











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
La cría extensiva de llamas en la Puna de Jujuy (Argentina) se encuentra en zonas de menor recurso forrajero en cantidad y calidad, lo cual sumado al escaso manejo nutricional, con la consecuente disminución de la condición corporal en etapas fisiológicas reproductivas, predisponen a menores índices productivos. La mortalidad embrionaria en llamas es mayor que en otras especies domésticas, habiéndose reportado tasas de mortalidad en camélidos sudamericanos (CSA) de 12% (Leyva y García, 1999), 10-15% (Bravo et al., 1995), 50% (Fernández-Baca et al., 1970) y hasta 80% (Alarcón et al., 1990) en los primeros 90 días de gestación. Varios factores, tales como el manejo de los animales, la nutrición, el ambiente (bajas temperaturas, escasa precipitación, etc.), fallas en la migración embrionaria entre cuernos uterinos, podrían ser responsables de la alta mortalidad embrionaria.
En rumiantes, una hormona antiluteolítica producida por el trofoblasto del conceptus en el momento de su elongación hasta la forma filamentosa, conocida como interferón tau (IFNô), es la principal señal embrionaria que previene la regresión luteal (Spencer y Bazer, 2004). Sin embargo, en el género Camelus, Skidmore et al. (1994) observaron que el embrión alrededor del día 10 produce un alto nivel de estradiol, lo que podría prevenir la luteólisis y favorecer el reconocimiento materno de la preñez (RMP). Por otro lado, en llamas preñadas, Aba et al. (1997) determinaron una disminución temporaria en la progesterona (P4) plasmática después del día 8 y aumentando nuevamente a partir del día 10, siendo esa disminución coincidente con el RMP.
Estudios en otras especies domésticas (Carter et al., 2008) demostraron que la elongación del conceptus, el aumento de la producción de IFNô y, en consecuencia, una mayor tasa de preñez sería el resultado de elevadas concentraciones plasmáticas de P4 posterior al servicio. Sin embargo, Palomino et al. (2006) y Abalos et al. (2022), con inyecciones de progestina y P4 de larga acción respectivamente, administrados después del apareamiento natural en llamas, no lograron aumentar la tasa de preñez.
La hipótesis de este estudio fue que el tratamiento con P4 exógena o la administración de hCG para inducir un cuerpo lúteo accesorio (CLA) después del servicio natural, aumenta la tasa de preñez o disminuye la mortalidad embrionaria/fetal en llamas. Por lo tanto, el objetivo del presente trabajo fue evaluar el efecto de la administración de P4 exógena por medio de un dispositivo intravaginal (DIV) colocado entre los días 3 y 7 o la inyección de hCG al día 6 posterior al apareamiento natural sobre la elongación del conceptus, la concentración plasmática de P4, las tasas de preñez y mortalidad embrionaria/fetal durante la estación reproductiva en llamas de la Puna argentina.
MATERIALES Y MÉTODOS
Lugar de Estudio
El trabajo de investigación se realizó en la Estación Experimental Abra Pampa del Instituto Nacional de Tecnología Agropecuaria (INTA), provincia de Jujuy (22º 49‘S, 65º 47´W), Argentina, a una altitud de 3484 msnm, en enero-marzo de 2021 (Experimento 1) y en enero-marzo de 2022 (Experimento 2). Los procedimientos experimentales siguieron las normas éticas y de bienestar animal y fueron aprobados por el Comité Institucional para el Cuidado y Uso de Animales de Experimentación (CICUAE, Protocolos CRS-J N.º 9/19 y 13/19) del INTA.
Experimento 1
Animales y diseño experimental
Se utilizaron 82 hembras adultas destinadas a la producción de carne (105.6 ± 5.4 kg de peso vivo y condición corporal de 3.0-3.5 (escala 1-5, donde 1 = delgado y 5 = obeso), con un folículo ovárico ≥ 7 mm de diámetro determinado mediante ultrasonografía transrectal (Honda HS101V, transductor lineal 5-MHz, Japón). Se realizó servicio natural individual controlado en llamas receptivas en corrales individuales de 2 x 3 m, con machos de probada fertilidad (día -2). El promedio de duración de la cópula fue de 25.2 ± 8.3 min.
En el día 0 (ovulación) se realizó un examen ecográfico de los ovarios para observar la desaparición del folículo presuntamente preovulatorio y, por lo tanto, confirmar la ocurrencia de la ovulación. En el día 3, se realizaron ecografías para observar la presencia de estructuras ováricas (folículos y CL en desarrollo).
