










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología Aplicada
versión impresa ISSN 1726-2216
Ecol. apl. vol.18 no.2 Lima jul./dic 2019
https://doi.org/10.21704/rea.v18i2.1333
DOI: http://dx.doi.org/10.21704/rea.v18i2.1333
ARTICULO ORIGINAL
ASPECTOS TAXONÓMICOS Y AVANCES EN EL CONOCIMIENTO DEL ESTADO ACTUAL DEL ÁRBOL DE LA QUINA (Cinchona officinalis L.) EN EL NORTE DE PERÚ
TAXONOMIC ASPECTS AND ADVANCES IN THE KNOWLEDGE OF THE CURRENT STATE OF THE QUINA TREE (Cinchona officinalis L) IN THE NORTH OF PERU
Lizeth Huamán1,2, Joaquina Albán1 y Eder Chilquillo3
1 Departamento de Etnobotánica y Botánica Económica / Museo de Historia Natural de San Marcos / UNMSM. Av. Arenales 1256, Jesús María / Lima 14 / Perú.
2 Autor de correspondencia: lizethuaman27@gmail.com
3 Programa de Pós-Graduação em Biologia Vegetal / Instituto de Biologia / Universidade Estadual de Campinas – UNICAMP. CP. 6109, 13083-970, Campinas / SP / Brasil.
Resumen
Las quinas o cascarillas (Cinchona L.: Rubiaceae) representan uno de los recursos vegetales emblemáticos de la flora peruana, debido a su uso como agente antimalárico y a la riqueza de especies ocurrentes en nuestro territorio andino. De las 23 especies reportadas en el mundo, Cinchona officinalis L. es una especie que es confundida con C. capuli L. Andersson, C. lancifolia Mutis y C. macrocalyx Pav. ex DC. por su similitud morfológica. El hábitat donde se encuentran estas especies corresponde a bosques montanos principalmente; y la mayoría está amenazada por el crecimiento urbano y la deforestación. Por esas razones, los objetivos del estudio fueron: esclarecer sus límites taxonómicos, corroborar la existencia de C. officinalis en el Perú, estimar su estado actual de conservación y analizar su hábitat potencial. Los resultados brindaron precisiones de los caracteres diagnósticos existentes en la literatura, a fin de distinguir a Cinchona officinalis de C. capuli, C. lancifolia y C. macrocalyx. La revisión de las exsicatas depositadas en herbarios y evaluaciones de campo, ha permitido confirmar su presencia en el Perú, específicamente en la región de Piura. Los 23 reportes de presencia, obtenidos hasta la actualidad, permitieron determinar la extensión de ocurrencia (EOO = 11 166.6 km ) y el área de ocupación (AAO = 72 km2), siendo ambos valores menores a 20 000 km2 de EOO y 500 km2 de AOO. Observaciones de campo evidenciaron que la quema y el crecimiento urbano fueron amenazas latentes para su hábitat. De este modo, se plantea que el estado actual de C. officinalis es En Peligro (EN) según los criterios de la IUCN (2012, Versión 3.1; 2017, Versión 13). Adicionalmente, su hábitat parece estar confinado a la parte alta de Piura y algunas pequeñas áreas limítrofes de Cajamarca según el algoritmo de Maxent (AUC = 997); y ello reflejaría el inicio de la ejecución de futuros planes de manejo sostenible y conservación.
Palabras clave : Cinchona officinalis, Quina, hábitat, extensión de ocurrencia, área de ocupación y estado de conservación.
Abstract
The quinas or cascarillas (Cinchona L.: Rubiaceae) represent one of the emblematic plant resources of the Peruvian flora, due to its use as an antimalarial agent and the richness of species that occur in our Andean territory. Of the 23 species reported in the world, Cinchona officinalis L. is a species that is confused with C. capuli L. Andersson, C. lancifolia Mutis and C. macrocalyx Pav. ex DC. due to morphological similarity. The habitat where these species are found corresponds mainly to montane forests; and the majority are threatened by urban growth and deforestation. For these reasons, the objectives of the study were: to clarify taxonomic limits, corroborate the existence of C. officinalis in Peru, estimate its current state of conservation and analyze its potential habitat. The results provided details of the diagnostic characteristics existing in the literature, in order to distinguish Cinchona officinalis from C. capuli, C. lancifolia and C. macrocalyx. The review of exsicatas deposited in herbariums and field evaluations has confirmed their presence in Peru, specifically in the Piura region. The 23 presence reports, obtained to date, allowed to determine the extent of occurrence (EOO = 11 166.6 km2) and the area of occupation (AAO = 72 km2), both being values less than 20 000 km2 of EOO and 500 km2 of AOO. Field observations showed that burning and urban growth were latent threats to their habitat. Thus, it is stated that the current status of C. officinalis is Endangered (EN) according to the IUCN criteria (2012, Version 3.1; 2017, Version 13). Additionally, its habitat seems to be confined to the upper part of Piura and some small border areas of Cajamarca according to the Maxent algorithm (AUC = 997); and this should reflect the beginning of the execution of future plans for sustainable management and conservation.
Key words: Cinchona officinalis, Quina, habitat, extent of occurrence, area of occupation and state of conservation.
