




![Efecto de la asistencia del líquido iónico [BMIM][BF4] en la cristalinidad y la morfología de nanoestructuras de Sb2S3 sintetizadas por ultrasonido](/img/es/next.gif)





Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Sociedad Química del Perú
versión impresa ISSN 1810-634X
Rev. Soc. Quím. Perú v.76 n.3 Lima jul./dic. 2010
TRABAJOS ORIGINALES
Algunas propiedades cinéticas de una L-aminoácido oxidasa purificada del veneno de la serpiente peruana Bothrops atrox "Jergón"
Some kinetic properties of a L-amino acid oxidase isolated from Bothrops atrox peruvian snake venom "Jergón"
Nora Ruiz1 ; Christian Solís1 ; Gustavo A. Sandoval1 ; Fanny Lazo1 ; Edith Rodríguez1 y Armando Yarlequé1*
1 Laboratorio de Biología Molecular. Facultad de Ciencias Biológicas. Universidad Nacional Mayor de San Marcos. Lima – Perú
RESUMEN
Se han determinado las principales propiedades cinéticas de una L-aminoácido oxidasa (L- AO) purificada del veneno de B. atrox, usando diferentes aminoácidos como sustratos, tales como: L-leucina, L-fenilalanina, L-metionina y L-arginina. Previamente se establecieron las curvas de pH óptimo para L-AO en función de cada aminoácido y luego se calcularon los valores de K
m, Vmax, Kcat y Kcat/Km a pH 7,5 y 8,5. Asimismo, se determinaron los valores de Ki al emplearse inhibidores específicos tales como: ácido antranílico, ácido benzoico, ácido sulfanílico y ácido salicílico. Entre los sustratos ensayados a pH 8,5, la L-AO presentó mayor actividad específica sobre L-fenilalanina, y luego sobre L-leucina, L-metionina y L-arginina, siendo los valores de pH óptimo de 8,1 con L-metionina; 8,9 con L-arginina, y 8,2 con L- fenilalanina. De acuerdo a la eficiencia catalítica, la L-leucina fue el mejor sustrato a 7,5 (Kcat/km = 40,90 x 104 s-1 M-1) y pH 8,5 (Km y Vmax (Kcat/km = 71,21 x 104 s-1 M-1). Asimismo, en los estudios de inhibición enzimática se encontró que el ácido antranílico tuvo el menor valor de Ki (0,008 mM), ajustándose al modelo de inhibición no competitiva, en tanto que el ácido benzoico fue considerado un inhibidor competitivo y los ácidos salicílico y sulfanílico se comportaron como inhibidores de tipo mixto.Palabras clave: L-aminoácido oxidasa, Bothrops atrox, parámetros cinéticos, inhibición enzimática, K
i.
SUMMARY
We have determined main kinetic properties of a L-amino acid oxidase (L-AO) purified from Bothrops atrox venom, on different aminoacids as L-leucine, L-phenylalanine, L-methionine and L-arginine. Previously optima pH curves were establish for each aminoacid and then
Km, Vmax, Kcat y Kcat/Km values at pH 7,5 and 8,5 were calculated. Furthermore Ki values using anthranilic, benzoic, sulphanylic and salycilic acids were obtained. Between substrates assayed at pH 8,5, L-AO showed higher specific activity on L-phenilalanine, followed by L- leucine, L-methionine and L-arginine, being optimum values of 8,1 for L-metionine, 8,9 for L-arginine, and 8,2 for L-fenilalanine. According to catalytic efficiency, L-leucine was the best substrate at pH 7,5 (Kcat/km = 40,90 x 104 s-1 M-1) and 8,5 (Kcat/km = 71,21 x 104 s-1 M-1). On the other hand, enzymatic inhibition assays showed that anthranilic acid had the lowest Ki value (0,008 mM), fitting it to a non-competitive model. Benzoic acid was considered a competitive inhibitor, while salicylic and sulphanylic acid showed mixed-type inhibition.Key words: L-amino acid oxidase, Bothrops atrox
, kinetics parameters, enzyme inhibition, Ki.
