











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La Leucosis Bovina Enzoótica es una enfermedad infecciosa de distribución mundial con mayor incidencia en las explotaciones lecheras debido, en gran medida, al hacinamiento y las diferentes prácticas veterinarias que facilitan la transmisión. Es causada por el Virus de la Leucosis Bovina (BLV), perteneciente a la familia Retroviridae.
El BLV es un virus de RNA cadena simple diploide, que contiene la información genética para las proteínas estructurales y enzimas en los genes gag, pol y env (Vogt, 1997). El BLV codifica, además, proteínas no estructurales en una región denominada pX, que se transcriben a través de splicing alternativo: Tax, Rex, y en menos abundancia R3 y G4 (Alexandersen et al., 1993). Estas proteínas no estructurales tienen fundamental importancia en la interacción del virus con la célula hospedadora, modulando la expresión de genes virales y celulares, la replicación viral y la patogénesis (Willems et al., 2000).
Las prevalencias de infección por BLV son altas en la mayoría de los países productores de leche (Figura 1), excepto en países de la Comunidad Económica Europea, los que, desde 1980, han implementado medidas de control y erradicación del virus. Las tasas de prevalencia en el continente americano son muy altas, tanto a nivel individual como a nivel de rodeo (Figura 2). La diseminación de la infección por BLV en estos países posiblemente sea consecuencia de que, durante muchos años, se subestimaran los efectos de la infección sin la implementación de las políticas sanitarias necesarias.
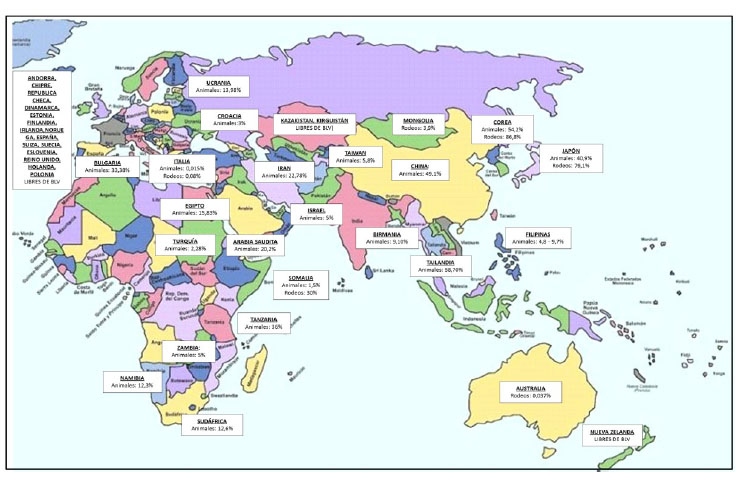
Figura 1 Prevalencias de infección por el virus de la leucosis bovina (BLV) en países de Europa, Asia, África y Oceanía. Se muestran datos de prevalencia a nivel individual (animales) y de rodeo
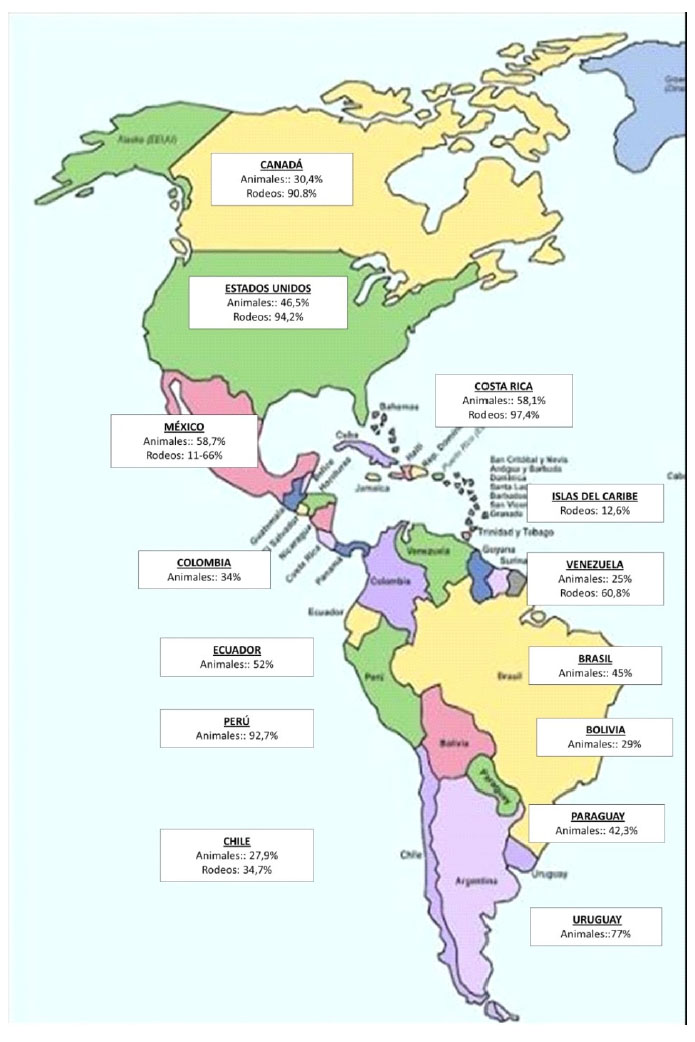
Figura 2 Prevalencias de infección por el virus de la leucosis bovina (BLV) a nivel individual (animales) y a nivel de rodeos infectados en el continente americano
En Argentina, desde hace más de 30 años se observa una amplia diseminación del virus, sin que hasta el momento se haya implementado un plan oficial de control/erradicación a nivel nacional. En consecuencia, esta enfermedad es una de las principales causas de muerte y descarte entre las vacas de producción láctea. Los primeros relevamientos serológicos de la situación en Argentina se realizaron en la cuenca Mar y Sierras entre 1979 y 1981, encontrando solo un 4.35% de rodeos infectados. Quince años después, este porcentaje llegaba al 68.5% (Ghezzi et al., 1997). Esta rápida diseminación del BLV también se observó en el resto de las cuencas lecheras argentinas. Los relevamientos serológicos realizados entre 2001 y 2004 en diferentes zonas del país mostraron valores similares (84-86%) de rodeos infectados para la provincia de Santa Fé (Trono et al., 2001; Mariño et al., 2003) y 97% para el sudeste de la provincia de Santiago del Estero (Revelli et al., 2004).
Datos más actuales de las cuencas lecheras de las provincias de Córdoba y Santa Fé, norte de la provincia de Buenos Aires y Sudoeste de la provincia de Santiago del Estero indican que más del 99% de los tambos están infectados (Barrios et al., 2012). Todas las zonas estudiadas muestran una alta tasa de diseminación de la infección dentro de los establecimientos, con valores entre 30 y 48.9% en el periodo 2001-2004 (Trono et al., 2001; Mariño et al., 2003; Revelli et al., 2004). Datos más recientes de establecimientos en estas provincias obtenidos por el Centro de Investigación Veterinaria de Tandil (CIVETAN) muestran que, en promedio, más del 80% de los animales en producción están infectados. En la categoría vaquillonas, la prevalencia de infección varía entre 15 y 75% (datos de los autores, no publicados), evidenciando una disminución de la edad a la cual los animales se infectan y el consecuente agravamiento de la situación.
