











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
En la región neotropical, por encima del límite superior de los bosques andinos de Ecuador, Colombia, Venezuela, Perú, Panamá y Costa Rica, se extiende el bioma de páramo que alberga una enorme y singular diversidad biológica, por lo cual, se considera el ecosistema de alta montaña más biodiverso en el planeta (Diazgranados & Castellanos 2021).
Colombia posee la mitad de la superficie global de los páramos, teniendo además la mayor riqueza con 90 especies reconocidas de frailejones (Diazgranados & Castellanos 2021), individuos que se identifican como las plantas más emblemáticas que representan el páramo. No obstante, los páramos son ecosistemas vulnerables que enfrentan grandes amenazas por la ganadería extensiva, el aumento de la frontera agrícola, el turismo no controlado, las especies invasoras y el cambio climático que a futuro puede afectar la presencia y el mantenimiento de diversas poblaciones de frailejones (Rivera & Rodríguez 2011, Díazgranados 2015).
Los frailejones producen resinas de aromas agradables que se utilizaban como incienso y en la fabricación de jabones (Baruch 1979, García- Barriga 1992). También, se han usado para tratar el reumatismo (Acosta-Solís 1937b citado por De la Torre et al., 2006), enfermedades respiratorias (Báez et al. (1999), problemas del sistema circulatorio, embarazo, parto y otitis (Bussmann et al. 2018).
De las especies de Espeletia y Espeletiopsis se han aislado e identificado diferentes compuestos químicos, entre ellos, diterpenos, triterpenos, sesquiterpenlactonas, flavonoides y derivados del ácido cafeico. A partir de la resina de Espeletia grandiflora se identificaron los diterpenos: ent-kaur-16-eno, ent-kaur-16-en-19-al, ent-kaur-16-en-19-ol, ácido kaurenoico, ácido grandiflorolico y ácido grandiflorenico (Piozzi et al. 1968, 1971 citado por Padilla-González et al. 2017). De las hojas de E. tunjana se aislaron el ácido kaur-9(11),16-dien-19-oico, ent-kaur-16-en-19-ol y acetato de longipilina (Torrenegra et al. 1994, Torrenegra & Téllez 1995). Del extracto etéreo de las hojas e inflorescencias de E. killipii se obtuvieron quercetina, 3-metoxiquercetina, ácido kaur-9(11),16-dien-19-oico y kaur-16-en-19-ol, Polimatina B, acetato de longipilina, friedelina, sitosterol y estigmasterol (Torrenegra et al. 1994). En las hojas de E. argentea y en E. barclayana se identificó un triterpeno tipo cicloartano (Tellez et al. 1998). De las hojas de E. semiglobulata se aislaron ácido kaurénico y kaurenal (Aparizio et al., 2013). De E. barclayana se han reportado el ácido 5-O-(E)-caffeoilquínico, ácido 1,3-di-O-(E)-caffeoilquínico, ácido 1,5-di-O-(E)- caffeoilquinico, ácido 3,4-di-O-(E)-caffeoilquinico y 3-O-metilquercetina-7-O-β-glucopiranosido (Padilla-Gonzalez et al., 2017).