Introducción
La familia Rubiaceae en el Perú comprende 110 géneros y 782 especies, siendo considerada una de las familias de mayor importancia económica en el país (Albán, 2015). Incluye especies representativas como el café (Coffea arabica L.), uña de gato (Uncaria guianensis (Aubl.) J.F. Gmel.) y las quinas o cascarillas (Cinchona L.), las cuales constituyen un recurso vegetal emblemático del Perú debido a su aporte mundial para el tratamiento efectivo de la malaria (Pavón & Ruiz, 1798; Lambert et al., 1821; Steele, 1964).
Cinchona L., en su circunscripción actual está formada por 23 especies de árboles o arbustos (Andersson, 1998). Representa un género monofilético perteneciente a la tribu Cinchoneae, relacionado con los géneros Ladenbergia Klotzsch y Remijia DC. (Andersson, 1995b; Andersson & Antonelli, 2005; Manns & Bremer, 2010). Su distribución se extiende desde Costa Rica y parte de las cadenas montañosas de Venezuela a lo largo de los Andes Tropicales hasta el centro de Bolivia; pero se concentra al sur de Ecuador y norte de Perú (Andersson, 1995a).
Las montañas y cordilleras peruanas son consideradas centros de diversidad y endemismos de especies. Entre los principales centros de diversidad de Cinchona destaca la región biogeográfica de Amotape – Huancabamba (Weigend, 2004). Se extiende de este - oeste a través de la cordillera nor-andina del Perú, abarcando los departamentos de Piura y Cajamarca (Quintana et al., 2017; Weigend et al., 2010). Actualmente, esta región biogeográfica representa un área sometida a fuertes presiones debido a la extensión de monocultivos, ganadería, quemas, construcción de carreteras y crecimiento urbano (Gobierno Regional de Piura, 2012).
Sin embargo, reportes botánicos recientes sugieren la ocurrencia de 11 especies de Cinchona (Albán, 2015) en Piura y Cajamarca, equivalente al 48% del total de especies del género en el Perú. Cinchona officinalis L. es una de estas especies y se conoce como el árbol de la quina o cascarilla gracias a sus propiedades medicinales contra la malaria. Esta especie suele ser confundida con C. capuli L. Andersson, C. lancifolia Mutis y C. macrocalyx Pav. ex DC, debido a la similitud del porte, el color de la flor y color de las hojas juveniles (rojizas). Con relación al estado de conservación de C. officinalis, se conoce que aún no ha sido evaluada con los criterios de la IUCN (2012, 2017); mientras, otras cinco especies si fueron evaluadas (Cinchona pyrifolia, C. capuli, C. mutisii y C. rugosa).
La necesidad de conocer el estado de conservación de C. officinalis se deba que, históricamente, esta especie fue explotada y se deduce que hoy en día quedarían remanentes de bosque. Existen varias teorías sobre el inicio de la explotación de Cinchona officinalis y entre las que destacan: (i) En 1737, cuando se redescubre la cascarilla durante la Misión Geodésica Francesa con Charles Marie de La Condaime, (ii) En 1631, cuando la esposa del virrey de Perú conde de Cinchón, llamada Cinchona, se cura con la corteza de la quina, (iii) En 1638, cuando Cinchona en llevada a Roma por el jesuita Alonso Messia Venegas.
En la colonia, la explotación y comercio de la cascarilla estuvieron focalizados en el norte de Perú y sur de Ecuador, mientras que el centro de producción fue en la zona de Loja, en los montes de Uritusinga y Caxanuma. Inicialmente, la explotación de la quina estuvo dirigida por los jesuitas, quienes realizaban una extracción controlada; pero, después de su expulsión en 1767, la explotación incrementó debida que, en 1839, Willian Hooker ordenó la tala completa de los árboles, en lugar de extraer la corteza superficial, provocando así la explotación hasta los límites de insostenibilidad (Jaramillo & Carrera, 2008).
Lamentablemente, la complejidad del reconocimiento del árbol de la quina en campo significó una de las complicaciones que enfrentó la explotación de este recurso (Tapia, 2013; citando a Garmendia, 1999), y por ello su extracción resultaba en una tala indiscriminada. La extracción de C. officinalis disminuyó cuando los científicos Pierre Pelletier y Joseph Caventou en 1820 lograron sintetizar el alcaloide de la quinina que se encuentra en la corteza del árbol. Pero, este suceso, no quiere decir que los estragos de su explotación terminaron, porque la resistencia a fármacos ha despertado interés en descubrir otros alcaloides en esta especie o que su concentración sea mayor en otros órganos diferentes a la corteza.
En ese sentido, la presente investigación pretende complementar algunos caracteres, adicionales a los ya existentes, para esclarecer los límites taxonómicos de C. officinalis. Asimismo, observaciones de campo y registros, obtenidos hasta la actualidad, permitirán corroborar la presencia de la especie en Perú y estimar su estado actual de conservación. Adicionalmente, se brindará un primer alcance sobre su hábitat potencial.