La infección por BLV genera importantes pérdidas económicas. Las pérdidas directas incluyen: muerte de animales en producción por leucemia o linfosarcoma, eliminación prematura de los animales y restricciones en el comercio internacional de bovinos y de material genético. Por otro lado, las pérdidas económicas indirectas (reemplazo del animal en producción, el diagnóstico y la atención del veterinario, y la pérdida de un ternero y de la producción de leche durante aproximadamente 10 meses) también son relevantes aunque difíciles de estimar (Rhodes et al., 2003). El efecto de la infección por el BLV en los parámetros productivos aún es controversial. Estudios previos no encontraron una asociación significativa entre la infección por BLV y la producción de leche (Jacobs et al., 1991; Kale et al., 2007; Tiwari et al. 2007; Sorge et al., 2011). Trabajos más recientes indican un efecto negativo en la producción y calidad de la leche debido a la infección. A nivel de rodeo se encontró una asociación lineal entre la mayor prevalencia de infección por el BLV y una menor producción de leche (Ott et al., 2003; Erskine et al., 2012). A nivel individual, se observó que la infección por BLV disminuye la producción de leche en 1.5%/vaca/año (Norby et al., 2016), estimándose una pérdida de 11 000 kg/animal en su vida productiva (Nekouei et al., 2016).
La infección por BLV también se ha asociado con un aumento en el recuento de células somáticas en leche, lo cual sucede especialmente en vacas con más de cuatro lactancias (Yang et al., 2016). El aumento en el recuento de células somáticas es un indicador de mastitis clínica y subclínica y, por ende, está asociado a una menor producción láctea, a leche con mayor carga bacteriana y menor porcentaje de grasa y caseína, a una menor producción y calidad de quesos, y a un menor tiempo de conservación de los productos lácteos. También se le ha asociado a un aumento del intervalo entre partos y mayor número de servicios por concepción, sugiriendo que el BLV podría tener influencia en el comportamiento reproductivo del ganado lechero (Vanleeuwen et al., 2010; Romero et al., 2015).
El BLV puede afectar las células del sistema inmune en muchos niveles, especialmente en la proliferación, apoptosis y expresión de receptores y citoquinas (Kabeya et al., 2001; Frie y Coussens, 2015; Iwan et al., 2017). Además, al afectar el crecimiento, la diferenciación y la sensibilidad de las células, podría regular la potencia y la duración de las reacciones inmunológicas (Kabeya et al., 2001; Blagitza et al., 2017; Brym et al., 2018). Se ha propuesto que la infección con este virus predispone o disminuye la resistencia a la infección contra otros patógenos. Frie et al. (2017) hallaron que las vacas infectadas con BLV presentan títulos significativamente más bajos de anticuerpos específicos de antígeno en comparación con las vacas no infectadas. Asimismo, se ha observado una respuesta serológica disminuida frente a la vacunación contra varios patógenos en animales positivos a BLV, sugiriendo una menor protección vacunal en las vacas infectadas (Erskine et al., 2011; Frie y Coussens, 2015; Frie et al., 2016; Puentes et al., 2016).
PATOLOGÍA INDUCIDA POR EL BLV EN SU HUÉSPED NATURAL
El blanco principal del BLV son los linfocitos B, aunque se sabe que el virus puede infectar in vivo e in vitro otras poblaciones celulares, como por ejemplo los macrófagos. La mayoría de las células B infectadas expresan el marcador CD5 (Depelchin et al., 1989; Meirom et al., 1993; Mirsky et al., 1993). Al estar infectado el linfocito B, el genoma de ARN es copiado a ADN por la enzima transcriptasa reversa codificada por el virus. Mediante infecciones experimentales, se sabe que el virus se replica rápida e intensamente a través del ciclo infeccioso, el cual involucra la producción de nuevos viriones, la infección de linfocitos, y la transcripción reversa e integración del genoma viral como provirus en las células infectadas. Este proceso conduce a un pico de carga proviral hacia las 4-8 semanas post-infección. Una vez que el animal infectado desarrolla la respuesta inmune específica, las células infectadas en esta etapa inicial son eliminadas masivamente por el sistema inmune, y la carga proviral desciende significativamente (Gillet et al., 2013). A partir de ese momento el virus comienza a propagarse a través de la expansión clonal de células infectadas, que, al duplicar su material genético replican también el genoma del BLV integrado como provirus, sin evidencia de transcripción reversa (Florins et al., 2007; Pomier et al., 2008).
La infección por BLV se ajusta al fenómeno conocido como "iceberg", típico de muchas infecciones virales (Figura 3). La infección es asintomática en la mayoría de los casos, mientras que en algunos animales desarrolla un incremento permanente y estable de linfocitos B en sangre periférica, conocido como linfocitosis persistente (LP) (Ferrer et al., 1978). La LP se considera una condición benigna que resulta de la acumulación de linfocitos B no transformados, debido a un desbalance entre la proliferación y la muerte celular (Debacq et al., 2003). La punta del iceberg está representada por 1-5% de los animales infectados que desarrollan la enfermedad tumoral (Ferrer et al., 1979).
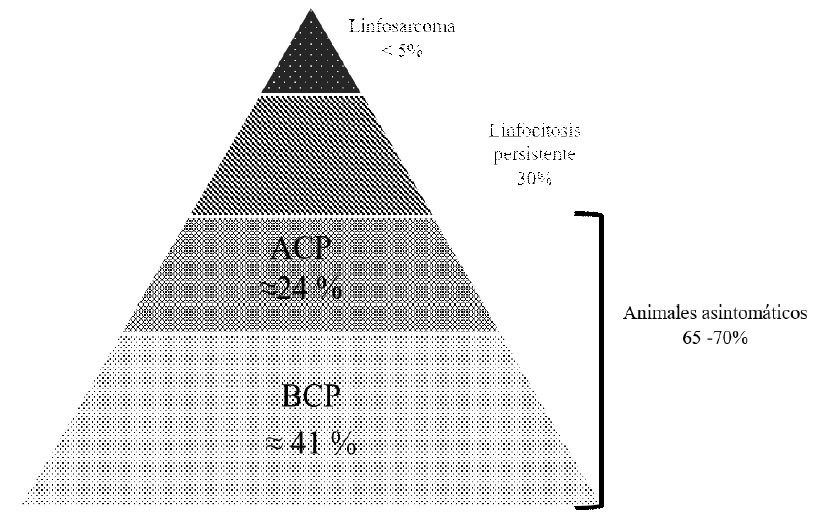
Figura 3 La infección por el virus de la leucosis bovina (BLV) se ajusta al fenómeno "iceberg".La mayoría de las infecciones por BLV son clínicamente inaparentes, con o sin alteraciones hematológicas (linfocitosis persistente). La infección inaparente se puede identificar por métodos serológicos (presencia de anticuerpos específicos) o moleculares (detección del provirus por PCR). Los casos clínicos de linfosarcoma se observan en menos del 5% de los animales infectados, y se identifican a través del examen clínico o como hallazgos de necropsia, y deben ser confirmados mediante métodos histopatológicos. ACP: alta carga proviral; BCP: baja carga proviral
La condición patológica (linfoma o linfosarcoma multicéntrico) resulta de la acumulación de linfocitos transformados por el virus en uno o más órganos, luego de un periodo de latencia prolongado que puede durar entre 1 y 8 años (Ferrer, 1980). Las lesiones se observan como masas tumorales firmes y blanquecinas, pero también puede presentarse como un infiltrado tisular difuso, manifestándose a la necropsia como un órgano de mayor tamaño y de color pálido, que puede ser confundido con un cambio degenerativo (Radostitis et al., 2007). El linfosarcoma puede comprometer diversos órganos, especialmente el abomaso, corazón, linfonodos viscerales y periféricos, bazo, útero y riñones. Esta condición es más frecuente en animales de 4-8 años, y muy raro en animales menores de 2 años (Ferrer, 1980).