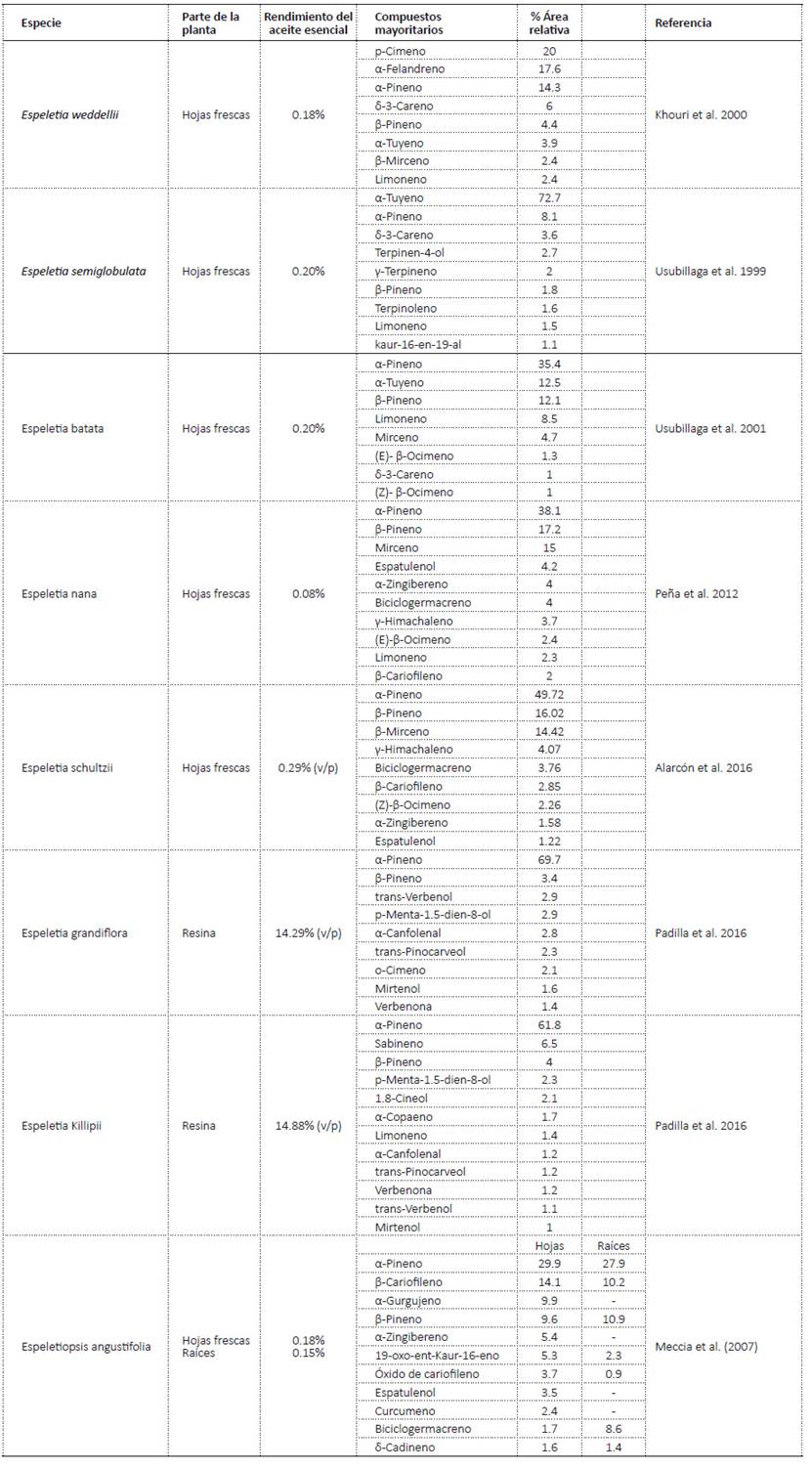
Los estudios de composición de los aceites esenciales de especies de los géneros Espeletia y Espeletiopsis han reportado entre los compuestos mayoritarios monoterpenos y sesquiterpenos (Tabla 1). Los aceites esenciales de Espeletia weddellii, Espeletia semiglobulata, Espeletia batata están constituidos mayoritariamente por monoterpenos tipo hidrocarburo, entre los que se destacan: p-Cimeno, α-Felandreno, α-Tuyeno, α-Pineno, β-Pineno, β-Mirceno y Sabineno (Usubillaga et al. 1999; Usubillaga et al. 2001; Khouri et al. 2000; Alarcón et al. 2016: Padilla et al. 2016). En los aceites esenciales de Espeletia nana, Espeletia schultzii se destaca la presencia mayoritaria de sesquiterpenos y sesquiterpenoides. Mientras que, en el aceite esencial de Espeletia grandiflora (Padilla et al., 2016) se reportan entre los mayoritarios monoterpenoides. De otra parte, en el aceite esencial de Espeletia semiglobulata se identificaron, además, diterpenos de tipo kaurano (Aparicio et al. 2013).
Tabla 1 Composición química de aceites esenciales de especies de frailejones del género Espeletia y Espeletiopsis angustifolia.
Algunos aceites esenciales de especies de Espeletia y Espeletiopsis han mostrado actividad al inhibir el crecimiento de bacterias Gram positivas. Los aceites esenciales de Espeletia schultzii, E. nana y Espeletiopsis angustifolia fueron activos frente a Staphylococcus aureus y Enterococcus faecalis (Alarcón et al. 2016; Peña et al. 2012; Meccia et al. 2007). Acosta y Torrenegra (2016) encontraron que el exudado de las hojas de E. killipii tiene efecto tóxico sobre larvas de lepidóptera quienes le atribuyeron el efecto a la presencia del ácido grandiflorénico, el cual fue mayor en hojas enfermas indicado un posible efecto de defensa de dicho compuesto contra el herbívoro.
En la presente investigación se estableció la composición química de los aceites esenciales de las hojas de cinco especies de frailejón (Espeletia argentea, Espeletia barclayana, Espeletia killipii, Espeletia summapacis y Espeletiopsis corymbosa) y se estudió su potencial antimicrobiano frente a hongos (Fusarium sp., Chaetomium sp., Botrytis sp., Penicillium sp., Alternaria sp.), Staphylococcus aureus (ATCC 25923) y Candida sp. con el propósito de ampliar el conocimiento en cuanto a la función biológica que cumplen los compuestos químicos frente al efecto de los microorganismos. Estas son especies que en su mayoría no cuentan con estudios relacionados con su composición química por lo cual, resulta de interés conocer sobre su diversidad química para profundizar y entender la importancia e incidencia que tienen los efectos de estos compuestos en el equilibrio de los ecosistemas naturales.