INTRODUCCIÓN
El veneno de las serpientes es considerado una mezcla de toxinas, enzimas y péptidos, los cuales inducen diversos efectos en sus víctimas. Su función primaria es la captura de sus presas; sin embargo, puede ser usado como defensa, causando daños severos a sus agresores, entre ellos los seres humanos
1 . Debido a su composición, los venenos de serpiente son objeto de numerosos estudios orientados a: 1) aislar una o más sustancias activas del veneno, que en bajas concentraciones puedan presentar efectos farmacológicos de gran interés para el desarrollo de medicamentos; 2) tener una mejor comprensión de la función de las sustancias aisladas del veneno de serpientes en el proceso del envenenamiento; y 3) verificar si existe algún principio activo, sintético o natural que pueda inhibir la acción del veneno y de algunas proteínas aisladas del mismo. En este aspecto, el veneno de las serpientes de la familia Viperidae presenta una composición más compleja que la de otras familias, pues contienen un gran número de proteínas bioquímica y farmacológicamente activas2.En el Perú existe una fauna ofídica muy variada habiéndose reportado hasta la fecha 47 especies de serpientes venenosas, de las cuales 24 pertenecen al género Bothrops. Entre ellas se destaca la especie Bothrops atrox, perteneciente a la familia Viperidae
3 , por ser la causante del 90% de mordeduras, lo cual constituye un problema de salud pública muy importante4 . Dentro de los principales componentes del veneno de B. atrox, estudiado en el Laboratorio de Biología Molecular de la Facultad de Ciencias Biológicas, encontramos a la L-aminoácido oxidasa o L-AO (EC 1.4.3.2), la cual es una flavoproteína responsable del color amarillo de los venenos y es específica para la desaminación oxidativa de un L-aminoácido hasta un α-cetoácido con la consiguiente producción de amoniaco y peróxido de hidrógeno5 . Además, se han aislado y caracterizado otras L-aminoácido oxidasas como las de Lachesis muta6 y Bothrops brazili7.En cuanto al estudio de L-AO de B. atrox, Lazo et al.
5 aislaron y determinaron varias características bioquímicas de esta enzima, como el peso molecular y la determinación de azúcares asociados. Así, también se determinó su acción biológica, encontrándose que presenta efecto antimicrobiano in vitro sobre microorganismos patógenos, efecto in vitro contra promastigotes de Leishmania braziliensis braziliensis, y contra epimastigotes de Trypanosoma cruzi, además de un marcado efecto edemático8 . Dado el potencial biotecnológico de la L-aminoácido oxidasa del veneno de la serpiente de mayor interés médico en Perú, Bothrops atrox, es necesario evaluar sus principales propiedades cinéticas. Por lo tanto, el objetivo del presente trabajo fue estudiar experimentalmente la cinética enzimática de esta L-AO para evaluar el efecto que tiene sobre ella, la naturaleza química de los sustratos, como también ensayar posibles inhibidores, evaluando el grado y la naturaleza de la interacción, para seleccionar así los mejores inhibidores candidato lo que permitirá establecer posibles mecanismos de regulación de esta enzima.PARTE EXPERIMENTAL
Enzima purificada
Se empleó la L-aminoácido oxidasa del veneno de B. atrox, la cual fue purificada siguiendo la metodología reportada previamente por Lazo et al
5 , utilizando para esto una cromatografía de filtración molecular en Sephadex G-75, seguido de una cromatografía de intercambio iónico en CM Sephadex C-50. La enzima purificada fue almacenada a -20 ˚C hasta la realización de los ensayos cinéticos.Sustratos e inhibidores en estudios
Como sustratos se emplearon los siguientes L-aminoácidos: L-leucina, L-metionina, L- arginina y L-fenilalanina (Sigma Chemical Co.) (figura 1). Asimismo, para los estudios de inhibición enzimática se utilizaron los siguientes compuestos: ácido salicílico, ácido sulfanílico, ácido benzoico y ácido antranílico. (Sigma Chemical Co) (figura 2).
