efectuado con el procedimiento OPU.
Evaluación de los Ovocitos
Actividad de la glucosa-6-fosfato deshidrogenasa (G6PD)
Un grupo de CCOs recuperados post mortem de ovarios de matadero (n=464) y mediante OPU (n=137), clasificados según su calidad, fueron incubados con azul brillante de cresilo (BCB, 26 µM) diluido en hSOF por 90 minutos, en una atmósfera a 38.5 °C, 90% de humedad y 5% de CO . Luego de lavarse tres veces en hSOF, los COCs fueron diferenciados en dos grupos dependiendo si el citoplasma estaba teñido (BCB+) o no (BCB-) de color azul, registrándose el número de ovocitos por categoría. La G6PD es una enzima que está presente durante el periodo de crecimiento ovocitario, pero cuya actividad desaparece o se reduce sustancialmente cuando los ovocitos culminan la fase de crecimiento. Así, debido a la ausencia de la enzima, los ovocitos que han terminado el crecimiento conservan la coloración azul en el citoplasma, mientras que, en los no teñidos, la actividad enzimática degrada el colorante.
Integridad de la membrana plasmática del ovocito
Otro grupo de CCOs obtenidos PM (n=422) y por OPU (n=134) fueron incubados en una solución de PBS al 0.15% del colorante azul tripán durante 10 minutos. Inmediatamente, los ovocitos fueron lavados tres veces en PBS y se observaron en un microscopio óptico para determinar si el citoplasma estaba teñido o no de color azul, y se registró el número de cada grupo. Esta tinción es usada para determinar el estado de la membrana plasmática del ovocito o viabilidad celular por exclusión de captación, debido a que la selectividad de la membrana plasmática de las células vivas impide la penetración del colorante, mientras que en las muertas ocurre libremente.
Morfometría Ovocitaria
Los ovocitos usados para la determinación de la actividad de la enzima G6PD y la viabilidad ovocitaria fueron incubados por un minuto en una solución de hialuronidasa en hSOF (1 mg/ml; p/v) y separados de las células del cúmulo por pipeteo sucesivo. Luego de lavados tres veces en hSOF, los ovocitos fueron fotografiados con una cámara digital (Excelis AU-600-HD; USA) acoplada a un microscopio de contraste de fase (Olympus CX31, Japón) y el diámetro de cada uno (excluyendo la zona pelúcida) medido con el software CaptaVision (v. 5.1, Accu-Scope, USA) diseñado para realizar mediciones microscópicas.
Análisis Estadístico
Las proporciones de ovocitos fueron analizadas mediante la regresión logística del programa estadístico SAS (Statistical Analysis System, 2012; v. 9.4), considerándose los efectos de la fuente de ovocitos (OPU y matadero) y de la repetición (n=10). Dado que los datos del diámetro y volumen ovocitario (estimado con la fórmula del volumen de una esfera) no siguieron una distribución normal según la prueba de Shapiro-Wilk, estos fueron transformados al Logaritmo 10 y procesados mediante el análisis de varianza de una vía, usándose el modelo lineal general (GLM) del SAS (p<.0.05). En el modelo se consideraron los efectos de la fuente, calidad ovocitaria y la repetición. Las diferencias entre las medias fueron comparadas mediante el método de los cuadrados mínimos del SAS.
RESULTADOS Y DISCUSIÓN
Los resultados mostraron que los ovocitos del grupo PM mostraron un mayor diámetro y volumen (p<0.01), así como un mayor porcentaje de integridad de la membrana plasmática (p<0.05) en comparación con los ovocitos del grupo OPU. Por otro lado, no hubo diferencias significativas entre grupos con relación a la actividad de la enzima G6PD (Cuadro 1).