Material y métodos
Material vegetal. Las colectas del material vegetal se realizaron en cuatro Páramos ubicados alrededor de la ciudad de Bogotá D.C. entre 2016 y 2020. (1) Páramo de San Francisco: donde se colectaron Espeletia argentea Humb. & Bonpl. y Espeletiopsis corymbosa (Humb. & Bonpl.) Cuatrec. (4°10’55.2’’N, 74°13’51.4’W / 4°34’32.0’’N, 74°00’11.7’’W). (2) Páramo el Tablazo donde se colectó Espeletia barclayana Cuatrec. (5°00’41.2’’ N, 74°12’12.7’’W), (3) Páramo de Pasquilla donde se colectó Espeletia Killipii Cuatrec. (5°00’41.2’’N, 74°12’12.7’’W) y el (4) Páramo de Sumapaz donde se colectó Espeletia summapacis Cuatrec. (4°16’26.2’’N, 74°12’08.7’’W). La identificación taxonómica fue realizada por el biólogo Carlos Iván Suárez y un exicado que representa cada especie estudiada reposa en el Herbario del Jardín Botánico José Celestino Mutis (JBB22660, JBB14593, JBB22499, JBB23299, JBB13571).
Obtención de los aceites esenciales. Los aceites esenciales fueron obtenidos a partir de las hojas frescas por medio de hidrodestilación utilizando una trampa tipo Clevenger. La extracción se dejó por 3 h. Los aceites esenciales se secaron con sulfato de sodio anhidro y se conservaron a 4°C en viales de vidrio. Los rendimientos de las extracciones fueron calculados con el peso en fresco del material vegetal.
Análisis de los aceites esenciales por GC/MS. El aceite esencial de E. summapacis fue analizado en un cromatógrafo de gases 7890A (Agilent) acoplado a un espectrómetro de masas 5975C TAD (Agilent), se analizó en dos columnas capilares, VF-5MS (50 m x 200 μm x 0.33 μm) y CP- Wax 52 CB (50 m x 200 μm x 0.20 μm). El programa de temperatura en el horno inició a 50°C (2 min), se incrementó a 8°C/min hasta 280°C y luego a 15°C/min hasta 320°C (8 min). La temperatura del puerto de inyección fue 280°C. 2 µL de una dilución de la muestra en Acetonitrilo (Merck) fueron inyectados con una relación de Split 20:1. Se utilizó Helio como gas de arrastre a 1 mL/min. La temperatura de la línea de transferencia fue de 280°C.
Los aceites esenciales de Espeletia argentea, E. barclayana, E. killipii, y Espeletiopsis corymbosa fueron analizados en un cromatógrafo de gases Agilent Technologies 6890A acoplado a un detector selectivo de masas (MSD, AT 5973N) operado en modo de barrido completo de radiofrecuencia (full scan). Se realizaron corridas en dos columnas, DB-5MS (60 m × 0.25 mm × 0.25 µm) y DB-WAX (60 m × 0.25 mm × 0.25 µm), 2 µL de una dilución de cada aceite se inyectó en el instrumento en modo Split (30:1). Los índices de retención fueron determinados con una mezcla certificada de hidrocarburos C6-C25 (AccuStandard, New Haven, CT). La identificación tentativa de los compuestos se realizó con base en la comparación de los espectros de masas (EI, 70 eV), con las bases de datos de Adams, Wiley y NIST.
Evaluación de la actividad antimicrobiana. Los aceites esenciales se evaluaron frente a Staphylococcus aureus (ATCC 25923) y Candida sp. mediante la técnica de difusión en pozos. La bacteria se aisló en agar nutritivo y la levadura se aisló en agar de papa dextrosa (PDA), éstas fueron incubadas a 37 ± 2°C por 24 h. Posteriormente, se tomaron de 2 a 5 colonias con ayuda de un asa redonda y se suspendieron en solución salina estéril (0.85% p/v) hasta alcanzar la turbidez comparable con el patrón N° 0.5 de McFarland. Para el ensayo de actividad antimicrobiana, se llevó a cabo una siembra masiva en agar Mueller Hinton y PDA para Candida sp.), disponiendo 100 μL del inóculo. Luego, se realizaron pozos de 6 mm de diámetro y en cada uno se depositaron 20 μL de las muestras de aceite esencial al 3% diluidas en dimetilsulfóxido (DMSO). Como controles positivos se usaron cloranfenicol (100 μg/mL) para los ensayos con S. aureus y clotrimazol (100 μg/mL) para Candida sp. Se usó DMSO como control negativo. Finalmente, las cajas Petri se incubaron a 37 ± 2°C por 24 horas (Alvarado et al. 2010). Los ensayos se realizaron por triplicado.