Contenido proteico
La cantidad de proteína fue calculada midiendo la absorbancia de luz UV a 280 nm
9 en un espectrofotómetro Shimadzu UV 120-02. Además, se empleó el método de Lowry10 modificado en nuestro laboratorio11 utilizando un fotocolorímetro Spectronic Bausch & Lomb, empleando albúmina sérica bovina como proteína estándar.Actividad de L-aminoácido oxidasa
Se determinó por el método descrito en el Worthington Enzyme Manual
1 2 , utilizando Buffer Tris-HCl 0,2 M pH 7,5 conteniendo O-dianisidina 0,0065%, peroxidasa de rabanito al 0,001% y L-leucina 1%. La mezcla fue preincubada durante 5 minutos a 37 ºC y luego se agregaron 20 μl de la solución de enzima, determinándose el incremento de absorbancia a 436 nm. La actividad enzimática fue expresada en μmoles de L-leucina oxidados por minuto. En tanto que la actividad específica se expresó en U/mg de proteína. Una unidad de actividad corresponde a la oxidación de 1 μmol de L-leucina por minuto. Del mismo modo, se evaluó la actividad de L- AO sobre L-fenilalanina, L-arginina y L-metionina.Determinación del pH óptimo
Se determinaron los pH óptimos de actividad de L-AO sobre L-fenilalanina, L-metionina y L-arginina siguiendo el método descrito en el Worthington Enzyme Manual
1 2 . Para esto se utilizaron en la mezcla de reacción buffer acetato de amonio 0,2 M (pH 5,0, 5,5, 6,0 y 6,5), buffer Tris-HCl 0,2 M (pH 7,0, 7,5, 8,0, 8,5, 9,0, 9,5) y buffer glicina-NaOH 0,2 M (pH 10,0 y 10,5). Los valores de pH óptimo se obtuvieron mediante el Modelo de Gauss utilizando el programa Microcal Origin versión 6,0.Determinación de los parámetros cinéticos
Se determinó el comportamiento cinético de la L-AO purificada en buffer Tris-HCl 0,2 M sobre L-leucina, L-metionina, L-fenilalanina y L-arginina a pH 7,5 y pH 8,5. La concentración de enzima utilizada fue de 6,85 x 10
-9 M, la cual se calculó a partir del peso molecular de la enzima (127,87 kDa) estimado previamente5 y expresado en g/mol. El incremento en la absorbancia a 436 nm se calculó de la porción inicial linear de la velocidad. Adicionalmente, se estimaron los parámetros cinéticos Km y Vmax según la ecuación de Michaelis-Menten (Vo = Vmax[S]/ km + [S]), mediante la representación de Michaelis Menten y Lineweaver-Burk, utilizando el software Sigmaplot versión 10,0. Se calcularon además los valores de Kcat y eficiencia catalítica.Ensayos con inhibidores sintéticos de la actividad enzimática
Se determinó la acción inhibitoria del ácido salicílico, ácido benzoico, ácido sulfanílico y ácido antranílico (0,005 - 16 mM sobre la actividad de L-AO utilizando L-leucina (0,2 - 1,5 mM) como sustrato a pH 8,5.mM. obteniéndose las curvas respectivas con las diversas concentraciones de inhibidor. En cada caso se determinó el tipo de inhibición y la constante de inhibición K
i, de acuerdo a la representación de Lineweaver-Burk, utilizando el software Sigmaplot versión 10,0.RESULTADOS Y DISCUSIÓN
Actividad de L-AO y determinación del pH óptimo
De los ensayos de actividad de L-AO, se encontró que la mayor actividad específica sobre los aminoácidos L-metionina, L-leucina, L-arginina y L-fenilalanina, a una concentración de 7,5 mM y pH 8,5. Asimismo, la enzima demostró tener acción en el rango de pH de 6,9 a 9,6, obteniéndose valores de pH óptimo de 8,9 para L-arginina, 8,1 para L-metionina y 8,2 para L- fenilalanina (tabla 1).