Los signos clínicos dependen de la ubicación y ritmo de crecimiento de los tumores. En la mayoría de los casos, el curso de la enfermedad es subagudo a crónico y su inicio muchas veces coincide con el parto, comprometiendo la siguiente lactancia. Los signos más frecuentemente observados son: debilidad, pérdida de apetito y de peso, disminución en la producción de leche, linfadenopatía externa e interna y paresia posterior (Radostitis et al., 2007). El curso es hiperagudo en el 5-10% de los casos, donde los animales mueren sin signos clínicos previos. Esta presentación puede ocurrir si se afectan las glándulas adrenales, o a consecuencia de una hemorragia interna por la ruptura del bazo o de una úlcera abomasal perforada (Radostitis et al., 2007). La enfermedad es invariablemente fatal. La descripción detallada de las características clínicas y hallazgos de necropsia puede encontrarse en Radostitis et al. (2007).
Los bovinos infectados con BLV que no presentan signos clínicos ni hematológicos, comprenden al menos dos grupos que pueden ser diferenciados según el nivel de carga proviral (CP) en sangre periférica; es decir, la cantidad de provirus que portan, y el nivel de su respuesta inmune hacia el BLV (Juliarena et al., 2007, 2017a). Algunos animales se caracterizan por alcanzar altos niveles de CP (>100 000 copias de provirus/µg ADN), medida a través de la técnica de PCR, y una elevada respuesta de anticuerpos contra la proteína más antigénica del BLV, la glicoproteína de 51kDa de la envoltura viral (BLVgp51). Este grupo se denomina de "alta carga proviral" (ACP), y no se diferencia de los animales con LP en términos de CP o respuesta de anticuerpos hacia el virus. El segundo grupo abarca aproximadamente el 60% de los animales clínica y hematológicamente sanos, y comprende aquellos animales que presentan muy baja carga proviral en sangre periférica y bajo nivel de anticuerpos específicos contra los principales antígenos del BLV, y se les denomina animales de "baja carga proviral" (BCP). Los bovinos con BCP usualmente tienen menos de 100 copias del provirus/µg ADN, de allí que muchas veces no son detectados por los métodos moleculares habitualmente utilizados (PCR y PCR en tiempo real) (Juliarena et al., 2007).
El mecanismo de patogenicidad del BLV no está aun completamente dilucidado. Este virus no porta oncogenes, pero codifica proteínas no estructurales con potencial oncogénico como Tax y G4 (Willems et al., 1990; Kerkhofs et al., 1998). La integración del provirus en regiones cercanas a genes "impulsores o controladores de cáncer" (cancer drivers), altera la actividad de estos genes y constituye un mecanismo inicial de tumorigénesis (Rosewick et al., 2017). Utilizando tecnologías modernas como el secuenciamiento profundo de RNA (RNAseq) se han identificado transcriptos antisentido y microRNAs codificados por el BLV, los que jugarían un rol importante en el ciclo vital del virus, en la persistencia en el hospedador y su patogenicidad (Durkin et al., 2016; Gillet et al., 2016).
Los primeros estudios epidemiológicos mostraron que tanto el linfosarcoma como la LP se observaban más frecuentemente en determinadas líneas genéticas, y que ambas condiciones están bajo el control de factores genéticos del hospedador (Ferrer et al., 1978; Ferrer, 1980). La estimación de la heredabilidad sugiere que la genética también influencia la susceptibilidad a la infección por BLV (Burridge et al., 1979). Las bases moleculares de la resistencia y susceptibilidad genética al desarrollo de LP en el bovino han sido mapeadas a nivel de las moléculas de clase II del Complejo Mayor de Histocompatibilidad (CMH) bovino (BoLA), más específicamente en el gen DRB3 (Xu et al. 1993a; Zanotti et al. 1996). El CMH comprende un conjunto aproximado de 150 genes que se encuentra presente en todos los vertebrados y que interviene en los procesos de inmunidad (Janeway et al., 2001).
Específicamente, el gen DRB3 codifica las cadenas β de las moléculas de clase II del CMH (Gutiérrez et al., 2017). La función de las moléculas de clase II del CMH es presentar péptidos antigénicos a los linfocitos T durante la inducción de la respuesta inmune específica (Takeshima y Aida, 2006). Estos genes son altamente polimórficos (para el gen DRB3 se han descripto 136 alelos) (European Bioinformatics Institute, s.f.) y han sido asociados con el nivel de respuesta inmune a los antígenos y con la resistencia o susceptibilidad a varias enfermedades (incluyendo mastitis, leucosis bovina, parasitosis, entre otras) (Takeshima y Aida, 2006). Otro gen perteneciente al CMH, el gen del factor de necrosis tumoral α (TNF-α) también ha sido asociado con el BLV. Altos niveles de esta citoquina en la fase temprana de la infección podrían contribuir a la eliminación del BLV (Kabeya et al., 1999). Un polimorfismo en este gen, que afecta el nivel de expresión de TNF-α, ha sido asociado con el linfosarcoma inducido por BLV y ACP en bovinos infectados (Konnai et al., 2006).
Estudios del presente grupo de investigación han demostrado que los fenotipos de BCP y ACP también están fuertemente asociados al gen BoLA DRB3 en la raza Holstein, sugiriendo que la respuesta inmune celular es importante para el control de la diseminación del BLV (Juliarena et al., 2008). Estos resultados han sido confirmados posteriormente por otros autores, y ampliados a otras razas bovinas (Miyasaka et al., 2013; Carignano et al., 2017; Hayashi et al., 2017; Hernandez et al., 2018).
TRANSMISIÓN
Normalmente el BLV no se encuentra en el animal infectado como virus libre sino integrado como provirus al genoma del hospedador. Por lo tanto, los fluidos que contienen linfocitos infectados como sangre, leche u otra secreción pueden, al entrar en contacto con las membranas mucosas o tejidos lesionados, o al ser directamente inoculados, transmitir la infección viral a animales susceptibles (Hopkins y DiGiacomo, 1997). La principal vía de transmisión del BLV es horizontal, ya sea en forma natural o iatrogénica (Nekouei et al., 2015). Las intervenciones humanas asociadas a prácticas médico-veterinarias y de manejo como: descornados, castraciones, extracciones de sangre, palpaciones rectales y aplicaciones de caravanas pueden, si se realizan sin la correcta desinfección del instrumental utilizado, causar el contacto de sangre entre animales (Hopkins y DiGiacomo, 1997). Probablemente esta sea la principal causa de la amplia diseminación del virus en la mayoría de los rodeos argentinos.