Materiales y métodos
Datos de Ocurrencia
Se obtuvieron los registros de C. officinalis y especies afines (C. capuli L. Andersson, C.lancifolia Mutis y C. macrocalyx Pav. ex DC) a partir de las expediciones realizadas entre los años 2004 y 2018 en las provincias de Jaén, San Ignacio, departamento de Cajamarca; provincias de Bongará, Chachapoyas, departamento de Amazonas; provincias de Moyobamba, Rioja, San Martin, departamento de San Martín; provincias de Ayabaca, Huancabamba, departamento de Piura; provincias de Chanchamayo, Satipo, departamento de Junín; provincia de Oxapampa, departamento de Pasco; provincia de Huánuco, departamento de Huánuco; provincia de Sandia, departamento de Puno. Las colectas botánicas obtenidas de las expediciones fueron examinadas y posteriormente depositadas al herbario USM. Se complementaron los registros con la revisión de exsicatas de los herbarios de Perú (HOXA, MOL, UNC y USM) y fotos de ejemplares de los herbarios internacionales (BM, GB, F, MA, MO, NY, P, QCNE y US). Los ejemplares tipo fueron revisados en línea a través de la página web de JSTOR Global Plants.
Delimitación morfológica y morfométrica
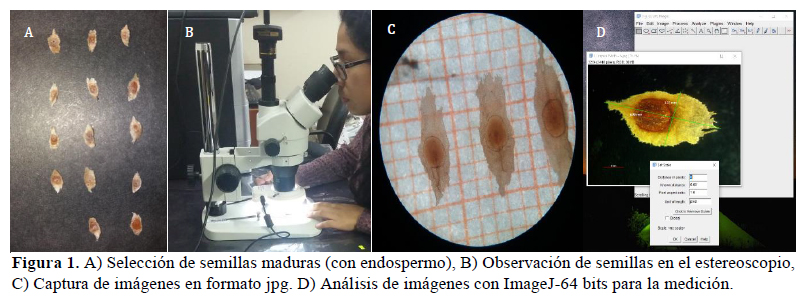
Se analizaron doce caracteres obtenidos a partir de las descripciones de Andersson (1995a, 1998) (Tabla 1). Cabe precisar que la descripción de los caracteres ha sido complementada con observaciones de muestras vivas y herborizadas. Se examinaron 200 ejemplares colectados en campo, de los cuales 50 correspondieron a cada especie (C. officinalis, C. capuli, C. lancifolia y C. macrocalyx). Los caracteres de indumento de la hoja, forma de domacio y márgenes de la semilla fueron observados en estereoscopio, mientras que el tamaño de limbo, tamaño del fruto y de la semilla fueron medidos en el programa ImageJ 64bits (Schindelin et al., 2012) (Figura 1). Para la determinación de las diferencias de los caracteres entre las especies, se empleó la prueba estadística de Kruskal-Wallis. Esta prueba es muy empleada en estudios de morfometría (Rohlf, 1990; Martínez et al., 2019), ya que permite identificar diferencias en la distribución de caracteres entre especies; y es calculada mediante la función kruskal.test del stats R Package (Hollander et al., 2013). Adicionalmente, el análisis de cluster mostró la similitud de caracteres entre las especies.

Aproximación del estado actual de conservación de Cinchona officinalis
Fue estimado a partir de los valores de la extensión de ocurrencia (EOO) y el área de ocupación (AOO) según los criterios de la IUCN (2012, 2017). La EOO y el AOO fueron delimitados en el software en línea GeoCAT (Bachman et al., 2011). Además de los valores de EOO y AOO, se consideró la información registrada mediante observaciones de campo, el cual aportó en la interpretación del estado de conservación. Esta información consistió en registrar, de manera descriptiva, la presencia de actividades antrópicas y/o climáticas que amenazan las poblaciones de la especie.
Estimación del hábitat potencial de Cinchona officinalis
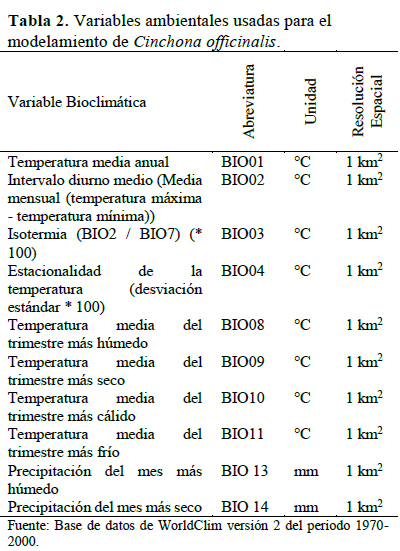
Fue obtenido a partir de los 23 puntos de ocurrencia y 10 variables bioclimáticas en formato raster de 1 km2 de resolución (Hijmans et al., 2005). Ambos datos fueron analizados con el algoritmo Maxent v.3.4.1 (Phillips et al., 2006), debido a su alta capacidad predictiva con pocos registros de presencia (Elith et al., 2006; Hernández et al., 2006; Pearson et al., 2007; Sarkinen et al., 2013). Las variables climáticas usadas fueron previamente seleccionadas con el coeficiente de correlación de Spearman (r < 0.8) (Dormann et al., 2013) (Tabla 2). La validez del modelo fue evaluado acorde a las curvas de respuesta (análisis de omisión/comisión) y sensibilidad Curva Operada por el Receptor (ROC)-Área Bajo la Curva (AUC) (Elith et al., 2006; Aguirre & Duivenvoorden, 2010) y pruebas Jackknife para medir el efecto de cada variable en el modelo (Hijmans et al., 2005; Ramos-Dorantes et al., 2017). El modelo obtenido en MaxEnt fue reclasificado en ArcMap 10.2, para obtener superficies en los diferentes niveles de probabilidad y con ello obtener el perfil bioclimático de la especie. Finalmente, la superposición del perfil bioclimático con otras variables como altitud y cobertura vegetal permitieron obtener una delimitación más acertada del hábitat potencial de la especie, ya que hay áreas donde la especie no podría encontrarse, tales como: áreas desprovistas de cobertura y elevaciones mayores a 2 800 m.
