










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.14 n.2 Lima dic. 2007
José Luis Mena1,2, Martha Williams1, Carla Gazzolo1 y Francesca Montero1
1 Departamento de Biología, Facultad de Ciencias, Universidad Nacional Agraria La Molina, Lima, Perú
2 Apartado Postal 11-0205, Lince, Lima 14, Perú.
Resumen
En la costa y vertiente occidental ocurren los principales endemismos de mamíferos peruanos. Así mismo, a lo largo de la costa central, sobresale el único mamífero endémico del departamento de Lima, el ratón arrozalero de Zúñiga, Melanomys zunigae (Sanborn, 1949), registrado en las lomas de Atocongo y el cerro San Jerónimo por Enrique Zúñiga en 1942. Desde entonces se han realizado esporádicas evaluaciones, no necesariamente dirigidas a su búsqueda, aunque sin ningún registro. Nuestro estudio tuvo como objetivo la búsqueda de evidencias de la presencia de M. zunigae en las lomas de Lima, basados en un trampeo no exhaustivo y posteriormente en un análisis de regurgitos de la lechuza de los arenales (Athene cunicularia). Desafortunadamente no registramos ningún individuo de M. zunigae; sin embargo, encontramos evidencias que el impacto de la expansión urbana estaría afectando los ensamblajes de mamíferos pequeños en las lomas de Lima. Varios factores pudieron haber determinado la desaparición de M. zunigae. Entre estas se incluyen la destrucción del hábitat, invasión de especies exóticas y el aislamiento de las lomas.
Palabras clave: Melanomys zunigae, Mamíferos pequeños, Lomas, estado de conservación, Lima, Perú
Abstract
The coastline and the western slopes of the Andes are main sites of Peruvian endemic mammals. Along the central coast occurs the only endemic rat from Lima, Zuniga's dark rice rat, Melanomys zunigae (Sanborn, 1949). It was collected by Enrique Zuñiga in 1942 at Lomas de Atocongo and Cerro San Jerónimo. Since then, sporadic searches have been conducted, not necessarily targeted at finding it, but without records. Our study was aimed at finding evidence of its presence in the Loma formations around the city of Lima. Our methods were non-exhaustive trappings and the collection of burrowing owl (Athene cunicularia) pellets. Unfortunately no evidence of the presence of M. zunigae was found. However, we found evidence that the impact of Lima´s urban sprawl would be affecting small mammal assemblages at the Lomas. Several factors could have determined M. zunigae´s disappearance: habitat destruction, exotic species invasions and Lomas isolation.
Keywords: Melanomys zunigae, Small mammals, Lomas, conservation status, Lima, Peru
Introducción
La costa occidental de Sur América (5—30° Latitud Sur) está dominada por condiciones desérticas que forman un cinturón continúo e hiper-árido, interrumpido sólo por los ocasionales valles fluviales (Dillon, 2005). En estos desiertos ocurre la única vegetación no ribereña conocida como Lomas, que se caracteriza por presentar una cobertura vegetal producto de las precipitaciones esporádicas y de la humedad de las neblinas provenientes del litoral en la época de invierno (Ordoñez y Faustino, 1983; Rundel et al., 1991). Estas formaciones vegetales son aisladas y presentan un alto grado de endemismo (Dillon, 2005). En el Perú, las lomas y los valles de la vertiente occidental constituyen áreas de importancia para los mamíferos debido a la presencia de endemismos (Ascorra et al., 1996; Pacheco, 2002). Una de las especies más resaltantes en estas regiones es Melanomys zunigae (Sanborn, 1949), el único roedor endémico de las lomas del departamento de Lima. Esta especie fue colectada en las Lomas de Atocongo y el cerro San Jerónimo por Enrique Zúñiga en 1942 (Sanborn, 1949; Zuñiga, 1942). Desde entonces no se ha capturado ningún individuo; por el contrario, la destrucción y degradación de las formaciones de Lomas ha ido en aumento debido al avance urbano, la actividad minera, el pastoreo intensivo y la introducción de especies exóticas (Cano et al., 1999; Kunimoto et al., 2002). Melanomys zunigae se encuentra seriamente amenazado y desafortunadamente no se encuentra protegido en ninguna unidad de conservación (Pacheco, 2002). Esta especie fue una vez considerada dentro de la categoría de bajo riesgo y menor preocupación por la UICN (Baillie, 1996); sin embargo, la última categorización de especies amenazadas de fauna silvestre en Perú cambió su estado al de especie en peligro crítico (Decreto Supremo N° 034-2004-AG).
El principal objetivo de nuestro estudio fue determinar la existencia de poblaciones de M. zunigae en las lomas de Lima. Adicionalmente, se caracterizaron las ensamblados de mamíferos pequeños no voladores de las lomas de Lima, en relación con el impacto de las actividades humanas, como un indicador del estado de conservación de estos ecosistemas.