El potencial de transmisión de un animal infectado depende del número de linfocitos infectados contenidos en la sangre periférica, por lo que los animales con fenotipo de ACP, especialmente con LP, son mejores transmisores del virus que los animales con fenotipo de BCP (Juliarena et al., 2007). La dosis mínima infectante proveniente de un animal con LP e inoculada en corderos es de 926 linfocitos (Mammerickx et al., 1987). El volumen de sangre estimado que contiene ese número de linfocitos infectados es menor a 0.5 µl para un animal con fenotipo de ACP, y mayor a 5 ml para un animal con fenotipo de BCP (Juliarena, 2008). En consecuencia, todos los bovinos que presentan el fenotipo de ACP son potenciales transmisores del BLV, mientras que los animales que presentan el fenotipo de BCP no transmiten la infección en las condiciones normales de manejo productivo (Juliarena et al., 2016). En condiciones experimentales es posible transmitir el BLV de un animal con perfil de BCP a corderos, lo cual demuestra que el BLV mantiene su poder infectante y su virulencia en los animales de BCP. Sin embargo, es necesario inyectar un volumen grande de sangre (entre 10 y 100 ml) para lograr la transmisión (Juliarena, 2008; Esteban et al., 2009; Juliarena et al., 2016).
Otra posible vía de transmisión horizontal del BLV es a través de insectos hematófagos, que pueden actuar como vectores mecánicos, especialmente en zonas de climas cálidos (Ohshima et al., 1981). Este tipo de transmisión depende de la densidad de insectos, de la proporción de animales infectados y del nivel de la carga proviral en los animales (Foil et al., 1988; Kohara et al., 2018). Esto explicaría que durante los meses de verano, coincidente con una mayor densidad de insectos hematófagos, se produce el contagio de la enfermedad en establecimientos donde se aplican medidas de higiene estrictas en las maniobras de manejo animal, tanto entre animales criados a campo como en estaca o en corrales contiguos (Kobayashi et al., 2014, 2015).
La transmisión vertical prenatal ocurre a tasas bajas (3-8%), principalmente por infección transplacentaria después de establecida la competencia inmunitaria (tercer mes de gestación) (Lassauzet et al., 1991; Hopkins y DiGiacomo, 1997) o durante el pasaje por el canal de parto. La LP y altos niveles de carga proviral en la madre son factores de riesgo para la transmisión prenatal del BLV (Lassauzet et al., 1991; Agresti et al., 1993; Mekata et al., 2014). La transmisión vertical postnatal en terneros alimentados con leche o calostro proveniente de madres infectadas es poco frecuente, probablemente debido a que los anticuerpos específicos obtenidos por transferencia pasiva natural les confieren protección (Ferrer y Piper, 1981). Sin embargo, los terneros nacidos de madres no infectadas son susceptibles a la infección si se los alimenta con leche de vacas infectadas, porque carecen de la protección que confieren los anticuerpos anti-BLV maternos (Romero et al., 1983). El BLV raramente se detecta en el semen de toros infectados (Choi et al., 2002; Dus Santos et al., 2007; Benitez et al., 2019b). La transmisión del BLV a través del semen no ha sido demostrada ni por medio de la inseminación artificial ni por servicio natural (Benitez et al., 2019a).
Recientemente, la preocupación sobre el potencial zoonótico del BLV ha reemergido, a raíz de publicaciones que sugieren una asociación entre el BLV y el cáncer de mama humano (Buehring et al., 2015, 2017; Schwingel et al., 2019). Un estudio reciente reporta la detección de secuencias del BLV y anticuerpos anti-BLV en un alto porcentaje de muestras de sangre humana, pero no se encuentra correlación entre la presencia del genoma y los anticuerpos específicos (Buehring et al., 2019). Sin embargo, la evidencia de esta asociación es controversial y la opinión de la comunidad científica internacional es escéptica acerca de la posible asociación entre el BLV y el cáncer de mama, o del potencial zoonótico de este retrovirus (Sinha, 2016). Zhang et al. (2016) no detectaron anticuerpos ni secuencias de BLV en muestras de sangre y tejidos de mujeres con y sin cáncer de mama en China. Asimismo, en otro estudio de secuenciamiento profundo de genoma completo efectuado sobre 51 muestras de cáncer de mama tampoco se identificaron secuencias del BLV (Gillet y Willems, 2016). A pesar de las controversias, y siendo el BLV un retrovirus emparentado con el virus de la leucemia a células T de humanos (HTLV), es necesario considerar el potencial riesgo de transformarse en una zoonosis.
DIAGNÓSTICO
En el diagnóstico de la leucosis bovina es importante diferenciar la enfermedad (leucosis o linfosarcoma) y la LP, de la infección por BLV. El diagnóstico de la enfermedad es el primer indicio de la presencia de la infección en el rodeo, mientras que el diagnóstico de la infección permitiría orientar su control.
La LP se define como un incremento en el recuento absoluto de linfocitos de tres o más desvíos estándar por encima de la media determinada para la raza y para la edad; incremento que debe mantenerse por más de tres meses. La detección de animales con LP a través de las denominadas "claves hematológicas" fue la base de un procedimiento utilizado en el pasado (previo al desarrollo de los métodos serológicos) para clasificar los animales como normales, sospechosos o linfocitóticos en función de los recuentos absolutos de linfocitos y la edad (Bendixen, 1965).
El diagnóstico de linfosarcoma se lleva a cabo a través del examen clínico o hallazgos de necropsia. Este diagnóstico debe ser confirmado mediante el análisis histopatológico de una biopsia de los órganos afectados. Sin embargo, este método no permite distinguir entre el linfoma esporádico y aquellos inducidos por BLV, por lo cual debe ser complementado con un análisis por PCR o una prueba serológica (EFSA 2015). La mayoría de los linfomas bovinos presentan un patrón difuso, con pérdida total de la arquitectura nodal. El tipo celular más frecuente es el difuso grande, con elevados índices mitóticos (Vernau et al., 1992). El análisis inmunofenotípico muestra expresión de moléculas de clase II del CMH, cadenas pesadas tipo γ y cadenas livianas λ (Vernau et al., 1997). Los animales afectados suelen tener un elevado recuento linfocitario en sangre periférica, con valores entre 20 000 y 30 000 linfocitos/µl, pudiendo en algunos casos elevarse hasta 80 000-90 000 linfocitos/µl, y más; sin embargo, en ciertos casos el recuento linfocitario puede estar disminuido (Radostitis et al., 2007).