Resultados
>Delimitación morfológica y morfométrica de C. officinalis y afines
Cinchona officinalis L. Tipo: La Condamine, Mém. Acad. Roy, Sci. (París) 1738: 1.5-6. 1738 (holotipo, epitipo, designado en Ecuador. Loja: Cerro de Cajanuma, La Condamine s.n. [P-JU 9899 + B)].
Cinchona officinalis se caracteriza por su porte arbustivo (≤ 6 m), con ramas pubérulas cuando son juveniles y glabras a subglabras cuando son adultas, estípulas interpeciolares con coléteres dispuestos en varias filas, hojas elípticas de consistencia coriácea cuando están frescas y cartácea cuando se secan, domacios en el envés de las hojas profusamente huecos con pelos en los márgenes, inflorescencias terminales congestas, cáliz con lóbulos que alcanzan más de la mitad de su longitud, corolas de color rosado con lóbulos densamente vilosos en el interior, cápsulas cilíndricas con dehiscencia acropétala y semillas diminutas aladas de borde entero a ligeramente dentado (Figura 2).

La especie fue registrada entre los 2 400 y 2 800 msnm en laderas montañosas con suelo franco arenoso, abundante materia orgánica y vientos muy fuertes. Arcytophyllum thymifolium (Ruiz & Pav.) Standl. (Rubiaceae) y Arcytophyllum filiforme (Ruiz & Pav.) Standl. (Rubiaceae), Elaeagia utilis (Goudot) Wedd. (Rubiaceae) y Blechnum glandulosum Kaulf. ex Link (Blechnaceae) fueron especies frecuentes asociadas a su hábitat. La época de floración de C. officinalis se registró entre mayo y junio; y la de fructificación entre julio y agosto.
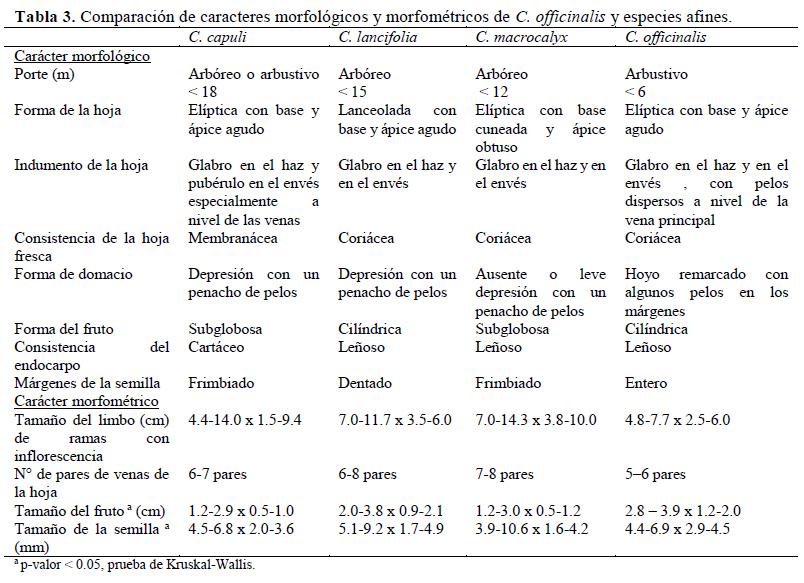
C. officinalis suele confundirse con otras especies similares como C. capuli, C. lancifolia y C. macrocalyx, debido a que comparten el hábitat en áreas circundantes a la región Amotape – Huancabamba Andersson (1998) y el rango altitudinal (Albán, 2015). Morfológicamente también son confundidas por el porte, la forma del limbo, el color de la flor, la longitud de la corola y el cáliz y la consistencia de la hoja. Sin embargo, existen doce caracteres diagnósticos descritos por Andersson (1998) que ayudan a la diferenciación de estas especies, los cuales fueron obtenidos a partir de exsicatas herborizadas. El presente estudio complementó estos caracteres con precisiones obtenidas a partir de observaciones de campo. Los caracteres morfológicos precisaron diferencias en el margen de la semilla, consistencia del endocarpo y forma del domacio, mientras que los caracteres morfométricos analizados por pruebas estadísticas de Kruskal-Wallis (KW) difícilmente mostraron diferencias claras; pese a que su p-valor fue menor a 0.5 (Tabla 3). Sin embargo, el análisis de clúster (con el índice de morisita) ha demostrado dos grupos: (i) grupo 1 compuesto por C. officinalis y C. lancifolia y (ii) grupo 2 compuesto por C. capuli y C. macrocalyx; y ciertamente es lo esperado porque las hojas del segundo grupo son generalmente más grandes que el grupo 1; caso contrario ocurre con los frutos, puesto que los frutos del grupo 1 son más anchos por ser globosos a diferencia del grupo 2 que son más aplanados o subglobosos. Adicionalmente, se debe mencionar que el número de pares de venas en el grupo 2 es mayor que en el grupo 1, y el tamaño de la semilla de C. officinalis fue menos alargada que C. capuli y C. lancifolia, pero más pequeña que C. macrocalyx.