Materiales y métodos
Entre julio de 2005 y octubre de 2006 visitamos diez lomas ubicadas en el departamento de Lima (Tabla 1; Fig. 1). Las evaluaciones se realizaron principalmente en la época de Lomas (julio _ octubre). La cobertura vegetal durante las visitas a las lomas estuvo representada por especies arbóreas de Capparidaceae (Capparis prisca) y Fabaceae (Parkinsonia aculeata y Caesalpinia spinosa); entre las especies arbustivas resaltaron Euphorbiaceae (Croton spp.), Verbenaceae (Lantana spp.), Nolanaceae (Nolana spp.) Solanaceae (Nicotiana spp., Lycopersicon spp., Solanum spp.), Adiantaceae (Adiantum spp.) y Piperaceae (Peperomia spp.), Oxalidaceae (Oxalis sp.), entre otras. Entre agosto y noviembre del 2005 se completó una primera etapa de captura y recaptura, implementándose parcelas de siete columnas y siete filas, con estaciones-trampa distanciadas 7 m una de otra. Se usaron trampas de captura Sherman de tres tamaños: pequeñas (16,5 x 5 x 6 cm), medianas (23 x 7,5 x 9 cm) y grandes (30 x 7,5 x 9 cm). Los individuos capturados fueron pesados en una balanza y se les midió la longitud del cuerpo, cola, oreja y pata. Este método resultó ser muy poco efectivo para la captura y registro de la diversidad de mamíferos pequeños.
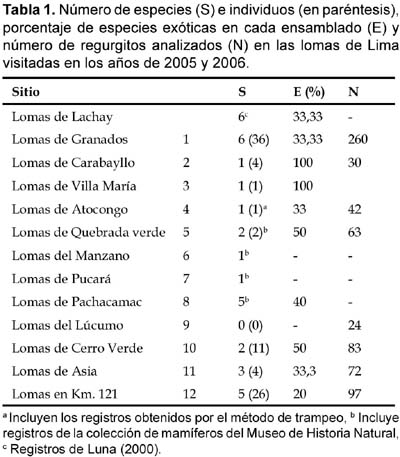

Debido a que el método de captura y recaptura resultó ineficiente, se optó por un método más eficaz. En este caso, el método de registro se basó en la búsqueda de regurgitos o egagrópilas de la lechuza de los arenales Athene cunicularia. Este método de registro ha demostrado ser exitoso para el inventario de mamíferos pequeños (Bonvicino y Bezerra, 2004; Hedrick et al., 1989; Luna, 2000; Torre et al., 2004). Las lomas fueron recorridas durante 4—6 horas cada una, tiempo que cubría la identificación de los regurgitaderos y la recolección de las egagrópilas. La mayor concentración de los regurgitaderos se localizó en las partes bajas de las lomas, tal y como se ha reportado previamente (Pulido, 1982), lo cual facilita su ubicación. Las egagrópilas fueron recogidas y guardadas en bolsas plásticas (25 x 4 cm). Posteriormente, las egagrópilas fueron examinadas al estereoscopio para la determinación de los restos presentes. Para este fin se tomó en cuenta los métodos descritos por Luna (2000). Para la determinación de las especies de mamíferos pequeños se consultó a Pearson (1958) y Luna (2000). Adicionalmente, para complementar el registro de especies, se revisó la colección de mamíferos del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, así como los estudios publicados anteriormente sobre la mastofauna de las lomas (Aguilar, 1985; Brack, 1974; Luna, 2000).
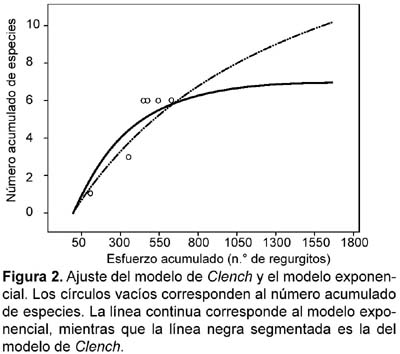
Para conocer si el esfuerzo de muestreo (número de regurgitos) fue adecuado, se usó las curvas de acumulación de especies (Colwell et al., 1995; Gotelli y Colwell, 2001). Una función de acumulación de especies es una curva no decreciente que representa el número acumulado esperado de especies diferentes encontradas dentro de un área geográfica como una función de la medida del esfuerzo (Díaz-Francés y Soberón, 2005). Para la predicción de la riqueza de especies de un sitio, considerando la curva de acumulación conocida, Soberón y Llorente (1993) sugirieron tres modelos: el logístico, el exponencial y el de Clench. Estos dos últimos modelos son útiles para describir situaciones en las cuales el área evaluada es pequeña, los taxones son bien conocidos, o los colectores acumulan experiencia, la cual incrementa la plausibilidad de detectar nuevas especies cuanto más tiempo se invierte en el campo. Estas características son las que describen la realidad de nuestro estudio, debido a que conocíamos cual fue el pool de especies de mamíferos pequeños que podían habitar en una Loma y además que las áreas estudiadas fueron relativamente pequeñas. Díaz-Francés y Soberón (2005) describieron en forma algorítmica, un análisis estadístico alternativo basado en un método probabilístico, que aporta rigor matemático a las ideas de Soberón y Llorente (1993) y mejora la estimación de los modelos mediante la incorporación del hecho que la varianza del error no es constante y que las observaciones están correlacionadas. Asimismo, permite conocer dentro de varios modelos, cuál es el que mejor se ajusta a los datos colectados en las lomas. En éste caso, el programa que se usó fue el Species accumulation functions, disponible en la dirección: http://cimat.mx/infophp?m=1&ind=5.