 Evaluación actividad antifúngica. Inicialmente, se realizó la evaluación de los aceites esenciales de cuatro especies (Espeletia argentea, E. barclayana, E. killipii, y Espeletiopsis corymbosa) mediante la prueba de enfrentamiento, basado en el estudio de Quiroz et al. (2008), con modificaciones, frente a cepas de Fusarium spp. y Chaetomium spp. aisladas de plantas de ecosistemas altoandinos que forman parte del banco de microorganismos del Jardín Botánico de Bogotá José Celestino Mutis (JBBJCM). Los hongos aislados en agar PDA fueron verificados mediante la prueba de tinción con azul de lactofenol. Las pruebas se realizaron a los 7 días de la siembra del microorganismo en agar PDA, tomando un bocado de 6 mm de diámetro de agar cubierto con la especie fúngica, la cual se dispuso en el centro de la caja Petri y a dos (2) cm de distancia se realizaron los pozos de 6 mm en los cuales se colocaron 15 μL del aceite esencial. Las cajas Petri se incubaron a 27 °C y se realizaron lecturas a las 72, 96 y 168 h, el crecimiento del microorganismo se monitoreó en el tiempo hasta cuando la caja del control negativo estuvo cubierta en su totalidad por el microorganismo. El Porcentaje del Crecimiento del Hongo (PCH) se calculó de acuerdo con lo reportado por Salazar et al. (2012) en Arce-Araya et al. (2019).
Evaluación actividad antifúngica. Inicialmente, se realizó la evaluación de los aceites esenciales de cuatro especies (Espeletia argentea, E. barclayana, E. killipii, y Espeletiopsis corymbosa) mediante la prueba de enfrentamiento, basado en el estudio de Quiroz et al. (2008), con modificaciones, frente a cepas de Fusarium spp. y Chaetomium spp. aisladas de plantas de ecosistemas altoandinos que forman parte del banco de microorganismos del Jardín Botánico de Bogotá José Celestino Mutis (JBBJCM). Los hongos aislados en agar PDA fueron verificados mediante la prueba de tinción con azul de lactofenol. Las pruebas se realizaron a los 7 días de la siembra del microorganismo en agar PDA, tomando un bocado de 6 mm de diámetro de agar cubierto con la especie fúngica, la cual se dispuso en el centro de la caja Petri y a dos (2) cm de distancia se realizaron los pozos de 6 mm en los cuales se colocaron 15 μL del aceite esencial. Las cajas Petri se incubaron a 27 °C y se realizaron lecturas a las 72, 96 y 168 h, el crecimiento del microorganismo se monitoreó en el tiempo hasta cuando la caja del control negativo estuvo cubierta en su totalidad por el microorganismo. El Porcentaje del Crecimiento del Hongo (PCH) se calculó de acuerdo con lo reportado por Salazar et al. (2012) en Arce-Araya et al. (2019).
Teniendo en cuenta que el Porcentaje de Inhibición de Crecimiento Micelial (PICM) es el inverso del crecimiento radial, éste se calculó tomando el promedio del crecimiento del hongo y empleando la siguiente fórmula
%P1CM = 100- %PCH
Como control positivo se usó Benomil (1 mg/mL), las medidas se tomaron con calibrador digital pie de rey. Posteriormente con los aceites de E. corymbosa y E. argentea se profundizó en el estudio ya que se tenía disponibilidad de estos. Las demás especies no se pudieron evaluar en la segunda fase, debido a que por restricciones derivadas del Covid-19, no fue posible hacer un segundo muestreo a las zonas de colecta por aislamiento y cierre de fronteras rurales.
Análisis de datos. Se evaluó la normalidad mediante la prueba de Shapiro-Wilks (modificado) (p≤0.05); con las variables que no cumplieron el supuesto de normalidad se hizo un análisis de Kruskal Wallis, y con las que sí lo cumplieron, se realizó un análisis de varianza, ANOVA usando la prueba de Tukey (p > 0.05) para determinar diferencias entre tratamientos para cada uno de los días de evaluación. Todos los análisis se realizaron en el programa InfoStat Versión 2017 (Di Rienzo et al. 2008).