Datos de Ocurrencia
Se obtuvieron 23 datos de ocurrencia de Cinchona officinalis, de los cuales ninguno correspondió a registros de Tropicos.org ni de Zevallos (1989). Los cinco reportes registrados en Tropicos.org correspondían a C. macrocalyx (De la Cruz et al., 5171, Schunke 10484), C. pubescens (Alyse y Samuel 123, Meyer y Chait 525) y C. capulí (Hodge 6056); y los ejemplares de Zevallos (1989) correspondían también a otras especies del género como, por ejemplo, el de Junín que correspondió a Cinchona nitida, de Huánuco, Lambayeque y Pasco a C. pubescens, de Piura a C. capuli, y de Cajamarca a C. macrocalyx.
Aproximación del estado actual de conservación de Cinchona officinalis
Los datos de ocurrencia indican que C. officinalis posee una EOO de 11 166.6 km2 y AOO de 72 km2 a escala mundial. En el Perú, su AOO y EOO son aún menores (16 km2 en ambos casos). Búsquedas activas para la especie en 18 localidades de seis distritos de los departamentos de Piura, Cajamarca y Amazonas no han sido exitosas, lo que indica que hay un intervalo bajo de incertidumbre sobre los parámetros de AOO y EOO calculados. Además, se observó que sus poblaciones han sido afectadas por la quema, la construcción de nuevas vías de acceso, la expansión de la agricultura y la ganadería. Por ejemplo, se evidenciaron quemas de laderas completas que afectaron aproximadamente 200 individuos de C. officinalis, en su mayoría adultos que juveniles y plántulas, en las localidades de la región de Piura (ex. Cruz Chiquita, Sóndor, Piura) (Figura 3). En base a las amenazas y los valores de AOO y EOO, la especie cumple para estar En Peligro (EN B1b (iii); B2b(ii, iii); C2a(i)) de acuerdo a los criterios de la UICN(2012, Versión 3.1; 2017, Versión 13).

Estimación del hábitat potencial de Cinchona officinalis
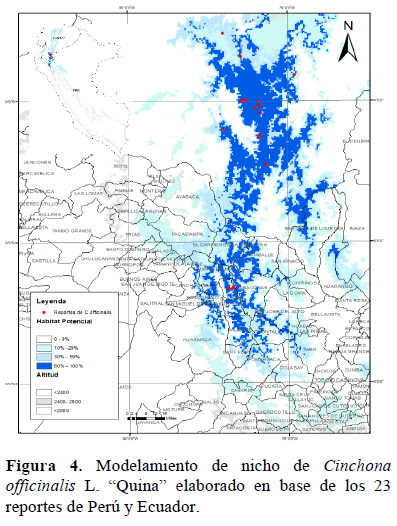
El modelamiento preliminar (AUC = 0.994) que consta de 18 puntos de ocurrencia de la especie para el sur de Ecuador, sugirió nichos potenciales situados en el norte del Perú, específicamente en los departamentos de Piura y Cajamarca. Diez variables climáticas y la elevación han permitido identificar que el hábitat donde se desarrolla adecuadamente la especie es en ambos departamentos, y no en otros departamentos del Perú, como si lo hacen las demás especies del género Cinchona. Posteriormente, un modelamiento más detallado que incluye cinco nuevos reportes de ocurrencia (Caserío Cruz Chiquita y Shilcaya) en el Perú (AUC = 0.997), mostró nichos más restringidos para la especie en varias localidades de los mismos departamentos (Figura 3). Observaciones de campo y análisis de exsicatas demostraron la existencia de C. officinalis en las localidades de Cruz Chiquita y Shilcaya, pertenecientes al distrito de Sóndor, provincia de Huancabamba, departamento de Piura, corroborando así las inferencias del modelamiento en las áreas de mayor ocurrencia. El modelamiento también indica que existen hábitats adecuados para la especie en los distritos de Sallique, Tabaconas, Namballe, San José de Lourdes, (Cajamarca), y en los distritos de Sondorillo, Carmen de la Frontera y Huancabamba (Piura) (Figura 4). De confirmarse la presencia de poblaciones en posteriores estudios, la categoría disminuiría a Vulnerable (VU B1b (iii); B2b (ii, iii); C2a (i)), ya que podría ubicarse dentro de áreas protección como Bosques de Montanos y Páramos de Huaricancha y del Santuario Nacional Tabaconas- Namballe.
Discusión
El margen de la semilla, la forma del domacio y la consistencia del endocarpo del fruto fueron los caracteres morfológicos que ayudaron a distinguir C. officinalis de las especies más afines. Sin embargo, los caracteres morfométricos aportaron poco en la diferenciación taxonómica, debido a que las poblaciones de las especies en distintas localidades presentaron alta plasticidad fenotípica de adaptación según las condiciones ambientales de cada localidad. Pero, es importante mencionar que los rangos morfométricos presentados en el presente estudio se basaron a la medición de un mayor número de ejemplares que la revisión de Andersson (1998); y además se trató de muestras observadas en campo y mediciones de material fresco. No se consideraron medidas detalladas de las flores porque Cinchona es uno de los géneros con alto grado de heterostilia (Ganders 1979; Barrett & Richards, 1990), que hace que esta estructura sea menos útil para esclarecer los límites taxonómicos. Se considera que el estudio de la heterostilia aportaría en la comprensión de la forma de propagación, en algunos casos la resiliencia, del género.