El diagnóstico de la infección por BLV puede realizarse en forma indirecta, a través de la detección de anticuerpos específicos, o por métodos directos que evidencien la presencia del virus. La presencia de anticuerpos específicos en suero o leche, determinada mediante métodos serológicos, es una evidencia clara de infección en animales mayores de 6 meses de edad (Gutiérrez et al., 2001). Los anticuerpos específicos pueden detectarse entre 2 y 6 semanas pos-infección, dependiendo de la sensibilidad de la técnica empleada (Klintevall et al., 1997). En estas pruebas se incluyen la inmunodifusión en gel de agar (IDGA) (Miller y Van Der Maaten, 1977) y las de radioinmunoprecipitación (RIP) utilizando como antígenos las proteínas más inmunogénicas del virus, la proteína principal de la cápside de 24kDa (p24) y la glicoproteína de la envoltura viral de 51kDa (gp51) (Levy et al., 1977, 1980). Los anticuerpos contra gp51 generalmente aparecen antes y alcanzan mayores títulos que los anticuerpos antip24 (Mammerickx et al., 1985).
La prueba de IDGA ha sido ampliamente utilizada por su simplicidad y especificidad. Sin embargo, los estudios comparativos muestran que la RIP, utilizando la gp51 como antígeno, seguido de la RIP con p24, son más sensibles que la IDGA para la identificación de animales infectados (Gupta y Ferrer, 1981; Miller et al., 1981). Además, la técnica de IDGA arroja frecuentemente resultados falso-negativos en muestras de vacas en el periparto (Burridge et al., 1982) o en animales que desarrollan bajos títulos de anticuerpos (Ferrer, 1980; Gupta y Ferrer, 1981).
Posteriormente se han desarrollado varios ELISAs que han ido reemplazando gradualmente a la IDGA y al RIP (Portetelle et al., 1989; Klintevall et al., 1991; Gutierrez et al., 2001), y varios de ellos están disponibles comercialmente. La principal ventaja del ELISA respecto de la IDGA es su mayor sensibilidad, y la posibilidad de aplicarlo a muestras de suero o plasma, leche y pool de leche. La prueba en pool de leche resulta útil para monitorear el estado situacional de los rodeos lecheros en forma económica, con la ventaja de que puede detectar los niveles de anticuerpos en leche que son significativamente inferiores al del suero. Se han obtenido resultados positivos en pool de leche con prevalencias de infección de 2.5-5% o superiores (Klintevall et al., 1991; Gutierrez et al., 2001). A pesar de la marcada superioridad de algunos ELISA respecto de la IDGA, ambas pruebas son recomendadas por la OIE para el diagnóstico serológico de la infección (OIE, 2018).
Debido a la ausencia de expresión viral y viremia en los animales infectados, la identificación directa del BLV en forma de partículas virales o sus antígenos requiere el cultivo in vitro de los leucocitos de sangre periférica (Ferrer, 1980). Este procedimiento activa la expresión de transcriptos, proteínas y partículas virales, los cuales pueden ser detectados por métodos moleculares, inmunológicos o microscopía electrónica. Estas metodologías son muy laboriosas y, por lo tanto, imprácticas para el diagnóstico de rutina, por lo que su uso está prácticamente restringido al campo de la investigación científica (Gutiérrez y Forletti, 2016). Los métodos moleculares, fundamentalmente la PCR y sus variantes, son considerados actualmente como los métodos más útiles y rápidos para la detección directa del BLV.
La detección directa del provirus es el método de elección en terneros menores de seis meses (Agresti et al., 1993; Eaves et al., 1994) (ya que los anticuerpos adquiridos pasivamente a través del calostro interfieren en el diagnóstico serológico), y en los estadios iniciales de la infección, previo al desarrollo de la respuesta inmune específica. También resultan de utilidad para clarificar resultados dudosos o inconsistentes obtenidos por métodos serológicos (Fechner et al., 1996; Rola-Luszczak et al., 2013), y para diferenciar casos clínicos de linfomas esporádicos y enzoóticos (Radostitis et al., 2007). Se han desarrollado numerosas variantes de la técnica de PCR, incluyendo la amplificación simple, PCR anidada (Fechner et al., 1996; Gutierrez et al., 2001), ELISA-PCR (Naif et al., 1992; Rola y Kuzmak, 2002), in situ PCR (Duncan et al., 2005) y amplificación isotérmica (Komiyama et al., 2009; Okuwa et al., 2017). La eficiencia en la detección del provirus por las distintas variantes de PCR es variable, dependiendo principalmente del gen viral amplificado y de los cebadores (primers) utilizados (Kuckleburg et al., 2003). Los mejores resultados se han obtenido amplificando regiones de los genes pol y env (Fechner et al., 1996; Kuckleburg et al., 2003) y utilizando el formato nested o anidado, para obtener un nivel de sensibilidad aceptable.
La técnica de PCR ha mostrado ser más sensible que los métodos serológicos, principalmente debido a la detección de animales infectados en la etapa inicial de la infección, antes de la seroconversión (Klintevall et al. 1994; Fechner et al., 1996; Beier et al., 1998). La introducción, en los últimos años, de equipamiento y reactivos que permiten realizar la PCR en tiempo real ha favorecido la aplicación de esta técnica para la detección y cuantificación del BLV (Kuckleburg et al., 2003; Lew et al., 2004; Heenemann et al., 2012; Rola-Luszczak et al., 2013). Sin embargo, por razones de costo, el uso de esta tecnología está restringido casi completamente a la investigación.
Se han reportado resultados negativos por PCR en animales serológicamente positivos (Jacobsen et al., 1985; Murtaugh et al., 1991; Fechner et al., 1996; Reichel et al., 1998; Gutierrez et al., 2001). Ante esto, no debe excluirse la posibilidad de infección con BLV a partir de un resultado negativo en PCR aplicada a células de sangre. Esta situación es frecuente en animales que portan un número extremadamente bajo de copias del provirus en sangre, es decir, animales con BCP. De hecho, el nivel de copias de provirus en animales con BCP está al límite o por debajo del nivel de detección de una PCR anidado, de manera que estos animales en muchos casos son indetectables mediante esta técnica (Juliarena et al., 2017b). Con relación a la respuesta serológica, los animales con BCP desarrollan usualmente títulos de anticuerpos contra las proteínas BLVp24 y BLVgp51 significativamente inferiores que los animales de ACP (Juliarena et al., 2007). Los anticuerpos anti-BLVgp51, a pesar de encontrarse en niveles inferiores en animales con BCP respecto de los animales con ACP, son fácilmente detectables por pruebas serológicas como el ELISA, cuando se utiliza la gp51 como antígeno (Gutierrez et al., 2001). Por el contrario, los anticuerpos anti-BLVp24 son indetectables en la mayoría de los animales con BCP, o alcanzan títulos muy bajos. Por esta razón, la sensibilidad diagnóstica de las pruebas de laboratorio basadas exclusivamente en BLVp24 suele ser relativamente baja.