En cuanto a la caracterización de las lomas evaluadas, se tomó en cuenta principalmente la complejidad del hábitat y el impacto de la actividad humana. Al respecto, varios estudios han revelado que la complejidad vegetal en las lomas varía con la altitud. De esta manera, dependiendo del rango altitudinal de cada loma, se considera que de altitudes bajas a medias (200—500 m), hay una preponderancia de herbáceas y sub-arbustos; mientras que en altitudes medias a altas (500—800 m), hay una mayor presencia de árboles y arbustos (Ceroni, 2003; Ordoñez y Faustino, 1983; Torres y López, 1982), con una mayor abundancia en los fondos de quebradas (Velarde, 1983). Esta información permitió caracterizar la complejidad de hábitat de cada loma, usando como sinónimo de la misma el rango altitudinal. Asimismo, para caracterizar el grado de perturbación se consideró la distancia al centro poblado más cercano, cercanía a campos de cultivo, y el aislamiento respecto de otras lomas (basado en la distancia a la loma más cercana). Estas variables fueron relacionadas al número de especies en cada loma, usando una regresión por pasos (Ramsey y Schafer, 2002). Todas las variables fueron previamente transformadas por medio de la función logarítmica para equiparar las varianzas. Todas las pruebas estadísticas fueron realizadas con el programa S-plus (Lam, 1999).
Resultados
Presencia de Melanomys zunigae
Desafortunadamente, no encontramos ninguna evidencia de la presencia de M. zunigae. Con el método de trampeo únicamente se capturó 14 individuos de Phyllotis amicus Thomas, durante la visita a tres lomas (Cerro verde, Manzano, y Quebrada verde), con un esfuerzo total de 672 trampas-noche (2,08% de éxito). Sin embargo, de los 671 regurgitos de lechuza colectados en las 10 lomas visitadas (Tabla 1), se identificó un total de 85 individuos, correspondientes a un marsupial: Thylamys sp.; tres roedores nativos: Aegialomys xantheolus (Thomas) (Weksler et al., 2006), Phyllotis amicus (Thomas), y Phyllotis andium Thomas, y dos roedores exóticos: Mus musculus Linnaeus y Rattus rattus (Linnaeus). En general, son muy pocos los estudios publicados sobre mamíferos de las lomas de Lima, casi todos corresponden a estudios realizados en las Lomas de Lachay (Kunimoto et al., 2002; Luna, 2000; Ramírez & Arana, 2005), con una sola excepción, la cual corresponde al estudio de Zúñiga (1942), en las lomas de Atocongo. No obstante, existen varias evaluaciones que han sido realizadas en el contexto de estudios de impacto ambiental en el departamento de Lima, los cuales tampoco han tenido éxito en la captura de M. zunigae.
Los resultados del Species accumulation functions sugieren que los modelos de Clench y el exponencial son los más adecuados para los datos de los regurgitos (Fig. 2). El modelo de Clench predijo que es posible encontrar 18 especies. En este sentido, el método de regurgitos podría ser considerado como poco eficiente, debido a que según Clench, sólo registró el 33,3% del total de especies; sin embargo, el modelo exponencial predijo un total de siete especies. Adicionalmente, considerando las ecuaciones de ambos modelos (Soberón & Llorente, 1993), estimamos el número de regurgitos necesarios para registrar el 90% de las especies predichas por los modelos de Clench y exponencial, el cual fue de 24548 y 1066 regurgitos, respectivamente. En términos prácticos, el modelo de Clench no parece ser un buen estimador de la riqueza de especies para el método de regurgitos, debido a que su predicción resultó por demás exagerada. Es poco probable, esperar más de cinco especies de mamíferos pequeños nativos en las lomas de Lima, las cuales fueron registradas en este estudio, aunque Phyllotis limatus ha sido registrada también en bajas altitudes en Lima (Steppan, 1998). En todo caso, la riqueza de especies esperada a partir del modelo exponencial parece acercarse más al de la comunidad real. De esta forma, al añadir las dos especies exóticas reportadas en las lomas (Mus musculus y Rattus rattus) se completan las siete especies estimadas por este modelo. Por otro lado, es importante señalar que el desempeño de estos modelos o su evaluación, dependen en gran medida del conocimiento que se tiene sobre el pool actual (real) de las especies presentes (Díaz-Francés & Soberón, 2005; Soberón & Llorente, 1993). Este conocimiento fue determinante en nuestro estudio.