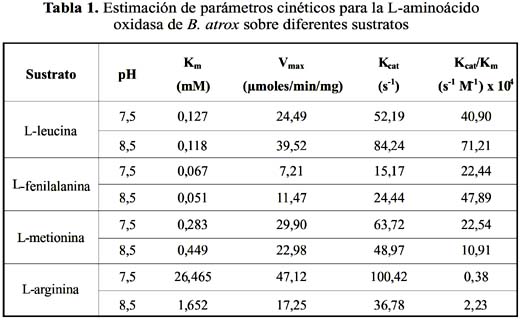
Parámetros cinéticos para los principales sustratos
Los valores de V
max (µmoles/min/mg) y Km (mM) obtenidos a pH 7,5 y 8,5 se muestran en la tabla 1. Para todos los casos se observó que la enzima se ajustó al modelo de cinética Michaeliana a concentraciones bajas de sustrato y también se observó inhibición por sustrato a altas concentraciones. Los menores valores de Km se obtuvieron a pH 8,5 para los L-aminoácidos: L-leucina, L-fenilalanina, y L-arginina, mientras que para L-metionina el menor valor de Km se obtuvo a pH 7,5. Asimismo, se muestran los valores de Kcat y eficiencia catalítica (Kcat/Km) para dichos sustratos.Modelo de inhibición y valores de Ki
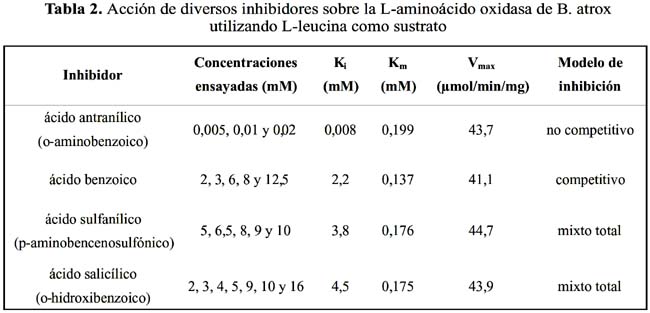
La actividad de L-AO fue inhibida de modo no competitivo por ácido antranílico con constante de inhibición (K
i) de 0,0082 mM. La inhibición de L-AO fue competitiva cuando se empleó ácido benzoico, con un valor de Ki de 2,2 mM. Al emplearse ácido sulfanílico y ácido salicílico, el modelo de inhibición fue mixto con valores de Ki de 3,8 y 4,5 mM, respectivamente (tabla 2).
Actividad de L-AO sobre diversos sustratos y pH óptimo
Se ha reportado que este tipo de enzimas presenta una especificidad muy marcada para cierto tipo de sustratos L-aminoácidos, la cual varía dependiendo de la especie de serpiente en estudio. De los resultados obtenidos en el presente estudio, la L-AO de B. atrox mostró mayor preferencia por aminoácidos apolares hidrofóbicos, como la L-leucina, L-fenilalanina y L- metionina, seguido por aminoácidos polares básicos, como la L-arginina. Esto sugiere que la cadena lateral del aminoácido debe ser apolar para que el sustrato sea más específico, debiendo existir en el centro activo de la enzima una región apolar que interaccione con dicha cadena lateral. La preferencia por este tipo de aminoácidos hidrofóbicos ha sido reportada para la L-AO de Bothrops pirajai
13. Asimismo, se encuentran resultados similares reportados para las L-AO de Lachesis muta6 y Bothrops brazili7.Con respecto a los ensayos para establecer el pH óptimo de L-AO de B. atrox, la máxima actividad se registra a pH 8,3 usando L-leucina como sustrato, siendo este valor similar al reportado previamente para esta misma enzima
5 . Además, se encontró que la enzima mantiene un 80% de esta actividad máxima en el rango de pH 7,5 a 8,7. Estos valores de pH óptimo son semejantes a los reportados para las L-AO de Lachesis muta6 (pH óptimo: 7,5, sobre L- leucina), Bothrops brazili7 (pH óptimo: 9,0, sobre L-lisina; 8,5 sobre L-leucina, L-metionina, L-fenilalanina y L-arginina; 8,0 sobre L-isoleucina y 7,5 sobre L-triptófano) y Bothrops pirajai13 (pH 6,0 y 7,4 sobre L-fenilalanina y L-leucina, respectivamente). En todos los casos reportados se observa que el pH óptimo varía dependiendo del aminoácido utilizado como sustrato.Análisis cinético
La L-AO de B. atrox exhibió una cinética enzimática que se ajusta al modelo de Michaelis- Mente, donde el valor de K