Resultados y discusión
Extracción de los aceites esenciales. Los aceites esenciales obtenidos presentaron olor característico, incoloros a excepción del aceite esencial de E. argentea que presentó color amarillo claro. Se determinaron los rendimientos de extracción para los cinco aceites esenciales, E. argentea (0.11%), E. barclayana (0.19%), E. corymbosa (0.07%), E. killipii (0.38%) y E. summapacis (0.81%). Los rendimientos reportados por Padilla et al. (2016) de la hidrodestilación de las resinas fueron de 14.29% y 14.88% para E. grandiflora y E. killipii, respectivamente. El rendimiento reportado por Alarcón et al. (2016) para el aceite esencial de E. schultzii fue de 0.29%, porcentaje que se encuentra dentro del rango de lo establecido en este estudio.
Caracterización química de los aceites esenciales.Los aceites esenciales de las especies de frailejón analizados están constituidos principalmente por monoterpenos tipo hidrocarburo (M) (63.4% hasta el 89.7%), en segundo lugar, se identificaron los sesquiterpenos (S), estos se encuentran entre un 20 y 21% en los aceites esenciales de E. argentea y E. barclayana. En menor proporción se encontraron monoterpenoides (MO) y sesquiterpenoides (SO). En el aceite esencial de E. argentea, el 4.2% correspondió a compuestos diterpénicos. Algunos de los compuestos identificados en estas muestras habían sido reportados para otras especies de Espeletia y en Espeletiopsis angustifolia (Khouri et al. 2000; Usubillaga et al. 1999; Usubillaga et al. 2001; Peña et al. 2012; Alarcón et al. 2016; Padilla et al. 2016).
El compuesto mayoritario en los aceites esenciales analizados fue el α-Pineno que se encontró entre 33.6% en E. killipii y 53.2% en E. barclayana. Otros monoterpenos abundantes fueron: β-Pineno, Sabineno, α-Felandreno, β-Felandreno. Los aceites esenciales de E. killipii y E. summapacis son, principalmente, monoterpénicos con 83.9% y 90.0%, respectivamente. Estos dos aceites esenciales son similares en su composición por la presencia de Sabineno, β- Pineno, α-Felandreno y β-Felandreno. La composición determinada para el aceite esencial de E. killipii en este estudio es semejante a lo reportado por Padilla et al. (2016) para un espécimen colectado en el Páramo de Sumapaz en 2015.
De otra parte, entre los compuestos sesquiterpénicos identificados en los aceites esenciales se destaca el Germacreno D presente en E. argentea, E. barclayana y E. corymbosa (12.3%, 5.9%, 2.3%, respectivamente), el Aristoleno en E. argentea (4.9%) y E. barclayana (4.4%) y el trans-β-Cariofileno en E. argentea, E. barclayana y E. corymbosa (1.9%, 1.1%, 2.2%, respectivamente). En menor proporción se identificaron el δ-Selineno, δ-Cadineno y α-Copaeno.
Se observó que los aceites esenciales de las cinco especies comparten similitudes en su composición pero con diferencias en las proporciones. Para evaluar qué tan distantes se encuentran las muestras entre sí, se hizo un análisis clúster utilizando el método de Ward con una distancia euclideana cuadrada. Este análisis se realizó con el paquete estadístico IBM: SPSS Statistics 22, donde las variables fueron las proporciones de los compuestos de los aceites esenciales. El Dendograma obtenido (Fig. 1) muestra las distancias entre las muestras de acuerdo con sus similitudes en su composición. En la gráfica, se observan varios grupos, a una distancia de 120, se identifican 3 grupos, uno corresponde a E. argentea, el segundo a E. barclayana y el tercero comprende a E. killipii, E. summapacis y E. corymbosa. Dentro de este tercer grupo se observa que hay mayor similitud entre la composición de los aceites esenciales de E. Killipii y E. summapacis. A pesar de que E. corymbosa hace parte de otro género botánico, en la química de su aceite esencial hay mayor relación que la que se observa entre las especies del género Espeletia. Además, la muestra del aceite esencial de E. argentea es la más alejada de las otras muestras. Este análisis es una aproximación desde los metabolitos secundarios presentes en los aceites esenciales que podrían ser correlacionadas con información genética como herramienta para la comprensión de las relaciones filogenéticas de los frailejones.
Tabla 2 Composición química de los aceites esenciales de Espeletia argentea, E. barclayana, E. Killipii, E. summapacis y Espeletiopsis corymbosa.