Diversidad y extinción
Los análisis de regresión revelaron interesantes asociaciones entre la riqueza de especies y la caracterización de las lomas. Se encontró una asociación significativa (r2= 0,54, F2,8= 4,71, P< 0,05) entre el número de especies y la distancia a centros poblados y el rango altitudinal de la loma (Número de especies = 3,84 + 0,21*Distancia — 0,73*Rango). Asimismo, encontramos una asociación significativa (r2= 0,92, F3,6= 22,05, P< 0,002) entre el porcentaje de exóticos presentes en las lomas con la distancia a centros poblados, aislamiento y rango altitudinal (Porcentaje de exóticos= 3,19 — 0,29*Distancia — 0,55*Aislamiento + 0.52*Rango).
Discusión
Presencia de Melanomys zunigae en las lomas de Lima
Estudios de regurgitos de lechuzas han permitido obtener registros complementarios para inventarios de diversidad (Bonvicino & Bezerra, 2004; Martin, 2003; Torre et al., 2004), actualización de los rangos de distribución (Martin, 2003; Pardiñas et al., 2003), o redescubrimiento de especies consideradas extintas (Dowler et al., 2000; Teta et al., 2006). En nuestro estudio, éste método resultó ser bastante efectivo para registrar los mamíferos pequeños presentes en las lomas, excepto para M. zunigae. Por otro lado, es importante mencionar que el registro de especies provenientes de líneas o parcelas de trampas pueden no reflejar la composición del ensamblaje de pequeños mamíferos, y en algunos casos la abundancia de ciertas especies pueden estar sub o sobre-representadas (Hedrick et al., 1989), por lo que es necesario el uso de métodos complementarios. Asimismo, el hecho que los roedores parecen constituir la principal fuente de alimento de Athene cunicularia (Luna, 2000; Pulido, 1982), califica al método de regurgitos como una alternativa adecuada para los objetivos del estudio. Es importante mencionar que el tamaño corporal de M. zunigae se encuentra dentro del rango de tamaño de las presas consumidas normalmente por A. cunicularia (Luna, 2000). Tales características validan la eficiencia del método, en la medida que logramos registrar el número de mamíferos pequeños nativos esperados de acuerdo a estudios previos en las lomas de Lima (Brack, 1974; Luna, 2000; Zuñiga, 1942).
Estimación de la diversidad y extinción
Debido a que no se encontraron evidencias de M. zunigae, buscamos identificar algunos factores que actualmente se encuentren influenciando la riqueza de especies de las lomas, con la finalidad de establecer qué pudo determinar la probable desaparición de esta especie. Los análisis de regresión revelan que por lo menos los factores de importancia que afectan la riqueza de especies, son la distancia a los centros poblados y la complejidad vegetal que existe en la zona de estudio, que en este caso es expresada en términos del rango altitudinal de la loma. Asimismo, sugieren que las especies exóticas constituyen el componente más importante de los ensamblajes de mamíferos pequeños en sitios cercanos a centros poblados (como era de esperarse). De hecho, las especies exóticas parecen ser las habitantes más conspicuas en las lomas más perturbadas y con un avanzado estado de aislamiento, como fue el caso de las lomas de Villa María y Carabayllo. Mus musculus es la especie con mayor número de registros tanto temporal como espacialmente (Tabla 2). En efecto, nuestro estudio reporta que esta especie exótica ha sido registrada en 10 lomas desde 1942, mientras que P. amicus (un roedor nativo) sólo ha sido registrado en nueve lomas. Todo lo contrario ocurre con M. zunigae que fue colectado solo en una loma en 1942.
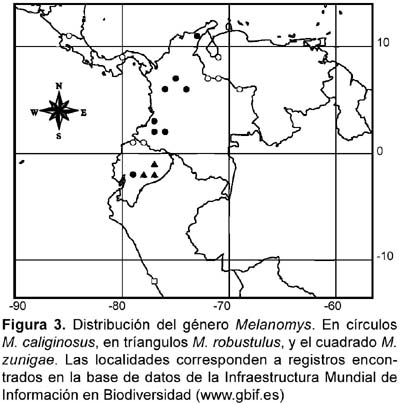
De acuerdo a lo reportado por Zúñiga (1942) y Sanborn (1949), si M. zunigae es confinado al ecosistema de lomas, no podría encontrarse más allá de los 8° S, ya que este es el límite norte de la distribución de las lomas (Cano et al., 1999; Rundel et al., 1991). En relación a esto es importante señalar que las otras especies del género Melanomys se encuentran distribuidas desde Centro América hasta el sur de Ecuador. Como se puede ver en la figura 3, M. zunigae constituye un grupo disjunto respecto de las otras especies del género, al igual que M. robustulus. Al parecer, el género está restringido a áreas boscosas en un rango altitudinal de 0 a 2500 m, desde Costa Rica a lo largo de la costa Pacífica y Caribe de Colombia, y la costa Pacífica y el sur oriente de Ecuador (Alberico et al., 2000; Allen, 1913; Musser & Carleton, 2005). Los registros de M. zunigae constituyen el límite sur de la distribución del género.