Actualmente, se conoce la existencia de C. officinalis en pequeños bosques fragmentados del distrito de Sóndor, provincia Huancabamba, departamento de Piura. Sin embargo, varias poblaciones están siendo afectadas por la quema y el crecimiento urbano, y en general esto suele ocurrir en bosques montanos y páramos de la provincia de Huancabamba (Rasal et al., 2012), lo cual es alarmante. Lamentablemente, otro factor que influye negativamente en la supervivencia de la especie es la naturaleza de su semilla. Aun cuando su tamaño pequeño, peso ligero y forma favorecen su dispersión, sus rasgos internos y la poca disponibilidad de recursos impiden tener éxito en la germinación (Weigend et al., 2005; Romero, 2015; Ocaña & Lombardi, 2016). Ello explica por qué no se observó abundancia de plántulas durante las expediciones realizadas entre el 2016 y 2017.
Basados en los análisis de modelamiento de habitat y las observaciones de campo, se corrobora la presencia de C. officinalis en el distrito de Sóndor, provincia de Huancabamba, departamento de Piura. Sin embargo, a la fecha no se ha registrado su presencia en los distritos de Huancabamba, El Carmen de la Frontera, Morropón, San José de Lourdes y San Ignacio; en donde si reportamos C. capuli, C. lancifolia, C. macrocalyx y C. scrobiculata. Esto se debe a que las características fisicoquímicas del suelo pudieron haber sido otro factor determinante, a parte de variables climáticas y elevación, en la distribución de la especie. Por ejemplo, poblaciones de C. officinalis se encontraron sobre suelo con escasa materia orgánica, menor porcentaje de humedad y en áreas más abiertas o claros con vientos fuertes; mientras que las demás especies se encontraron sobre suelos con mayor porcentaje de materia orgánica y biomasa y en zonas de sotobosque que son más húmedas. Posteriores estudios de suelos, podrían confirmar dicha hipótesis que fue formulada a partir de observaciones de campo; así como afinar el presente modelamiento con variables edafológicas. Por otro lado, los distritos de Sallique, Sondorillo, Tabaconas y Namballe serían nuevas áreas para explorar en futuros estudios.
Cinchona officinalis parece estar confinada a la parte alta del departamento de Piura y algunas pequeñas áreas limítrofes del departamento de Cajamarca (ex. Santuario Nacional Tabaconas Namballe). Los registros actuales permitieron afirmar que la especie actualmente no se distribuye en los departamentos de San Martin, Huánuco, Pasco, Junín, Madre de Dios y Puno, como fue citado en otros estudios (Zevallos, 1989; Campos-Ruíz et al., 2014).
Si bien la presente investigación ha brindado una primera aproximación de nuevas áreas (Sallique, Sondorillo, Tabaconas y Namballe) donde podría encontrarse individuos o poblaciones de la especie, es necesario que futuras investigaciones puedan afirmar aquellas predicciones; y de esta manera pueda servir para iniciar programas de conservación y/o reforestación de la especie; e incluso generar nuevos ingresos a comunidades campesinas y/o nativas a través del uso sostenible del recurso. Asimismo, es necesario que se realicen estudios de biología floral, polinizadores y citogenética para comprender mejor su mecanismo de supervivencia.
Conclusiones
El margen de la semilla, la forma del domacio y la consistencia del endocarpo del fruto permiten distinguir a Cinchona officinalis L. de especies afines. Además, Cinchona officinalis parece restringirse al departamento de Piura y zonas limítrofes de Cajamarca en Perú, aparte de su área de distribución principal en el sur de Ecuador. Finalmente, se plantea que la especie se encuentra En Peligro según los criterios de la IUCN (International Union for Conservation of Nature) (2012, Versión 3.1; 2017, Versión 13).
Agradecimientos
Agradecemos gentilmente al Consejo Nacional de Ciencia, Tecnología e Innovación del Perú (CONCYTEC) y el Vicerrectorado de Investigación de la Universidad Nacional Mayor de San Marcos (UNMSM) por el soporte económico para la presente investigación. Asimismo, queremos agradecer al Museo de Historia Natural de la UNMSM, Laboratorio de Dendrología – Universidad Nacional Agraria La Molina (UNALM) y el Instituto Regional de Desarrollo.
- IRD-Selva de la UNALM. Agradecemos a los curadores de los herbarios USM, MOL, HOXA por la colaboración científica. Finalmente agradecemos a los revisores anónimos por las sugerencias brindadas al manuscrito original.
Literatura citada
Aguirre G. & Duivenvoorden J. 2010. Can we expect to protect threatened species in protected areas? A case study of the genus Pinus in México. Revista Mexicana de Biodiversidad, 81: 875-882. [ Links ]
Albán J. 2015. Etnobotánica de Rubiaceae peruanas. Tesis para optar el grado de Doctor. Universidad Nacional Mayor de San Marcos. Lima. Perú [ Links ].