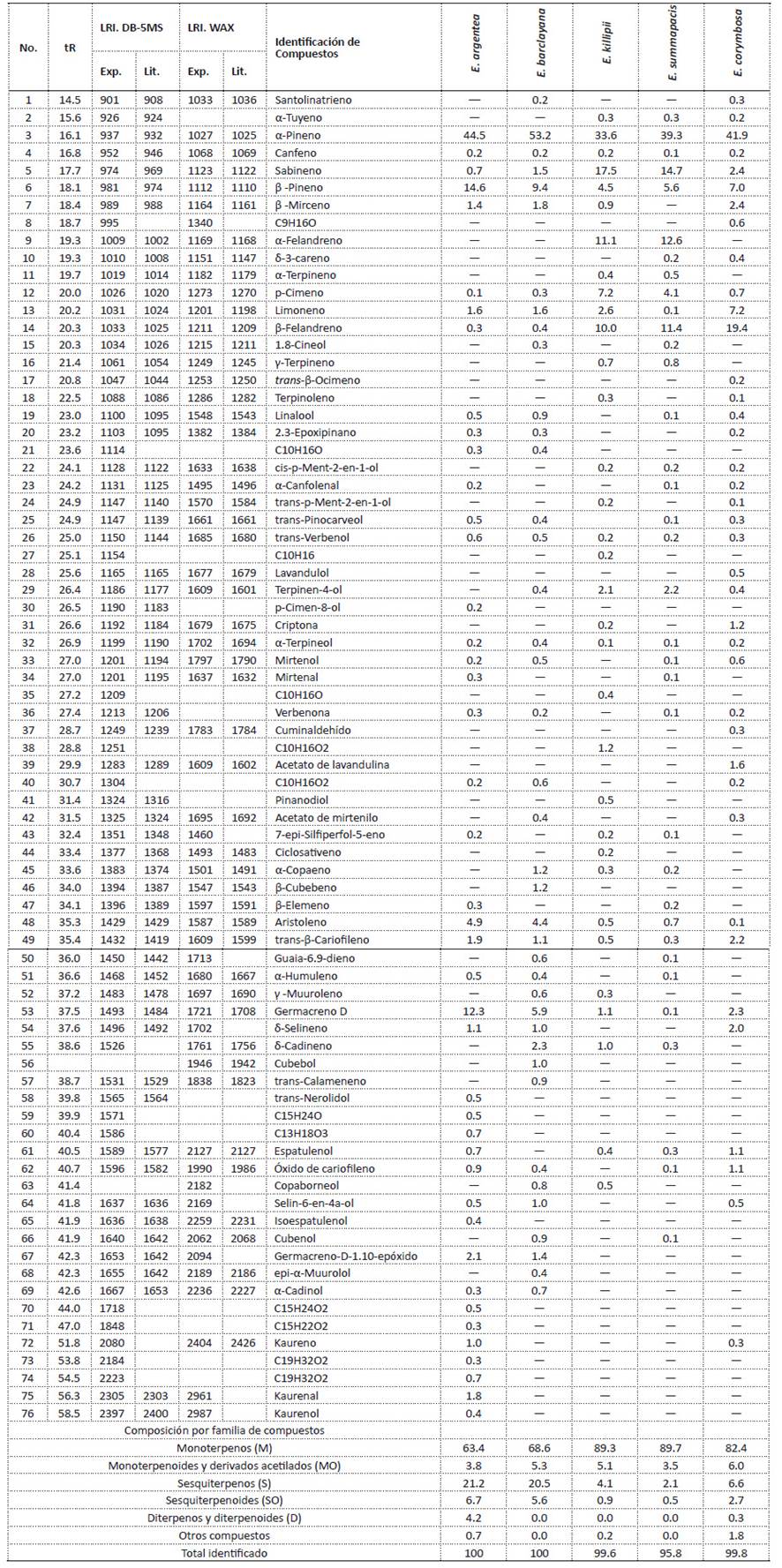
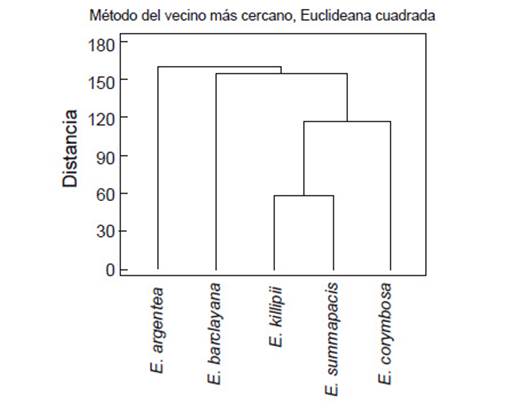
Figura 1 Dendograma del análisis clúster (SPSS) de la composición de los aceites esenciales E. argentea, E. barclayana, E. Killipii, E. summapacis y E. corymbosa.
Actividad antimicrobiana. Los aceites esenciales analizados no presentaron inhibición frente a Candida sp, estos resultados se relacionan con los obtenidos por Alarcón et al. 2016, quienes evaluaron la efectividad del aceite esencial de E. schiltzii, frente a dos cepas de Candida y no encontraron resultados positivos.