Varios argumentos han sido propuestos para declarar a una especie como extinta. Por ejemplo, de acuerdo a la Unión Internacional para la Conservación de la Naturaleza, la extinción se define cuando no hay duda razonable que el último individuo ha muerto (Rodrigues et al., 2006). Sin embargo, se puede concluir convencionalmente que una especie ha llegado a extinguirse si ella no ha sido registrada por un lapso de 50 años (Smith et al., 1993); dicho intervalo es totalmente arbitrario, ya que en ocasiones se ha redescubierto especies luego de 50 o más años desde su descripción (Arroyo-Cabrales et al., 2005; Percequillo et al., 2004). No obstante, en la actualidad se considera que un taxón está presumiblemente extinto cuando evaluaciones exhaustivas en hábitat conocidos o potenciales, en tiempos apropiados (diurno, estacional o anual), por todo su rango histórico han fracasado en el registro de un individuo (UICN, 2001, 2003).
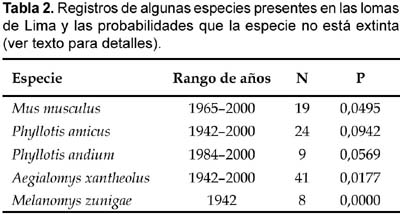
Desde la descripción de M. zunigae (Sanborn, 1949) han transcurrido 57 años de esporádicas evaluaciones, siendo la reportada en este caso la única llevada a cabo para buscar evidencias de la existencia del ratón arrozalero de Zúñiga. A pesar de la poca información que se tiene sobre esta especie, es posible estimar una probabilidad de extinción a partir de datos de avistamientos (e. g. registros de museos). En efecto, Burgman et al. (1995) propusieron que la probabilidad que una especie no llegue a estar extinta desde el último avistamiento realizado (p), puede ser calculada con la ecuación:
p = (Ce/CT)N
En donde Ce es el intervalo de tiempo entre el comienzo del período de observación y la última colecta, CT es el intervalo entre la primera y ultima observación (en unidades definidas tales como meses o años), y N es el número de veces en que la especie fue previamente observada. Aplicamos esta fórmula a los registros de algunas especies presentes en la colección de mamíferos del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos. Los resultados sugieren que respecto a M. zunigae su probabilidad de no extinción es prácticamente nula y que probablemente A. xantheolus, la otra especie de la tribu Oryzomini, podría tener algún grado de amenaza (ver Tabla 2). Es importante resaltar que la ecuación usada no asume que la colección de especimenes ha sido sistemática. Los supuestos representan más cercanamente un proceso en el cual las colecciones son enteramente oportunistas. De esta manera, a pesar de que estas estimaciones nos permiten conocer algunas tendencias, y llamar la atención de los conservacionistas (Burgman et al. 1995); no obstante, consideramos que estos resultados deben ser tomados con precaución.
Actualmente, una las grandes amenazas para la supervivencia de estas especies es la acelerada expansión urbana de la ciudad de Lima (Pacheco, 2002), la cual ha avanzado en desmedro de la superficie de Lomas, en especial en las lomas de Atocongo (localidad tipo) y las lomas aledañas (Fig. 1). En efecto, nuestro análisis de regresión revela que la cercanía a los centros urbanos influye negativamente en la riqueza de especies de mamíferos pequeños de las lomas, lo mismo que el aislamiento, y que estos rasgos por el contrario, favorecen la dominancia de especies exóticas. Es un hecho comprobado que las especies exóticas representan una de las principales causas de extinción de las especies silvestres (Primack et al., 2001; Sakai et al., 2001), sobre todo en islas (Courchamp et al., 2003). Posiblemente, debido a los rasgos aparentemente insulares de las formaciones de lomas, una combinación de invasión por exóticos y destrucción de hábitat pueden haber contribuido a una pauperización de las poblaciones de M. zunigae y otros mamíferos pequeños. Mus musculus parece haber encontrado un ambiente adecuado en las lomas, pues ha sido registrado en Lachay y Atiquipa (Arana et al., 2006; Kunimoto et al., 2002; Zeballos et al., 2000), y varias de las lomas que hemos evaluado. Inclusive, esta especie es favorecida enormemente durante eventos extraordinarios como El Niño, como ha sido el caso en las Lomas de Lachay en 1998 (J. L. Mena, in litt). En general, M. musculus es una de las especies exóticas con más éxito, pues ha colonizado prácticamente todo lugar en donde habita el ser humano y áreas silvestres aledañas.(Álvarez-Romero & Medellín, 2005; Jones et al., 2003).