Cuadro 1 Características morfométricas y funcionales de ovocitos de acuerdo con la fuente ovocitaria (1157 ovocitos analizados en 10 repeticiones)
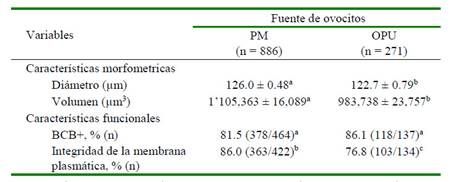
PM, grupo de ovocitos recuperados post mortem; OPU, grupo de ovocitos recuperados in vivo por aspiración folicular. Letras diferentes dentro de filas son significativamente diferentes (a,b p<0.01; b,c p<0.05)
Al categorizar los ovocitos según su calidad, los resultados mostraron que los ovocitos de calidad A (p<0.01) y B (p<0.02) del grupo PM tuvieron mayor tamaño que los del grupo OPU. Asimismo, hubo una mayor proporción de ovocitos de calidad A (p<0.05) y menor proporción de calidad C (p<0.01) en el grupo PM que en los de OPU (Cuadro 2). Sin embargo, no hubo diferencias significativas entre grupos cuando fue comparado el porcentaje de integridad de la membrana plasmática y el estado de maduración (GDP) en los ovocitos de calidades A y B; no obstante hubo diferencias significativas para los ovocitos de calidad C (Figura 1).
Cuadro 2 Diámetro (µm) y frecuencia (n) de ovocitos obtenidos de ovarios de matadero o in vivo mediante la técnica de ovum pick-up, según su origen y calidad

Letras diferentes dentro de filas son significativamente diferentes (a,b p<0.01; b,c p<0.02; c,dp<0.05)
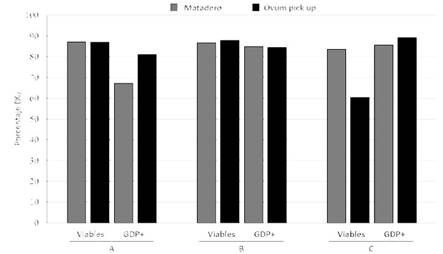
Figura 1 Porcentaje de ovocitos viables y negativos a la actividad de la enzima G6PD (BCB+) de acuerdo con la fuente y calidad ovocitaria
Los resultados obtenidos demostraron que los ovocitos recuperados in vivo mediante OPU mostraron un tamaño y un porcentaje de integridad de la membrana plasmática menor que los ovocitos recuperados post mortem, aunque el estatus metabólico para proseguir la maduración (BCB+) fue similar entre ambos grupos. Asimismo, la calidad, viabilidad y actividad de la enzima G6PD fue estadísticamente similar en los ovocitos A y B entre ambas fuentes. Estas dos categorías de ovocitos son las más apropiadas para la maduración y producción de embriones in vitro (Ward et al., 2000) y son las que utilizan rutinariamente en los laboratorios comerciales y de investigación.
La diferencia en las medidas morfométricas (diámetro y volumen) entre ambos grupos pudiera atribuirse al componente racial; es decir, al hecho de que se trabajó con ovocitos colectados de vacas nativas de la sierra alta de Ecuador, por un lado, y de ovarios post mortem provenientes de diferentes grupos raciales lecheros y sus mestizos. No obstante, no hay datos publicados del efecto racial sobre las características morfométricas de los ovocitos que permita darle soporte científico a este racionamiento; sin embargo, como se indica más adelante, el análisis de varios trabajos en los que se hizo medición de ovocitos de diferentes grupos raciales permite deducir que el diámetro ovocitario no varía considerablemente entre ellos.
La calidad de los CCOs recolectados por OPU y su competencia para madurar in vitro se vió afectada por la presión de aspiración folicular ejercida durante la punción (Bols et al., 1996; Fry et al., 1997; Vos et al., 1999). La técnica OPU realizada en vacas y búfalas a una presión de 40 mm Hg ha demostrado proporcionar una calidad similar a la técnica de recuperación de ovocitos de matadero post mortem (Neglia et al., 2003).
Reportes previos han demostrado que la calidad de los ovocitos y la competencia para desarrollarse como embriones es considerablemente afectada cuando la aspiración folicular se efectúa a presiones superiores a 50 mmHg (Ward et al., 2000).