De los aceites evaluados los de E. killipii y E. corymbosa, presentaron una leve inhibición 11.70% y 5.04% frente S. aureus, respectivamente. La evaluación de la actividad biológica de algunas especies de Espeletiinae, se ha centrado en la evaluación frente a baterías patógenas, mostrando una mayor inhibición frente a microorganismos Gram positivos como S. aureus, en concentraciones de 1000 a 10 ug/mL (Alarcón et al. 2016; Peña et al. 2012, Meccia et al. 2007). El efecto frente al crecimiento en esta bacteria puede darse a los compuestos presentes en los aceites esenciales, tales como, α- Pineno, que tiene actividad antibacteriana y se encuentra como compuesto mayoritario en estas especies. Se ha determinado que estos compuestos ejercen efectos tóxicos sobre la membrana e inhiben el transporte de iones y respiración (Magiatis 1999).
Actividad antifúngica. La evaluación frente a Fusarium sp. permitió evidenciar que entre los aceites esenciales y el control positivo (Benomil 1mg/L), no hubo diferencias estadísticas significativas (p>0.05) a las 72 h, pero si a las 96 h donde se observa que el el aceite esencial de E. barclayana presenta una inhibición similar a la del testigo Benomil (1g/L) (Tabla 3). En el caso de Chaetomium sp. a las 72 h, el que mejor respuesta tuvo fue el control comercial, sin embargo, a las 96 h de observación no se encontraron diferencias estadísticas significativas en comparación con el control positivo (Benomil 1mg/L).
Tabla 3 Porcentajes de inhibición del crecimiento micelial de los tratamientos a las 72 y 96 horas frente a Fusarium sp. D.E. = Desviación Estándar
| Tratamiento | Fusarium sp. | Chaetomium sp. | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 72 horas (p=0.0578) | 96 horas (p=0.0423) | 72 horas (p=0.0417) | 96 horas (p=0.0921) | ||||||
| Media | D.E. | Media | D.E. | Media | D.E. | Media | D.E. | ||
| T1-Benomil | 93.43 | 4.91 | 96.40 | 5.89 | 97.10 | 5.02 | 100 | 0.00 | |
| E. argentea | 63.20 | 25.33 | 70.17 | 31.69 | 61.57 | 4.52 | 73.60 | 73.60 | |
| E. barclayana | 88.67 | 8.66 | 91.83 | 3.87 | 73.93 | 15.71 | 77.30 | 77.30 | |
| E. corymbosa | 81.13 | 4.33 | 81.37 | 5.14 | 54.37 | 13.23 | 64.40 | 64.40 | |
| E. killipii | 80.13 | 1.67 | 82.87 | 3.87 | 71.73 | 4.35 | 79.10 | . | |
El seguimiento del ensayo se realizó hasta los 7 días, donde el Benomil mostró un porcentaje de inhibición promedio del 79.88%, mientras que, en los ensayos con los aceites esenciales los hongos colonizaron toda la caja. Lo anterior puede deberse a la naturaleza volátil de los compuestos presentes en los aceites esenciales (Simões & Spitzer 2000), situación que se presentó en el interior de las cajas de Petri (Rozwalka et al. 2008). Dado los resultados de actividad frente a los hongos, los aceites esenciales de E. argentea y E. corymbosa se probaron con otras cepas fúngicas: Botrytis sp., Penicillium sp. y Alternaria sp.
Botrytis sp.: el análisis estadístico de los datos mostró diferencias significativas a los dos días (p>0.05). El aceite esencial de E. argentea presentó un porcentaje de inhibición similar al del control químico después del sexto día de observación, superando el PICM mayor del 50% durante el tiempo de evaluación (Tabla 4), en el caso de E. corymbosa el PICM fue mayor al 30%. Palacios, et al. (2020), evaluaron el efecto antagónico del aceite esencial de Tagetes lemmonii, especie de la familia Asteraceae, frente a B. cinerea inhibiendo en 36%, en el mismo estudio se observó una inhibición micelial de 65.8%; resultado similar al obtenido con el aceite esencial de E. argentea a los 6 días de observación. Por su parte, el control empleado (Benomil) mostró efecto sobre B. cinerea con efectos inhibitorios hasta del 80% (Taborda et al., 2015), por lo cual, es un buen control empleado para comparar la inhibición, en este estudio el resultado del aceite esencial de E. argentea fue similar al obtenido con el control fúngico de referencia.