Conservación de las lomas
La destrucción del hábitat parece haber sido un factor determinante en la desaparición de M. zunigae de las lomas de Lima. No obstante, el deterioro de las lomas data desde tiempo de la conquista con la tala desmedida de la vegetación arbórea (Rostworwoski, 2005). Como es conocido, la importancia de las precipitaciones en las lomas está ligada a la cobertura vegetal (Ordoñez & Faustino, 1983). Zúñiga (1942) señaló que las lomas de Atocongo tenían "una vegetación abundante que casi llegaba hasta la planicie, inclusive con pequeñas fuentes de agua que parecían durar todo el año". Asimismo, Ferreyra (1953) reportó que las precipitaciones en las Lomas de Atocongo inclusive superaba a la de las Lomas de Lachay (la única área natural protegida que conserva el ecosistema de Lomas). Durante la colecta científica de E. Zúñiga, no se registró ningún espécimen de M. musculus; inclusive el éxito de captura (44 especimenes en 325 trampas-noche, 13,5%) fue uno de los más altos para ecosistemas áridos de las lomas y desiertos del Pacífico (Arana et al., 2002; Kunimoto et al., 2002; Luna, 2000), donde por lo general, el éxito es realmente bajo (Jaksic et al., 1999). Estos hechos podrían evidenciar un gran deterioro del hábitat de M. zunigae, que no solo habría afectado a esta especie. En general, las formaciones de lomas están seriamente amenazadas. Las comunidades vegetales de las lomas, donde se cuentan varias especies endémicas (Dillon, 2005; Rundel et al., 1991), en algunos casos incluyen especies de plantas exóticas (Cano et al., 1999). En cuanto a los mamíferos pequeños, es necesaria una reflexión y acción seria para eliminar los impactos causados por las especies exóticas, además de proponer alternativas de desarrollo que no afecten la conservación de las lomas. En el caso del ratón arrozalero de Zúñiga, consideramos que es necesario apoyar nuestros resultados con evaluaciones continuas usando diversos métodos de captura, sobre todo en las lomas con mejor estado de conservación, lo cual podría contribuir a confirmar o refutar nuestros resultados, probablemente confirmando la extinción de un mamífero más de nuestra mastofauna.
Agradecimientos
Esta investigación fue financiada gracias a fondos obtenidos a través de la Iniciativa de Especies Amenazadas (IEA) que Conservación Internacional desarrolla en cooperación con socios locales de los países que conforman el Centro de Conservación de la Biodiversidad de los Andes Tropicales (CBC-Andes). En el Perú el Programa se desarrolla a través de las "Becas María Koepcke" coordinado por Conservación Internacional-CI Perú y la Asociación Peruana para la Conservación de la Naturaleza- APECO. Agradecemos a Víctor Pacheco por permitirnos revisar y consultar la base de datos y la colección de mamíferos del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos. Registros adicionales fueron obtenidos a través del Sitio Web de www.gbif.org. A todos los asistentes de campo que colaboraron en la investigación, a Sidney Novoa por su ayuda en la identificación de la flora de algunas de las lomas visitadas y a la familia Boyco por su hospitalidad en las cercanía de una de ellas. CI brindó su apoyo a través del préstamo de algunas trampas para el trabajo de campo, y WWF-Perú proporcionó información cartográfica relevante. A los revisores, que con sus observaciones contribuyeron al enriquecimiento de la información presentada.
Literatura Citada
Aguilar, P. G. 1985. Fauna de las lomas costeras del Perú. Boletín de Lima 41:17-28.
Alberico, M., A. Cadena, J. Hernández-Camacho & Y. Muñoz-Saba. 2000. Mamíferos (Synapsida: Theria) de Colombia. Biota Colombiana 1:43-75.
Álvarez-Romero, J. & R. A. Medellín. 2005. Mus musculus. Vertebrados superiores exóticos en México: diversidad, distribución y efectos potenciales. México. D.F.: Instituto de Ecología, Universidad Nacional Autónoma de México. Bases de datos SNIB-CONABIO. Proyecto U020.
Allen, J. A. 1913. Revision of the Melanomys group of American Muridae. Bulletin of the American Museum of Natural History 32:533-554.
Arana, M., O. Ramirez, S. Santa Maria, C. Kunimoto, R. Velarde, C. De La Cruz & M. L. Ruiz. 2002. Population density and reproduction of two Peruvian leaf-eared mice (Phyllotis spp.). Revista Chilena de Historia Natural 75:751-756.
Arana, M., M. L. Ruiz-Luna, S. S. Maria & O. Ramirez. 2006. Population fluctuations of the house mouse in a Peruvian loma and the functional response of burrowing owls. Austral Ecology 31:956-963.
Arroyo-Cabrales, J., E. K. Kalko, R. K. Laval, J. E. Maldonado, R. A. Medellín, O. J. Polaco & B. Rodríguez-Herrera. 2005. Rediscovery of the Mexican flat-headed bat Myotis planiceps (Vespertilionidae). Acta Chiropterologica 7:309-318.
Ascorra, C. F., D. M. Leo, L. O. Rodríguez & V. Pacheco. 1996. Áreas importantes para la conservación de los mamíferos en el Perú. En: L. O. Rodríguez, ed. Diversidad biológica del Perú: zonas prioritarias para su conservación, Proyecto Fanpe GTZ - INRENA, Lima. Pp. 71-78.
Baillie, J. 1996. Melanomys zunigae. En: IUCN 2006. IUCN Red List of Threatened Species <www.iucnredlist.org> Downloades on 14 October 2006.