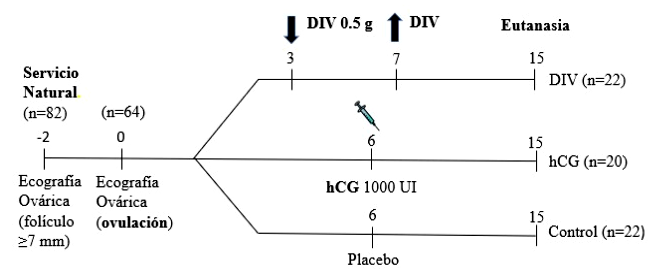
Figura 1. Esquema temporal de los tratamientos DIV, hCG y Control utilizados en el Experimento 1. Ecografías del aparato reproductor los días -2, 0, 3, 6, 7, 9, 12 y 14
Las hembras que ovularon (n = 64; 78.0%) se distribuyeron en tres tratamientos (Figura 1):
Tratamiento 1 (Grupo DIV, n = 22): se colocó un DIV impregnado con 0.5 g de progesterona (DIB 0.5g®, Syntex, Argentina), desde el día 3 hasta el día 7. Este dispositivo, fue originalmente diseñado para su uso en bovinos, pero se adaptó para su uso en llamas mediante una ligadura con hilo a la altura media de las ramas, con el fin de causar el menor efecto mecánico posible sobre la mucosa vaginal. El DIV se colocó dentro de un aplicador especial provisto por el fabricante. Antes de la inserción, se realizó la limpieza de la vulva con una esponja impregnada en desinfectante. El aplicador se lubricó con vaselina para facilitar la inserción y una vez dentro de la vagina, se depositó el DIV y se extrajo el aplicador. Antes de cada aplicación en cada hembra, el aplicador fue lavado y desinfectado.
Tratamiento 2 (Grupo hCG, n = 20): en el día 6, a las hembras que presentaron un CL desarrollado y un folículo ≥ 7 mm de diámetro, producto de la nueva onda folicular, se les administró vía IM 1000 UI de gonadotrofina coriónica humana (hCG, Ovusyn, Syntex, Argentina) para inducir la ovulación del folículo presente y el desarrollo de un CLA.
Tratamiento 3 (Grupo Control, n = 22): se administró solución fisiológica (ClNa 0.9% p/v) por vía IM en el Día 6.
Se continuó con las ecografías a todos los animales los días 7, 9, 12 y 14 para observar la presencia de estructuras ováricas (folículos y CL).
Eutanasia
Se procedió con la eutanasia de los animales (día 15) de los tres tratamientos en la sala de necropsia de la EEA Abra Pampa, de acuerdo con el procedimiento presentado en la CICUAE, con personal de la EEA con experiencia para realizar la tarea. Para la sujeción e inmovilización se hizo una manea en los cuatros miembros. La llama fue colocada en decúbito ventral y sostenida a la altura del cuello por un operario utilizando una capucha en la cara para tapar la visión. Posteriormente se procedió a la sedación y analgesia con 0.3 mg/kg de clorhidrato de xilacina 2% (Alfasan, Allignani Hnos., Argentina) por vía IM y 2.0 mg/kg de clorhidrato de tramadol 5% (Algidol Inyectable, Zoovet) por vía endovenosa (EV) en la vena yugular. Seguidamente se procedió a la eutanasia administrando 10 mg/kg de tiopental sódico (Pentovet, Richmond Vet Pharma) por vía EV. Luego se procedió a la extracción del aparato reproductor para su inmediata evaluación en el laboratorio.
Disección del ovario
Se realizó la disección de los ovarios y se pesaron individualmente (OHAUS, Scout-Pro 400 g, precisión 0.01 g). Los CL fueron separados, pesados y medidos con un vernier.
Lavado uterino y recolección de embriones
Para la recolección de los embriones se utilizó la técnica modificada descripta por Picha et al. (2013). Se colocó una pinza hemostática en el cuello del útero, se realizó una incisión en el extremo anterior del cuerno uterino izquierdo para permitir la inserción en el lumen uterino de la punta de una jeringa cargada con solución fisiológica. Una vez llenada la cavidad uterina con el medio de lavado, se hizo una incisión en el extremo anterior del cuerno derecho para recuperar el embrión dentro de un filtro especial para embriones (EmCon, EE. UU.). El embrión fue depositado en una placa de Petri, se midió su longitud con una regla graduada en milímetros y luego se observó con una lupa estereoscópica (40x) (Figura 2).