Andersson L. 1995a. Diversity and origins of Andean Rubiaceae. Páginas 441-450. En: Churchill, S.P., Balslev, H., Forero, E. y Luteyn, J.L. (eds) Biodiversity and Conservation of Neotropical Montane Forest. New York Botanical Garden, New York. [ Links ]
Andersson L. 1995b. Tribes and genera of the Cinchoneae Complex (Rubiaceae). Annals of the Missouri Botanical Garden. 82(3): 409–427. DOI:https://doi.org/10.2307/2399891Links ]org/10.2307/2399891" target="_blank">.
Andersson L. 1998. A revision of the genus Cinchona (Rubiaceae-Cinchoneae). Memoirs of the New York Botanical Garden. 80(1): 1–75. [ Links ]
Andersson L. & Antonelli A. 2005. Phylogeny of the tribe Cinchoneae (Rubiaceae), its position in Cinchonoideae, and description of a new genus, Ciliosemina. Taxon. 54(1): 17–28. DOI: https://doi.org/10.2307/25065299Links ]org/10.2307/25065299" target="_blank">.
Bachman S., Moat J., Hill A.W., Torre J. De & Scott B. 2011. Supporting Red List threat assessments with GeoCAT : geospatial conservation assessment tool. ZooKeys. 150: 117–126. DOI:https://doi.org/10.3897/zookeys.150.2109Links ]org/10.3897/zookeys.150.2109" target="_blank">.
Barrett S. & Richards J. 1990. Heterostyly in tropical plants. Memoirs of the New York Botanical Garden 55: 35-61. [ Links ]
Campos-Ruíz J., Cerna-Rebaza L. & Chico-Ruíz J. 2014. Efecto del ácido giberélico, nitrato de potasio y agua de coco en la germinación de semillas de quina, Cinchona pubescens. Rebiolest. 2(1): 1-11. [ Links ]
Dormann C., Elith J., Bacher S., Buchmann C., Carl G., et al. 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography. 36(1): 27–46. DOI: https://doi.org/10.1111/j.1600-0587.2012.07348.x [ Links ]
Elith J., Graham C., Anderson R., Dud´ık M., Ferrier S., et al. 2006. Novel methods improve prediction of species’ distributions from occurrence data. Ecography. 29: 129–51. DOI: https://doi.org/10.1111/j.2006.0906-7590.04596.x.
Ganders F. 1979. The biology of heterostyly. New Zealand Journal of Botany 17: 607-635. [ Links ]
Gobierno Regional de Piura. 2012. Estrategia Regional y Plan de Acción para la Conservación de la Diversidad Biológica de la Región de Piura. Piura, Perú: Grupo Técnico de Diversidad Biológica - Comisión Ambiental Regional de Piura. [ Links ]
Hijmans R., Cameron S., Parra J., Jones P. & Jarvis A. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965-1978. [ Links ]
Hernández P., Graham C., Master L. & Albert D. 2006. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography. 29: 773-785. [ Links ]
Hollander M., Wolfe D. & Chicken E. 2013. Nonparametric Statistical Methods. Edition 3. Wiley Series in Probability and Statistics. John Wiley & Sons Inc. Nueva York. [ Links ]
IUCN (International Union for Conservation of Nature). 2012. IUCN Red List Categories and Criteria. Version 3.1, Second edition. Gland, Switzerland and Cambridge, UK. [ Links ]
IUCN (International Union for Conservation of Nature). 2017. Guidelines for using IUCN Red List Categories and Criteria: Version 13. Prepared by the Standards and Petitions Subcommittee. URI:http://nc.iucnredlist.org/redlist/content/attachment_files/edListGuidelines.pdfLinks ]iucnredlist.org/redlist/content/attachment_files/RedListGuidelines.pdf" target="_blank">.
Jaramillo J. & Carrera D. 2008. Una aproximación CTS al análisis de la industria de la quinina en la zona de Loja en el siglo XVIII. En Cortocircuito, 17: 58-64. Recuperado de: http://200.0.29.102/material/migracionOA/UTPL/Tecnica/Electronica/19/Revista%20EnCORTOCIRCUITO17(Agosto2008).pdf y/o http://docplayer.es/8212988-Una-aproximacioncts-al-analisis-de-la-industria-de-la-quinina-en-lazona-de-loja-en-el-siglo-xviii.htmlLinks ]es/8212988-Una-aproximacioncts-al-analisis-de-la-industria-de-la-quinina-en-lazona-de-loja-en-el-siglo-xviii.html" target="_blank">.
Lambert A.B., Humboldt A. von, Laubert C.J. & Ruiz H. 1821. An illustration of the GENUS CINCHONA; comprising descriptions of all the Officinale Peruvian barks, including several new species. Searle. London. DOI: https://doi.org/10.5962/bhl.title.715Links ]org/10.5962/bhl.title.715" target="_blank">.
Manns U. & Bremer B. 2010. Towards a better understanding of intertribal relationships and stable tribal delimitations within Cinchonoideae s.s. (Rubiaceae). Molecular Phylogenetics and Evolution. 56(1): 21–39. DOI: https://doi.org/10.1016/j.ympev.2010.04.002Links ]org/10.1016/j.ympev.2010.04.002" target="_blank">.