Bonvicino, C. R. & A. M. R. Bezerra. 2004. Use of Regurgitated Pellets of Barn Owl (Tyto alba) for Inventoryng Small Mammals in the Cerrado of Central Brazil. Studies on Neotropical Fauna and Environment 38:1-5.
Brack, A. 1974. Los vertebrados de las lomas costeras del Perú. Anales científicos UNA La Molina 12:85-92.
Burgman, M. A., R. C. Grimson & S. Ferson. 1995. Inferring threat from scientific collections. Conservation Biology 9:923-928.
Cano, A., J. Roque, M. Arakaki, C. Arana, M. La Torre, N. Llerena & N. Refulio. 1999. Diversidad florística de las lomas de Lachay (Lima) durante el evento "El Niño 1997-98". Revista Peruana de Biología Vol. extraordinario:125-132.
Ceroni, A. 2003. Distribución altitudinal de las especies vegetales más frecuentes de las lomas de Atiquipa de la costa desértica sur del Perú (Arequipa). Zonas Áridas 7:177-197.
Colwell, R. K., J. A. Coddington & D. L. Hawksworth. 1995. Estimating terrestrial biodiversity through extrapolation. En: Biodiversity: measurement and estimation (D. L. Hawkswoth, Ed.). Chapman and Hall and The Royal Society, London. Pp. 101-118.
Courchamp, F., J. L. Chapuis & M. Pascal. 2003. Mammal invaders on islands: impact, control and control impact. Biological Reviews 78:347-383.
Díaz-Francés, E. & J. Soberón. 2005. Statistical estimation and model selection of species-accumulation functions. Conservation Biology 19:569-573.
Dillon, M. O. 2005. Solanaceae of the Lomas formations of Coastal Peru and Chile. En: V. Hollowell, T. Keating, W. Lewis & T. Croat (eds.) "A festschrift for William G. D'Arcy: The Legacy of a Taxonomist". Monographs in Systematic Botany from the Missouri Botanical Garden, Missouri Botanical Garden Press, pp. 131-155.
Dowler, R. C., D. S. Carrol & C. W. Edwards. 2000. Rediscovery of rodents (Genus Nesoryzomys) considered extinct in the Galápagos islands. Oryx 34:109-118.
Ferreyra, R. 1953. Comunidades vegetales de algunas lomas costaneras del Perú. Boletín Est. Exper. La Molina 53.
Gotelli, N. J. & R. K. Colwell. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology letters 4:379-391.
Hedrick, P. W., M. S. Gaines & M. L. Johnson. 1989. Owl feeding habits on small mammals. Occasional Papers of the Museum of Natural History. The University of Kansas Lawrence, Kansas 133:1-7.
Jaksic, F. M., J. C. Torres-Mura, C. Cornelius & P. A. Marquet. 1999. Small mammals of the Atacama Desert (Chile). Journal of Arid Environments 42:129-135.
Jones, A. G., S. L. Chown & K. J. Gaston. 2003. Introduced house mice as a conservation concern on Gough Island. Biodiversity and Conservation 12:2107-2119.
Kunimoto, C., C. De La Cruz, M. Arana & O. E. Ramírez. 2002. Observaciones sobre la ecología poblacional del ratón doméstico en Lachay, Perú. Bulletin de I'Institut français d'études andines 31:323-328.
Lam, L. 1999. An introduction to S-PLUS for Windows. Amsterdam: Candiensten.
Luna, L. 2000.Dinámica poblacional de los pequeños mamíferos en la Reserva Nacional Lomas de Lachay, Lima, y su relación al evento "El Niño Oscilación Sur". Tesis para optar el título profesional de biólogo con mención en zoología, Universidad Nacional Mayor de San Marcos, Facultad de Ciencias Biológicas.
Martin, G. 2003. Nuevas localidades para marsupiales patagónicos (Didelphimorphia y Microbiotheria) en el noroeste de la provincia del Chubut, Argentina. Mastozoología Neotropical 10:148-153.
Musser, G. G. & M. D. Carleton. 2005. Superfamilia Muroidea. En: D. E. Wilson & D. M. Reeder (eds.) "Mammal species of the World: a taxonomic and geographic reference". Johns Hopkins University Press, pp. 894-1531.
Ordoñez, J. L. & J. Faustino. 1983. Evaluación del potencial hídrico en lomas costeras del Perú (Lomas de Lachay-Iguanil). Zonas Áridas 3:29-42.
Pacheco, V. 2002. Mamíferos de Perú. En: G. Ceballos & J. A. Simonetti (eds.) Diversidad y Conservación de los Mamíferos Neotropicales, CONABIO-UNAM, México D. F., pp. 503-549.
Pardiñas, U. F. J., P. Teta, S. Cirignoli & D. H. Podest 2003. Micromamíferos (Didelphimorphia y Rodentia) de Norpatagonia extra Andina, Argentina: taxonomia alfa y biogeografia. Mastozoología Neotropical 10:69-113.
Pearson, O. P. 1958. A taxonomic revision of the rodent genus Phyllotis University of California Publications. Zoology 56:391-495.