TRATAMIENTO Y CONTROL
Tratamiento Farmacológico
Una característica importante de la infección por el BLV es que, como todo retrovirus, el virus incorpora su información genética al genoma del hospedador, manteniéndose en un estado silente, como provirus, dentro de las células infectadas. Este es un mecanismo de evasión del sistema inmunitario, evitando así que las células infectadas sean eliminadas. Si bien hasta el momento no existe ningún tipo de tratamiento para aquellos animales infectados con el BLV, se han evaluado algunas estrategias de tratamiento en forma experimental como la administración de compuestos capaces de revertir la latencia del BLV para hacer que el virus se exprese. La expresión viral en las células infectadas permite que estas puedan ser reconocidas y eliminadas por parte del sistema inmunitario (Achachi et al., 2005).
Cierto grupo de sustancias, llamadas inhibidores de la acetilación de histonas (iAH) entre ellas, el anticonvulsivo ácido valproico (VPA) y el antibiótico tricostatina A (TSA) han demostrado tener la capacidad de activar la expresión de virus latentes (Chen y Townes, 2000). Las drogas iAH se han probado en preclínica y ensayos clínicos solas o en combinación con fármacos anti-retrovirales como terapéuticos para infecciones por el virus de inmunodeficiencia humana (VIH) y los virus de leucemia a células T de simios (STLV) y HTLV, surgiendo entonces un nuevo potencial terapéutico para retrovirus (Lehrman et al., 2005; Lezin et al., 2007; Afonso et al., 2010). Achachi et al. (2005) probaron el tratamiento con VPA en ovinos leucémicos observando un aumento transitorio de la carga proviral, una disminución en el número de linfocitos, y la regresión de la leucemia/linfoma. No obstante, a pesar de estos resultados iniciales alentadores, los animales quedaron persistentemente infectados. En un estudio de seguimiento de dos años en ovejas donde la carga proviral disminuyó inicialmente, la interrupción del tratamiento llevó a la reaparición de una leucemia incontrolable y a la falta de respuesta a la nueva administración de VPA, siendo necesario utilizar drogas más potentes (Gillet et al., 2012).
Recientemente se han reportado los resultados de un experimento piloto utilizando el inhibidor de la enzima ciclo oxigenasa-2, meloxicam, combinado con anticuerpos antiligando de muerte programada-1 (PD-L1) (Sajiki et al., 2019). Este nuevo tratamiento fue probado por estos autores obteniendo una reducción significativa de la carga proviral. Sin embargo, la evidencia de este ensayo está limitada a un solo animal, por lo que se necesitan más estudios para poder evaluar su eficacia.
Vacunas
Una solución más práctica y económica sería contar con una vacuna que prevenga la infección por BLV. Sin embargo, debido a que el BLV es un retrovirus, el desarrollo de una vacuna eficaz no es una tarea fácil. Si bien, tanto los bovinos como los ovinos, son capaces de desarrollar una fuerte respuesta inmunitaria antiviral frente a la infección primaria por el BLV, las respuestas inmunitaria humoral e inmunitaria mediada por células son deficientes para la eliminación de células infectadas que portan el virus latente (Abdala et al., 2019).
Debido a que los anticuerpos maternos transferidos a la cría a través del calostro son capaces de proteger al recién nacido de la infección durante varios meses, los primeros intentos de inmunización estuvieron dirigidos a inducir anticuerpos neutralizantes (Callebaut et al., 1993, 1994). Inicialmente, se probó una vacuna que contenía un provirus defectuoso (Altaner et al., 1987a, 1988, 1991). Esta vacuna, compuesta por células infectadas, indujo la producción de anticuerpos neutralizantes en las ovejas vacunadas, pero fue desechada al encontrarse que el provirus podía transmitirse por el contacto entre células, representando un riesgo de transmisión de la enfermedad (Altaner et al., 1987b). También se probaron vacunas a virus inactivado, pero solo una porción de los animales resistió al desafío con una dosis baja de virus infectante (Miller y Van Der Maaten, 1978; Fukuyama et al., 1993). Por otro lado, las vacunas a subunidades virales, si bien produjeron una respuesta inmunitaria humoral, también fallaron en prevenir la infección del ganado vacunado (Burkhardt et al., 1989; Cherney y Schultz, 1996).
La primera evidencia de que la respuesta mediada por células podría ser importante en la protección sobrevino con el uso de vacunas recombinantes a subunidades (env) que utilizaban vectores virales como el virus vaccinia (Ohishi et al., 1991). Esta vacuna no previno la infección con el BLV, pero redujo significativamente la diseminación del virus entre los linfocitos en los animales vacunados. En este ensayo, no se encontró una correlación con los anticuerpos neutralizantes presentes en el suero de los animales, sugiriendo que la respuesta mediada por células cumplía un rol más importante en limitar la multiplicación viral (Ohishi et al., 1992, 1996).
Hasta el momento, la vacuna perfecta que pueda inducir la producción de anticuerpos neutralizantes y una respuesta inmunitaria protectora mediada por células no ha sido alcanzada. Los animales están expuestos a la reinfección durante toda su vida (Asfaw et al., 2005), de allí que la capacidad para inducir una respuesta inmunitaria protectora de larga duración depende de un estímulo antigénico adecuado. Esto se lograría principalmente con una vacuna viva atenuada, con capacidad de replicación in vivo (Siegrist, 2013). Asimismo, la respuesta inducida por esta vacuna también debería diferenciar entre animales vacunados e infectados.
En los últimos años se ha proclamado el éxito en el desarrollo de una vacuna que utiliza un provirus con múltiples modificaciones genéticas (Rodriguez et al., 2011; Gutierrez et al., 2014; Abdala et al., 2019). Sin embargo, no se ha mostrado una prueba de concepto, así como tampoco existe información sobre las características de este provirus mutado, de la protección inmunitaria conferida a largo plazo a los animales vacunados, de la evaluación de seguridad de la vacuna, ni tampoco de la estrategia a ser utilizada para diferenciar animales vacunados de animales infectados. La nueva evidencia científica sobre la biología del virus y sobre la alteración en la función de genes represores de tumores que representa la incorporación de retrovirus en el genoma del hospedador enfatiza aún más la necesidad de precaución al momento de utilizar vacunas retrovirales vivas atenuadas (Gillet et al., 2016; Rosewick et al., 2017).
En conclusión, hasta la fecha, ninguna vacuna contra el BLV ha tenido éxito, y una vacuna preventiva adecuada tomará muchos años en materializarse. Tal como ocurre en otras enfermedades retrovirales, se ha fallado en conseguir un estímulo antigénico continuo del sistema inmunitario que genere una inmunidad esterilizante.
Estrategias de Control
Estrategias tradicionales
La estrategia de control y erradicación del BLV más eficiente consiste en la identificación y eliminación de todos los animales infectados del rodeo. Esta medida sanitaria ha tenido excelentes resultados en varios países europeos, logrando la erradicación de la enfermedad (EFSA, 2015).
En Argentina, desde 1994, se ha implementado un programa voluntario de erradicación de la leucosis bovina. La Resolución N.º 337/94 del Servicio Nacional de Sanidad y Calidad Agroalimentaria (SENASA) establece las normas y procedimientos para la certificación de establecimientos libres de leucosis (SENASA, 2005). Como primera medida se debe hacer el diagnóstico de la situación inicial por medio de una prueba serológica oficial (IDGA o ELISA) a todos los animales del rodeo mayores de seis meses a fin de clasificar al rodeo como "libre" o "infectado". Para certificar un rodeo como "libre de BLV" se deberá repetir el examen serológico a la totalidad de los animales después de 60-90 días, y se recertificará anualmente previo análisis serológico negativo de todos los animales mayores de 15 meses.