Martínez-C., Isaza C. & Betancur J. 2019. Distribución espacial y estructura de la población de Pitcairnia huilensis (Bromeliaceae) en el valle alto del río Magdalena (Huila, Colombia). Caldasia, 41(1): 165-178. DOI: https://doi.org/10.15446/caldasia.v41n1.71328Links ]org/10.15446/caldasia.v41n1.71328" target="_blank">.
Ocaña J.C. & Lombardi I. 2016. Análisis de la vegetación en un ecosistema de montaña en el Alto Piura (Mijal, Chalaco, Morropón). Revista Forestal del Perú. 31(1): 30–41. [ Links ]
Pavón J. & Ruiz H. 1798. Flora Peruviana, et Chilensis, Sive Descriptiones, Et Icones Plantarum Peruvianarum, Et Chilensium, Secundum Systema Linnaeanum Digestae, Cum Characteribus Plurium Generum Evulgatorum Reformatis. Tomus 1. Typis Gabrielis de Sancha. DOI: https://doi.org/10.5962/bhl.title.814Links ]org/10.5962/bhl.title.814" target="_blank">.
Pearson R., Raxworthy C., Nakamura M. & Peterson A. 2007. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. Journal of Biogeography 34: 102–117. [ Links ]
Phillips S.J. Anderson R.P. & Schapire R.E. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling. 190(3–4): 231–259. DOI: https://doi.org/10.1016/j.ecolmodel.2005.03.026Links ]org/10.1016/j.ecolmodel.2005.03.026" target="_blank">.
Quintana C., Pennington R.T., Ulloa C.U. & Balslev H. 2017. Biogeographic barriers in the Andes: Is the Amotape— Huancabamba zone a dispersal barrier for dry forest plants? Annals of the Missouri Botanical Garden. 102(3): 542–550. [ Links ]
Ramos-Dorantes D.B., Villaseñor D., Ortiz E. & Gernandt D. 2017. Biodiversity, distribution, and conservation status of Pinaceae in Puebla, México. Revista Mexicana de Biodiversidad, 88(1): 215-223. DOI: https://doi.org/10.1016/j.rmb.2017.01.028Links ]org/10.1016/j.rmb.2017.01.028" target="_blank">.
Rasal M., Troncos J., Lizano C., Parihuamán O. & Quevedo D. 2012. La vegetación terrestre del bosque montano de Lanchurán (Piura, Perú). Caldasia. 34(1): 1-24. DOI: Consultado en:https://revistas.unal.edu.co/index.php/cal/article/view/36419/38120Links ]unal.edu.co/index.php/cal/article/view/36419/38120" target="_blank">.
Rohlf F.J. 1990. Morphometrics. Annual Review of Ecology and Systematics, 21: 299-316. https://www.jstor.org/stable/2097027Links ]jstor.org/stable/2097027" target="_blank">.
Romero M. 2015. Rasgos morfológicos de frutos, semillas y embriones de Cinchona officinalis L. en el sur del Ecuador. Revista Ecuatoriana de Medicina y Ciencias Biológicas, REMCB. (36): 27-35. [ Links ]
Sarkinen T., Gonzáles P. & Knapp S. 2013. Distribution models and species discovery: the story of a new Solanum species from the Peruvian Andes. PhytoKeys. 31: 1–20. DOI: https://doi.org/10.3897/phytokeys.31.6312Links ]org/10.3897/phytokeys.31.6312" target="_blank">.
Schindelin J., Arganda-Carreras I. & Frise E. 2012, Fiji: an open-source platform for biological-image analysis. Nature Methods. 9(7): 676-682. DOI: https://doi.org/10.1038/nmeth.2019Links ]org/10.1038/nmeth.2019" target="_blank">.
Steele A.R. 1964. Flowers for the king - The expedition of Ruiz and Pavon and the Flora of Peru. Durham, N.C. : Duke University Press. [ Links ]
Tapia J. 2013. Estudio de factibilidad para la producción orgánica y comercialización de quina (Cinchona officinalis) en el Cantón de Loja. Tesis de licenciatura. Universidad San Francisco de Quito, Quito. [ Links ]
Weigend M. 2004. Additional observations on the biogeography of the Amotape-Huancabamba zone in Northern Perú: Defining the South-Eastern limits. Revista Peruana de Biología. 11(2): 127–134. [ Links ]
Weigend M., Cano A., Rodríguez E.R. & Breitkopf H. 2010. Four New Species of Ribes (Grossulariaceae), Primarily from the Amotape–Huancabamba Zone in Northern Peru. Novon: A Journal for Botanical Nomenclature. 20(2): 228–238. DOI: https://doi.org/10.3417/2008090Links ]org/10.3417/2008090" target="_blank">.
Weigend M., Rodríguez E. & Arana C. 2005. The relict forests of northwest Peru and southwest Ecuador. Revista Peruana de Biología. 12(2): 185–194. Consultado en: http://www.scielo.org.pe/scielo.php?pid=S1727-99332005000200004&script=sci_arttextLinks ]scielo.org.pe/scielo.php?pid=S1727-99332005000200004&script=sci_arttext" target="_blank">.
Zevallos P.A. 1989. Taxonomía, distribución geográfica y status del Género Cinchona en el Perú. Universidad Nacional Agraria La Molina, Lima-Perú [ Links ].
Presentado: 23/07/2018
Aceptado: 25/11/2019