Tabla 4 Resultados del % de inhibición de los tratamientos frente a Botrytis sp., Penicillium sp. y Alternaria sp. en el tiempo. Control=Benomil (1mg/mL). DE= Desviación Estándar.
| Botrytis sp. ** | |||||||
|---|---|---|---|---|---|---|---|
| Tratamientos\Día | n | Día 2 (p=0.0004) | Día 6 (p=0.0131) | Día 9 (p=0.1210) | |||
| Media | DE | Media | DE | Media | DE | ||
| E. argentea | 3 | 52.78a | 8.67 | 49.51ab | 3.36 | 54.90a | 18.91 |
| E. corymbosa | 3 | 40.28a | 8.67 | 37.86a | 9.55 | 35.29a | 5.88 |
| Control | 3 | 88.89b | 2.41 | 62.14b | 3.36 | 56.47a | 4.08 |
| Penicillium sp. * | |||||||
| Tratamientos\Día | n | Día 2 (p=0.0036) | Día 6 (p=0.0071) | Día 9 (p=0.0643) | |||
| Media | DE | Media | DE | Media | DE | ||
| E. argentea | 3 | 68.00 | 3.46 | 68.00 | 3.46 | 23.27 | 4.41 |
| E. corymbosa | 3 | 86.00 | 3.46 | 86.00 | 3.46 | 65.31 | 28.28 |
| Control | 3 | 99.10 | 0.10 | 99.10 | 0.10 | 13.47 | 2.12 |
| Alternaria sp. ** | |||||||
| Tratamientos\Día | n | Día 2 (p=0.5120) | Día 6 (p=0.0104) | Día 9 (p=0.0793) | |||
| Media | DE | Media | DE | Media | DE | ||
| E. argentea | 3 | 80.95a | 8.25 | 80.95a | 8.25 | 31.11a | 13.88 |
| E. corymbosa | 3 | 80.95a | 10.91 | 80.95a | 10.91 | 48.89a | 10.18 |
| Control | 3 | 73.81a | 4.12 | 73.81a | 4.12 | 43.70a | 3.39 |
Penicillium sp.: el análisis estadístico mostró diferencias significativas en los días 2 y 6. Los ensayos con los aceites esenciales causaron un menor crecimiento fúngico con respecto al control negativo, siendo más efectivo el aceite esencial de E. corymbosa el cual presentó PICM entre el 65 a 86% para el caso de E.argentea la inhibición estuvo entre el 23 al 68% (Tabla 4). Algunos aceites esenciales, han mostrado tener efecto inhibitorio frente a Penicillium sp., como fue el caso del aceite esencial de mandarina frente a cepas de Penicillium digitatum y P. italicum causando un efecto inhibitorio en el crecimiento, esporulación y germinación del hongo (Velásquez et al. 2014). Los resultados obtenidos en esta investigación muestran que el uso de los aceites esenciales reduce el crecimiento de este microorganismo en comparación con el control negativo.
Alternaria sp.: no se observaron diferencias significativas en ninguno de los días de evaluación. Los aceites esenciales evaluados inhibieron el crecimiento del microorganismo, con porcentajes de inhibición del 31 al 80% (Tabla 4). La acción fúngica de algunos aceites esenciales frente a Alternaria sp. ha sido estudiada en diversas investigaciones mostrando efecto inhibitorio hasta del 100% (Duarte, et al., 2013).
En resumen, los aceites esenciales de las especies de Espeletias y Espeletiopsis corymbosa están constituidos principalmente por monoterpenos (48 - 90%), monoterpenoides (5 - 17%) y en menor proporción por sesquiterpenos (4 - 13%) y sesquiterpenoides (4 - 13%). Los componentes mayoritarios identificados en los aceites esenciales fueron α-Pineno, β-Pineno, Sabineno, p-Cimeno, Limoneno, α-felandreno y β-Felandreno. El análisis de clúster de la composición de los aceites esenciales mostró una mayor relación entre los aceites esenciales de las especies E. killipii y E. summapacis, y de éstos con E. corymbosa, mientras que, E. argentea presentó mayores diferencias con las demás muestras. De los aceites esenciales evaluados el de E. argentea mostró un amplio rango de inhibición frente a las cepas fúngicas de Fusarium sp., Botrytis sp., Penicillium sp. y Alternaria sp. Esta actividad puede relacionarse con la diferencia encontrada en la caracterización del aceite esencial. Este análisis es una aproximación desde los metabolitos secundarios presentes en los aceites esenciales que podrían ser correlacionadas con información genética como herramienta para la comprensión de las relaciones filogenéticas de los frailejones.
Así mismo se abre la puerta a estudios que permitan relacionar las lecciones presentes en los frailejones como son homaposis, mertesis, homosis entre otras, las cuales son causadas por cepas fúngicas y correlacionar el grado de afectación según las características de los aceites esenciales y el potencial antifúngico de especies como E.argente y E.corymbosa, las cuales en esta investigación mostraron inhibición frente a hongos patógenos y ver si existe una correlación con la presencia de estos compuestos que actúan como defensa química ante el ataque de hongos y por ende de afectaciones sanitarias.