En los casos donde el establecimiento se encuentra en la categoría de "infectado", y de acuerdo con el número de animales afectados, se puede optar por la eliminación simultánea o gradual de los animales positivos. Los establecimientos se clasifican según el porcentaje de animales infectados en: Infectado Clase A (1-15% de positivos); Infectado Clase B (15-30% de positivos); Infectado Clase C (>30% de positivos). En general, el plan de saneamiento a seguir depende de la edad de los animales afectados, del porcentaje de animales infectados en el rodeo, de la infraestructura del establecimiento, de las prácticas de manejo y de las posibilidades económicas del productor.
Si la tasa de infección es baja (<10%), es conveniente eliminar los animales positivos, implementar medidas de manejo higiénico-sanitarias estrictas y realizar el control serológico cada tres meses para ir descartando los animales infectados, que pudieran haber sido no detectados en el testeo inicial (infecciones recientes). Una vez que la totalidad de los animales tiene serología negativa, se realizará un control anual, manteniendo siempre las medidas de higiene en las prácticas semiológicas y quirúrgicas habituales. El establecimiento podrá declararse libre de leucosis, una vez que se obtengan resultados negativos en la totalidad de los animales, en dos controles consecutivos. A partir de ese momento se deberá hacer un seguimiento anual. Todos los bovinos que se incorporen deben ser serológicamente negativos y mantenerse en aislamiento. Podrán incorporarse al rodeo si resultan negativos a una segunda prueba serológica realizada después de tres meses.
Si la tasa de infección es alta (>10%) se establecen medidas de control estrictas en todas aquellas prácticas que involucren transferencia accidental de cualquiera de los fluidos biológicos con el objetivo de disminuir la transmisión en el rodeo. Las medidas de bioseguridad y de manejo que se recomienda implementar son las siguientes:
Uso de agujas y jeringas descartables para extracciones de sangre e inyecciones.
Uso de guantes obstétricos individuales para palpación transrectal (o guantes separados para vacas no infectadas de vacas infectadas con BLV).
Uso de material descartable o adecuadamente desinfectado para todos los procedimientos que conlleven un riesgo de contaminación con sangre (descornes, castraciones, tatuado, etc.)
Control de insectos hematófagos.
Alimentación de los terneros con leche de vacas no infectadas con BLV, con sustitutos de leche o con leche de vacas infectadas con BLV inactivada por calor.
Utilización de inseminación artificial o monta natural con toros no infectados con BLV.
Introducción al rodeo solo de animales BLV negativos provenientes de rodeos libres de BLV, o con cuarentena de 2-3 meses para confirmar la negatividad.
La aplicación de estas medidas en rodeos donde conviven animales infectados con animales libres de BLV requiere de una minuciosa y estricta sistematización del manejo animal y, aun así, la eficacia de este enfoque puede verse socavada por factores humanos y ambientales. Como medidas adicionales a las ya descritas, se recomienda aplicar conjuntamente programas de control de vectores hematófagos, como aplicación de pesticidas sobre los animales o colocación de mallas mosquiteras en los establos donde habitan los bovinos (Ooshiro et al., 2013; Kohara et al., 2018). Se recomienda además, ordeñar las vacas infectadas al final de cada ordeñe, especialmente los animales con ACP.
Otra estrategia utilizada para mantener e incrementar el número de bovinos libres de BLV del establecimiento es dividir el rodeo en tres grupos y en corrales distantes: 1) vacas infectadas, 2) vacas no infectadas y 3) animales de reemplazo en cuarentena. Se requiere, además, del seguimiento periódico de los animales de los grupos 2 y 3 para detectar seroconversiones (Shettigara et al., 1989; Suh et al., 2005)
Estrategias alternativas de control mediante selección genética de animales resistentes
Los enfoques tradicionales de control son económicamente inviables en rodeos con alta prevalencia de infección por BLV. Para estos casos, se ha diseñado un programa de control y erradicación de BLV basado en la selección genética de animales asistida por un marcador molecular (Juliarena et al., 2017b). Se consideran alelos de resistencia al BLV a los alelos del exón 2 del gen BoLADRB3 que están asociados al fenotipo de BCP. El alelo BoLA-DRB3*0902 mostró una fuerte asociación con el fenotipo de BCP, tanto en bovinos de raza Holando Argentino como Japanese Black (Juliarena et al., 2008; Miyasaka et al. 2013; Carignano et al., 2017; Hayashi et al., 2017; Takeshima et al., 2019). Los estudios realizados en animales infectados Holando Argentino, tanto natural como experimentalmente, demostraron que la penetrancia de este alelo es notablemente alta, toda vez que más del 80% de las vacas portadoras del alelo DRB3*902 desarrollan el fenotipo de BCP (Juliarena, 2008; Juliarena et al., 2008). Además, la identificación de animales portadores de este alelo se puede realizar fácilmente mediante análisis de PCR alelo específica o por PCR en tiempo real (Forletti et al., 2013). Por lo tanto, se considera que el alelo BoLA-DRB3*0902 es un buen marcador molecular para la identificación de animales que desarrollarán BCP en las poblaciones actuales de las razas Holando Argentino y Japanese Black.
El diseño del programa de control por selección genética de animales resistentes incluye dos etapas (Figura 4). En una primera etapa, los animales con ACP (que presentan alto riesgo de transmisión de la infección) se reemplazan paulatinamente por animales con BCP o por animales no infectados, portadores de alelos de resistencia. En la segunda etapa se reemplazan progresivamente los bovinos de BCP por animales no infectados hasta declarar el establecimiento libre de BLV. En esta etapa, la genética de los animales de reposición es irrelevante; es decir, pueden ser portadores o no de alelos de resistencia, ya que todos los bovinos infectados que quedan en el rodeo presentan BCP y, por lo tanto, no transmitirán el virus. Esto último ha sido confirmado en un rodeo experimental en producción (Juliarena et al., 2016), donde las vacas de BCP portadoras del alelo BoLADRB3*0902 convivieron durante 20 meses con vacas no infectadas, sin que se haya aplicado alguna medida de control para minimizar la transmisión de BLV, teniendo en cuenta que los animales estuvieron expuestos a una gran densidad de insectos hematófagos.