Muestras de sangre y determinaciones hormonales
Se colectaron muestras de sangre por venopunción de la yugular los días 0, 3, 6, 9, 12 y 15 para determinar la concentración plasmática de P4. El plasma se separó dentro de la hora de la extracción mediante centrifugación a 2000 g/20 min (Presvac, Argentina) y se conservó a -20 °C en tubos Eppendorf. Las concentraciones de P4 se determinaron por Quimioluminiscencia de micropartículas (CMIA - kit B/K770, Laboratorio Abbott, EE.UU.) por duplicado en un único ensayo, utilizando un analizador Abbott ARCHITECT® i2000SR, serie 15R603773 (EE. UU.), siguiendo las instrucciones del fabricante. El coeficiente de variación intra-ensayo fue <7.5% para concentraciones comprendidas entre 0.1 y 40.0 ng/ml.
Experimento 2
Se trabajó con 100 hembras adultas destinadas a producción de carne (110.3 ± 6.5 kg de peso vivo) y 3.0-3.5 de condición corporal (escala 1-5), con presencia de folículo ovárico ≥ 7 mm de diámetro, determinado por medio de ultrasonografía transrectal. Se realizó servicio natural individual controlado y único con machos de probada fertilidad (día -2) en las llamas sexualmente receptivas. La cópula tuvo una duración de 23.1 ± 6.6 min.
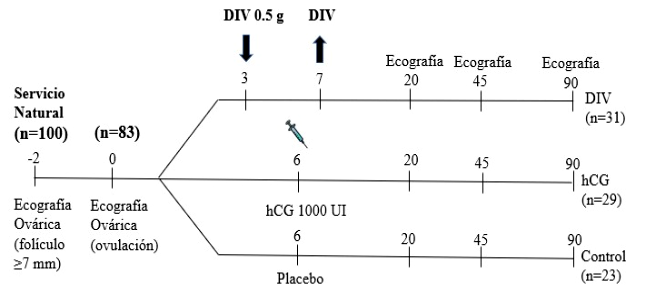
En el día 0 (ovulación) se realizó un examen ecográfico de los ovarios para determinar la desaparición del folículo presuntamente preovulatorio y confirmar la ocurrencia de la ovulación. En el día 3, las hembras que habían ovulado (n = 83) fueron distribuidas a los mismos tratamientos que en el Experimento 1; es decir, T1 (Grupo DIV, n = 31), T2 (Grupo hCG, n = 29) y T3 (Grupo Control, n = 23) (Figura 3). Se realizaron ecografías ováricas a todas las llamas cada 48 h desde el día 6 hasta el día 14 para observar la onda folicular y el crecimiento del CL. En los días 20, 45 y 90 se realizaron ecografías para el diagnóstico de gestación y evaluar la mortalidad embrionaria/fetal.
Análisis Estadístico
Se utilizó un Diseño Completamente Aleatorizado, donde las unidades experimentales (animales) fueron asignadas a los tres tratamientos de cada experimento. Se analizaron las concentraciones plasmáticas de P4 y se compararon entre los grupos control y tratamiento como medidas repetidas en el tiempo utilizando el procedimiento PROC MIXED. Los efectos fijos incluyen el tratamiento, el día y la interacción tratamiento x día. Para analizar datos de proporción (categóricos), se utilizó la prueba de Chi cuadrado procedimiento PROC FREQ. Las variables continuas fueron analizadas mediante análisis de varianza (ANOVA) utilizando el procedimiento PROC GLM. Antes de realizar el análisis, se verificaron los supuestos de normalidad, homogeneidad de varianza e independencia. Se utilizó el paquete estadístico SAS y se consideraron como estadísticamente significativas las probabilidades menores a 0.05% (p<0.05).
Experimento 1
El peso promedio de los ovarios fue mayor (p=0.006) en el tratamiento con hCG en comparación con los tratamientos DIV y Control (Cuadro 1), debido a la presencia de un CLA (Figura 4), así como un mayor número y peso total de los CLs en el tratamiento con hCG con respecto a los otros dos tratamientos. Además, el peso promedio del CL del tratamiento DIV fue significativamente menor (p=0.005) que el peso promedio del CL de los otros dos tratamientos. El peso promedio (±EE) de los CL originales (1.5±0.8 g) no fue diferente (p=0.19) del peso promedio de los CLA (1.2±0.4 g) (Cuadro 1).