Percequillo, A. R., P. R. Goncalves & J. A. De Oliveira. 2004. The rediscovery of Rhagomys rufescens (Thomas, 1886), with a morphological redescription and comments on its systematic relationships based on morphological and molecular (cytochrome b) characters. Mammalian Biology 69:238-257.
Primack, R., R. Rozzi, P. Feinsinger, R. Dirzo, F. Massardo, R. Primack, R. Rozzi, P. Feinsinger, R. Dirzo & F. Massardo. 2001. Fundamentos de conservación biológica: perspectivas Latinoamericanas. México D. F.: Fondo de cultura económica.
Pulido, V. M. 1982. La lechuza de los arenales de las lomas de Lachay, Lima-Perú. Zonas Áridas 2:75-78.
Ramírez, O. E. & M. Arana. 2005. Albinism in the Andean leaf-eared mouse, Phyllotis andium (Rodentia, Cricetidae). Mastozoología Neotropical 12:269-270.
Ramsey, F. L. & D. W. Schafer. 2002. The statistical sleuth: A course in methods of data analysis. Pacific Grove: Duxbury-Thomson Learning.
Rodrigues, A. S. L., J. D. Pilgrim, J. F. Lamoreux, M. Hoffmann & T. M. Brooks. 2006. The value of the IUCN red list for conservation. Trends in Ecology & Evolution 21:71-76.
Rostworowski, M. 2005. Ensayos de historia andina I. Lima: Instituto de Estudios Peruanos.
Rundel, P. W., M. O. Dillon, B. Palma, H. A. Mooney, S. L. Gulmon & J. R. Ehleringer. 1991. The phytogeography and ecology of the coastal Atacama and Peruvian deserts. Aliso 13:1-49.
Sakai, A. K., F. W. Allendorf, J. S. Holt, D. M. Lodge, J. Molofsky, K. A. With, S. Baughman, R. J. Cabin, J. E. Cohen, N. C. Ellstrand, D. E. Mccauley, P. Oïneil, I. M. Parker, J. N. Thompson & S. G. Weller. 2001. The population biology of invasive species. Annual Review of Ecology and Systematics 32:305-332.
Sanborn, C. C. 1949. A new species of rice rat (Oryzomys) from the coast of Perú. Publicaciones del Museo de Historia Natural "Javier Prado" UNMSM. Serie A. Zoología 3:1-4.
Smith, F. D. M., R. M. May, R. Pellew, T. H. Johnson & K. R. Walter. 1993. How much do we know about the current extinction rate? Trends in Ecology & Evolution 8:375-378.
Soberón, J. M. & J. Llorente. 1993. The use of species accumulation functions for the prediction of species richness. Conservation Biology 7:480-488.
Steppan, S. J. 1998. Phylogenetic relationships and species limits within Phyllotis (rodentia: Sigmodontinae): Concordance between mtDNA sequence and morphology. Journal of Mammalogy 79:573-593.
Teta, P., U. F. J. Pardiñas & G. D'Elia. 2006. Rediscovery of Chacodelphys: a South American marsupial genus previously known from a single specimen. Mammalian Biology 71:309-314.
Torre, I., A. Arrizabalaga & C. Flaquer. 2004. Three methods for assessing richness and composition of small mammal communities. Journal of Mammalogy 85:524-530.
Torres, J. & C. López. 1982. Estudio bioecológico de la Loma Paloma: 1. Flora y Vegetación. Zonas Áridas 2:61-66.
UICN. 2001. Categorías y criterios de la lista roja de la UICN: Versión 3.1. Comisión de supervivencia de especies de la UICN. Gland, Suiza y Cambridge, Reino Unido UICN.
UICN. 2003. Directrices para emplear los criterios de la Lista Roja de la UICN a nivel regional: Version 3.0. Comisión de Supervivencia de Especies de la UICN. Gland, Suiza y Cambridge, Reino Unido: UICN.
Velarde, D. 1983. Evaluación de la fauna de vertebrados de las Lomas de Iguanil. Zonas Áridas 3:83-89.
Weksler, M., A. R. Percequillo & R. S. Voss. 2006. Ten new genera of Oryzomyine rodents (Cricetidae: Sigmodontinae). American Museum Novitates 3537:1-29.
Zeballos, H., L. Villegas, R. Gutiérrez, K. Caballero & P. Jiménez. 2000. Vertebrados de las Lomas de Atiquipa y Mejía, sur del Perú. Revista de Ecología Latinoamericana 7:11-18.
Zuñiga, E. 1942. Observaciones ecológicas sobre los mamíferos de las lomas. Boletín del Museo de Historia Natural "Javier Prado" 22-23:392-399.
Correspondencia
1 Departamento de Biología, Facultad de Ciencias, Universidad Nacional Agraria La Molina, Av. La Molina s/n, Lima, Perú
2 Apartado Postal 11-0205, Lince, Lima 14, Perú.
Email José Luis Mena: jlmena@viabcp.com
Presentado: 01/02/2007
Aceptado: 02/11/2007