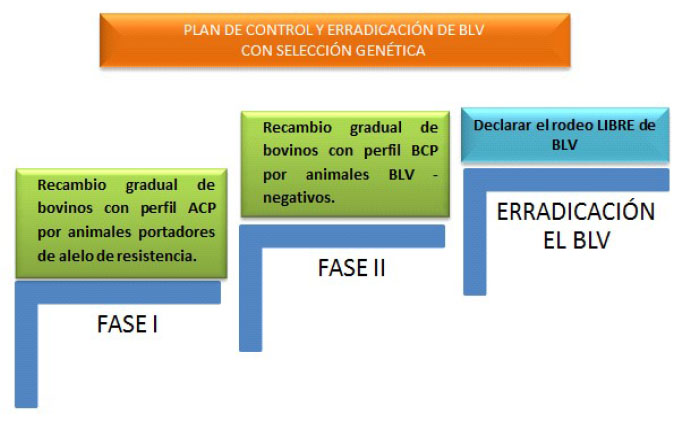
Figura 4 Esquema del diseño del plan de control y erradicación del BLV mediante selección genética de animales utilizando el alelo BoLA-DRB3*0902 como marcador molecular. (ACP: alta carga proviral; BCP: baja carga proviral)
En dicho lapso no se observó transmisión del BLV de los animales de BCP a los animales negativos, mientras que la tasa de incidencia en otros rodeos cercanos que utilizan el manejo usual para la región varió entre 0.06 a 0.17 contagios cada 100 días de contacto entre animales infectados y no infectados.
Así como se han identificado alelos de resistencia, también se han identificado varios alelos del gen BoLA-DRB3 asociados con el fenotipo de ACP, que se han denominado "alelos de susceptibilidad" (Juliarena, 2008). El alelo BoLA-DRB3*1501 es el que presenta mayor asociación con el fenotipo de ACP (Juliarena et al., 2008). Existen otros alelos de este gen que han sido identificados como alelos de resistencia o de susceptibilidad, pero su asociación con los perfiles de infección es más débil o el alelo identificado ha diferido en los diferentes estudios (Juliarena et al., 2008; Miyasaka et al., 2013; Carignano et al., 2017; Takeshima et al., 2019). Por otro lado, se observa que no todos los animales de BCP son portadores de alelos de resistencia, lo cual sugiere el efecto de algún otro factor indeterminado sobre este fenotipo (Juliarena et al., 2008). Esto también podría explicar el desarrollo de ACP en alrededor de un 20% de animales con el marcador molecular de resistencia. Se ha sugerido que pueden existir varios genes asociados con el desarrollo del perfil de infección, u otros factores epigenéticos o ambientales que pueden influenciar el desarrollo del fenotipo (Juliarena et al., 2008). Otra hipótesis es que estas asociaciones pueden simplemente ser la consecuencia del desequilibrio de ligamiento entre el marcador molecular y otros loci (Juliarena et al., 2008). Si esta última hipótesis fuera correcta, el hecho que desde hace casi 30 años el alelo *0902 (antiguamente *11) ha sido identificado como alelo de resistencia (Xu et al., 1993b; Zanotti et al., 1996; Juliarena et al., 2008; Miyasaka et al., 2013; Carignano et al., 2017; Hayashi et al., 2017; Takeshima et al., 2019), demuestra que en este caso la recombinación intracromosómica es muy baja y, por ende, el alelo BoLA-DRB3*0902 constituye el mejor marcador molecular de resistencia identificado, especialmente para la raza Holando Argentino.
Este plan de control con intervención genética ha sido puesto en práctica por una cooperativa lechera comercial, que presenta altas prevalencias de infección con el BLV en sus tambos. En un principio, la prevalencia de infección calculada para los animales de primer parto fue similar a la observada en los rodeos de primer parto de otros establecimientos de la misma cuenca lechera. Sin embargo, la incorporación del alelo BoLADRB3*0902 como criterio de selección genética en los animales de recambio, aumentó significativamente el porcentaje de vacas de primera parición con fenotipo BCP (p<0.0001), en concordancia con el aumento de la frecuencia del alelo *0902 en el rodeo (Juliarena et al., 2017a). Estos resultados muestran que, en la práctica, la aplicación de este plan de control alternativo basado en la selección genética, ya en su primera fase, puede disminuir la carga proviral promedio del rodeo.
Debido al papel relevante de los genes del complejo mayor de histocompatibilidad (BoLA) en la respuesta inmunitaria, se plantea el interrogante sobre si la aplicación de este plan de selección genética podría aumentar la susceptibilidad de las poblaciones seleccionadas por el genotipo BoLA a otros agentes infecciosos. Se conoce que los animales con genotipo de resistencia presentan títulos bajos de anticuerpos contra las dos proteínas estructurales principales del BLV, env gp51 y gag p24 (Juliarena et al., 2007). Sin embargo, estos animales han demostrado desarrollar una buena respuesta inmunitaria humoral cuando son infectados con otros patógenos prevalentes del ganado lechero (Esteban et al., 2009; Juliarena et al., 2009).
Además, hasta la fecha el alelo BoLADRB3*0902 no ha sido asociado con la susceptibilidad a otros agentes infecciosos (Lützelschwab et al., 2016; Gutiérrez et al., 2017), ni ha afectado negativamente rasgos de producción o reproducción. Más aún, los alelos de resistencia se asocian con parámetros de mayor producción y con resistencia a infecciones intramamarias (Dietz et al., 1997; Rupp et al., 2007).
La expansión de la población de vacas portadoras de estos alelos con el objetivo de controlar la infección por BLV, proporcionaría el beneficio adicional de la resistencia a la mastitis. Sin embargo, con el fin de mantener la mayor diversidad genética posible del gen BoLA-DBR3, este plan propone la aplicación de selección genética para este marcador solamente hasta que los animales de ACP hayan sido eliminados del rodeo, y se recomienda que los animales de reemplazo sean heterocigotas para el alelo *0902. La única excepción a esta recomendación es evitar introducir alelos asociados a susceptibilidad al desarrollo de ACP, especialmente el alelo *1501. Su presencia disminuye la penetrancia del alelo *0902 de un 80% a un 50%, y requiere un manejo especial de la descendencia, ya que el 50% de los descendientes son portadores de alelos de susceptibilidad.
Una de las principales limitaciones para la aplicación generalizada del plan de control propuesto es la baja disponibilidad de reproductores portadores del marcador de selección. Sin embargo, es factible lograr la rápida expansión de animales portadores del alelo BoLA-DRB3*0902 por cruzamiento dirigido o inseminación artificial, ya que a nivel poblacional (y específicamente para este gen altamente polimórfico) la frecuencia del alelo *0902 es relativamente alta (3.5-9.5%) y el patrón de herencia es codominante (Juliarena et al., 2008; Nikbakht Brujeni et al., 2016; Carignano et al., 2017; Hayashi et al., 2017; Takeshima et al., 2019). Por otro lado, la creciente demanda de semen de toros portadores del marcador ha comenzado a repercutir en las empresas comercializadoras de semen, lo que probablemente incida en la oferta de reproductores con la genética deseada a nivel internacional.
CONCLUSIONES
El BLV es un virus lento y persistente. Las elevadas prevalencias de infección que se registran en varios países generan preocupación por los efectos negativos de la infección sobre la productividad y la salud de los animales, y por su potencial riesgo para la salud humana. Hasta el momento no existe ningún tratamiento o vacuna eficaz para el control del BLV. La oportunidad de controlar la infección a través del diagnóstico y eliminación de animales infectados ya no es posible para muchos países. La selección de animales resistentes, portadores del marcador BoLA-DRB3*0902 surge como una estrategia natural para limitar la diseminación del BLV y, por ende, para el control del linfosarcoma asociado