Cuadro 1. Respuesta ovárica (media ± EE) y desarrollo embrionario temprano (día 15) en llamas tratadas con un dispositivo intravaginal (DIV)1 o con gonadotrofina coriónica humana (hCG)
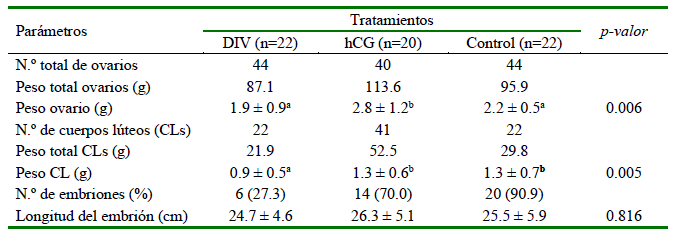
1 DIV = dispositivo intravaginal con 0.5 g de progesterona entre los días 3 y 7 posovulación
2 hCG = administración IM de 1000 UI de hCG el día 6 posovulación
En cada lavado uterino se colectó solo un embrión. No se observaron diferencias (p=0.816) en la longitud del embrión entre los tratamientos. Todas las llamas de los tratamientos DIV y Control presentaron un CL al día 15 posovulación. Las concentraciones plasmáticas de P4 se midieron únicamente en las hembras en las que se obtuvieron embriones, a fin de asegurar que las mediciones de P4 estuvieran relacionadas directamente con la presencia de embriones. La interacción tratamiento x día no fue significativa (p>0.05). El tratamiento con hCG produjo una mayor (p<0.05) concentración de P4 plasmática que los otros tratamientos en los días 12 y 15 producto de la inducción del CLA. A partir del día 6, se observó un aumento en las concentraciones plasmáticas de P4 en todos los tratamientos (Cuadro 2).
Cuadro 2. Concentración (media ± DE) de progesterona (P4) plasmática (ng/ml) en llamas con presencia de embrión según tratamiento hormonal
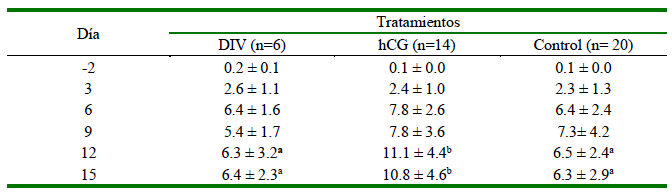
Tratamiento DIV = dispositivo intravaginal con 0.5 g de progesterona entre los días 3 y 7 posovulación
Tratamiento hCG = administración IM de 1000 UI de hCG al día 6 posovulación
a,b Valores con diferentes letras dentro de una misma fila son significativamente diferentes (p<0.05)
Experimento 2
La tasa de preñez no fue afectada por los tratamientos (Cuadro 3); sin embargo, en el diagnóstico temprano de gestación (días 20 y 45 posovulación), el grupo hCG evidenció una tendencia a una mayor proporción de hembras preñadas (p=0.06). La tasa de preñez se mantuvo en los días 20 y 45 posovulación en los tres grupos, sin detectarse diferencias (p>0.05) en el día 90 entre grupos, probablemente influenciado por una mayor pérdida fetal en el grupo hCG. Las tasas de pérdida embrionaria en los grupos DIV (5.9%, 1/17) y hCG (12.5%, 3/24) fueron similares (p>0.05). No se registraron pérdidas embrionarias en el Grupo Control. Las llamas del grupo hCG presentaron un CLA en la ecografía del día 14 posovulación.
Cuadro 3. Número y porcentaje (%) de llamas gestantes en los días 20, 45 y 90 posteriores al apareamiento natural según tratamiento hormonal
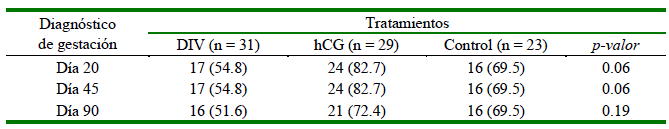
Tratamiento DIV = dispositivo intravaginal con 0.5 g de progesterona entre los días 3 y 7 posovulación
Tratamiento hCG = administración IM de 1000 UI de hCG al día 6 posovulación
DISCUSIÓN
Los resultados indican que la administración exógena de P4 por medio de un DIV o la inyección de hCG posterior al apareamiento en llamas no aumentan la longitud del embrión ni la tasa de preñez y tampoco reducen la mortalidad embrionaria/fetal.
El tratamiento con hCG se ha utilizado para producir la ovulación de un folículo en crecimiento presente en fase luteal temprana, con la formación de un CLA y, en consecuencia, aumentar la concentración de P4 en cabras (Fonseca et al., 2006), en ovinos (Bruno Vergani et al., 2020) y en bovinos (Rizos et al., 2012).