Argudo et al. (2017, 2018) analizaron el tamaño de 489 y 782 ovocitos, respectivamente, recuperados de ovarios de matadero de la misma localidad (Sierra ecuatoriana) y obtuvieron un diámetro ovocitario de 124.1 ± 0.3 µm. Ese estudio demostró que el diámetro disminuye según se incrementa la presión de aspiración folicular (por ejemplo, 125.8 ± 0.5, 123.3 ± 0.5 y 122.2 ± 0.5 µm para ovocitos aspirados con 50, 65 y 80 mmHg, respectivamente). Los resultados de la presente investigación demostraron que los ovocitos aspirados por OPU se vieron afectados al usar una presión de aspiración alta (90 mmHg).
Hay que tener en cuenta que durante la OPU, los COCs están sometidos a fuerzas físicas que, dependiendo de la presión de succión y del diámetro de la aguja utilizados, afectan en mayor o menor cuantía la morfología de los COCs y la competencia de estos para desarrollarse a embriones (Bols, 1997). Aparentemente, durante la punción folicular, el líquido folicular con los ovocitos sufre turbulencias que provoca choques repetidos de estos con las paredes internas de la aguja y del extenso tubo que la conecta con el recipiente de colección (línea de aspiración), haciendo que el ovocito pierda progresivamente las células del cúmulo que lo rodean (Bols et al., 1996; Bols, 1997; Hasler, 1998; Bizarro et al., 2016), exponiendo la zona pelúcida y la membrana plasmática del ovocito a daños irreversibles.
Aunque la mención anterior no puede aplicarse totalmente a este estudio, dado que se utilizó la misma bomba y presión de vacío, y longitud de la aguja y de la línea de aspiración para obtener los ovocitos de ambas fuentes, la recuperación de COCs in vivo es más engorrosa y requiere que los animales sean tranquilizados, bloqueados epiduralmente e inmovilizados.
Los ovocitos bovinos se vuelven meióticamente competentes con un diámetro de aproximadamente 110 mm (Fair et al., 1995). Además, la variación, tanto en la capacidad meiótica de los ovocitos (capacidad para alcanzar la metafase II), como en la capacidad de desarrollarse a blastocistos, se ha evidenciado más en ovocitos mayores a 110 mm de diámetro (Fair et al., 1996). Por lo tanto, los resultados obtenidos en este estudio sugieren que, independientemente si la recuperación fue in vivo o post mortem, los ovocitos tenían un diámetro adecuado para completar la meiosis I e iniciar la II.
Reportes anteriores han investigado el tamaño folicular y de los ovocitos obtenidos de ovarios de matadero, probablemente de vacas Bos taurus. Entre estos, Fair et al. (1995) reportaron diámetros de ovocitos que variaron entre 98.9 ± 11.7 y 117.2 ± 9.7 µm que fueron colectados de folículos que median entre <1 y >4 mm de diámetro. Otoi et al. (1997) reportaron un diámetro promedio de 114.0 ± 4.8 µm, encontrando que el 82% midió entre 110 y 125 µm. En el caso de vacas Bos indicus, Nunes et al. (2000) reportaron tamaños entre 154.7 ± 11.6 µm en folículos pequeños (1-2 mm) y 164.4 ± 9.1 µm en folículos grandes (>9 mm), incluyendo la zona pelúcida. En el presente estudio, el diámetro de los ovocitos estuvo dentro de los rangos reportados en la literatura científica.
CONCLUSIONES
Los CCOs recuperados por OPU fueron más pequeños y tuvieron una viabilidad menor que los obtenidos post mortem, aunque ambos grupos presentaron similar estado metabólico para proseguir la maduración.
Los COCs de tipo A y B obtenidos de matadero o mediante OPU fueron estadísticamente similares en integridad de membrana y en estado metabólico, aunque los recuperados mediante OPU fueron cerca de 5 µm más pequeños que los obtenidos de matadero.
Los COCs de categoría C recuperados mediante OPU tuvieron considerablemente menor viabilidad que los tipos A y B del mismo grupo y los C del grupo post mortem. Asimismo, los COCs de calidad C tuvieron similar diámetro en ambas fuentes ovocitarias