En CSA, el diámetro promedio del folículo en el día 6 de la primera onda de desarrollo folicular ovárica después de la ovulación es de 6-7 mm (Adams et al., 1990); por consiguiente, si la inyección de hCG se administra en este periodo existe una alta proba-bilidad de ovulación y formación de un CLA. En llamas, el primer trabajo realizado para inducir la ovulación y la formación de un CLA fue con la administración de un análogo de GnRH (gonadorelina) 7 días después del apareamiento, donde las concentraciones de P4 y las tasas de preñez fueron similares a las obtenidas en hembras no tratadas (Abalos et al., 2018). En el presente trabajo, el tratamiento con hCG al día 6 posterior a la ovulación, indujo el desarrollo de un CLA y un aumento en las concentraciones plasmáticas de P4, lo cual es consistente con los resultados observados en llamas tratadas con hCG en el día 7 después del apareamiento (Gallelli, et al., 2022). El desarrollo del CLA después de la aplicación de hCG fue registrado en todos los animales de ese grupo (100%), tanto en el ovario izquierdo como en el derecho, mientras que una llama presentó dos CLAs debido a la presencia de dos folículos ≥ 7 mm en el mismo ovario en el momento de la aplicación. Por otro lado, no se observaron CLAs en los otros dos grupos al no ocurrir ovulación espontánea del folículo de la primera onda folicular posterior a la ovulación inducida por la cópula en los animales.
Rosetto et al. (2023) sugirieron que la administración de P4 (0.3 g) por medio de un DIV en fase luteal temprana afecta negativamente la función del CL y podría activar prematuramente el proceso luteolítico. En el presente trabajo se observó una disminución en el peso promedio del CL en las llamas tratadas con un DIV y la consecuente disminución de la concentración plasmática de P4 luego de su retiro, en comparación con los valores registrados en las llamas del grupo Control.
En los tres tratamientos se recuperaron embriones que no estaban adheridos a la superficie endometrial al día 15 posovulación, similar a lo observado por Picha et al. (2013), pero se observó que dos embriones estaban fijados a la papila oviductal en el momento del lavado. La implantación del embrión se produciría el día 21 después de la monta (Fernández-Baca, 1971), aunque Bravo y Sumar (1981) sugirieron que el embrión permanece libre dentro del útero durante los primeros 30 días. La longitud de los embriones resultó mayor que las obtenidas por Picha et al. (2013) en alpacas, quienes observaron un cierto alargamiento a partir del día 9, en especial en los embriones que se encontraban en el cuerno uterino izquierdo. La mayor longitud de los embriones de llamas con respecto al de alpacas podría deberse a la diferencia de tamaño corporal y del aparato reproductor entre estas especies (Fowler, 1989).
La mayor concentración de P4 por la formación de un CLA en el tratamiento con hCG no se vio reflejada en una mayor longitud del embrión. Sin embargo, elevadas concentraciones circulantes de P4 en bovinos en el periodo inmediato posconcepción estuvieron asociadas con una avanzada elongación del conceptus (Carter et al., 2008), así como con un aumento en la producción de IFN-ô (Kerbler et al., 1997) y mayor tasa de preñez (Stronge et al., 2005).
En camellos, el estradiol podría desempeñar un papel clave en el mantenimiento de la función luteal durante la gestación, en donde es secretado in vitro por el conceptus durante el desarrollo embrionario temprano (Skidmore et al., 1994), al igual que los blastocistos de llamas que secretan 17βestradiol con mayor producción en su etapa de elongación entre los días 11 y 13 después de la ovulación (Powell et al, 2007). Por lo tanto, la administración de benzoato de estradiol desde el día 7 al 15 podría mejorar y extender transitoriamente la producción de P4 luteal y desempeñar un papel importante en el RMP. En CSA, con la administración de estradiol al día 8 y 9 después de la ovulación se obtuvieron resultados contradictorios; en donde mejoró la supervivencia embrionaria entre un 30-50% en alpacas (Chipayo et al., 2003; Palomino et al., 2006) o tendió a disminuir la tasa de preñez en llamas después de la transferencia de embriones (Trasorras et al., 2011).
CONCLUSIONES
La administración exógena de progesterona (P4) por medio de un dispositivo intravaginal (DIV) o la inducción de una mayor concentración plasmática de P4 por medio de la inyección de hCG al día 6 posterior a la ovulación no aumenta la longitud del conceptus determinada al día 15 ni la tasa de preñez al día 90 de gestación en llamas.
El empleo de un DIV o la administración de hCG no disminuyó la tasa de mortalidad embrionaria/fetal en llamas.