m corresponde a la concentración de sustrato a la cual se alcanza la mitad de la velocidad máxima, Vmax, y es una medida de la afinidad de la enzima por el sustrato. En la tabla 2 se muestran los valores de Km para la L-aminoácido oxidasa empleando L- leucina, L-metionina y L-arginina como sustratos. A pH 7,5 y 8,5, se observa que la cadena lateral del aminoácido debe ser apolar para que el sustrato sea más específico, obteniéndose valores de Km menores. Esto nos indica que en el centro activo de la enzima existiría una región apolar que interacciona con la cadena lateral de los aminoácidos apolares. Los menores valores de Km se obtuvieron a pH 8,5, y son resultados muy similares a los obtenidos para Calloselasma rhodostoma14, N. naja kaouthia15,16 y Ophiophagus hannah17, donde se encuentra la misma especificidad, es decir: L-fenilalanina > L-leucina > L-metionina > L- arginina. Esto demuestra que las L-AO de venenos de serpientes exhiben un comportamiento cinético muy similar.La presencia de una región apolar en el centro activo podría favorecer una mayor actividad de la enzima respecto a otras del mismo tipo, ya que polariza no sólo el grupo catalítico sino también los posibles enlaces con el sustrato, exhibiendo una mayor reactividad. En cuanto a la eficiencia catalítica encontramos que el mayor valor lo exhibe L-leucina tanto a pH 7.5 como 8.5, lo cual está en relación con su grado de hidrofobicidad y sobretodo por su abundancia en el medio celular
1 9 .Modelos de inhibición
Debido a la preferencia de L-AO de B. atrox sobre aminoácidos hidrofóbicos y aromáticos, se evaluó la actividad inhibitoria de un grupo de moléculas sintéticas que presenten un anillo aromático en su estructura, ya que esta característica ofrecería un mayor ajuste del sustrato al sitio activo. De los parámetros cinéticos que muestra el efecto de inhibición enzimática se consideró el valor de K
i (constante de inhibición), que corresponde a la concentración de inhibidor que reduce a un tercio el valor de Vmax, cuando [S] = Km. El valor de Ki es comparable al de Km ya que revela el grado de afinidad de la enzima por el inhibidor. De acuerdo a los resultados de la tabla 2, el ácido antranílico, ácido acetilsalicílico, ácido benzoico y ácido sulfanílico, inhibieron la actividad de L-AO, si bien en diverso grado y presentando diferentes modelos de inhibición.En el caso de la inhibición por el ácido benzoico, ésta se ajustó al modelo de inhibición competitiva, la cual se explicaría por la presencia del anillo bencénico. Por otro lado, el ácido antranílico u o-aminobenzoico, resultó ser el inhibidor más potente, donde la presencia del sustituyente amino en el anillo bencénico incrementa la polaridad positiva y por ende la afinidad de la enzima por esta molécula. Estas moléculas presentan una semejanza estructural con los sustratos, por un lado poseen un grupo carboxilo cargado y por otro un grupo hidrofóbico, el anillo bencénico: por lo que se estarían reaccionando en la misma región de la enzima que en el caso de los sustratos. Estos resultados sugieren que las porciones electrostáticas e hidrofóbicas presentes en la estructura del inhibidor estarían involucradas en su trayectoria hacia el sitio activo, siendo ambas características complementarias para lograr una inhibición de esta enzima. Sin embargo, el modelo de inhibición para el ácido antranílico es no competitivo, lo que sugeriría que la interacción de éste con los residuos positivos no se da a nivel de residuos básicos en el sitio activo, sino más bien a lo larga de la trayectoria hacia el sitio activo.
Diversos estudios de inhibición enzimática realizados con las L-aminoácido oxidasas han mostrado que este tipo de enzimas responde al modelo de inhibición competitiva. En 1948, Zeller señaló que el ácido benzoico, ácido salicílico, ácido mandélico, las sulfonamidas, los ácidos sulfónicos aromáticos y los ácidos alifáticos
α-aminosulfónicos, son inhibidores competitivos de la L-aminoácido oxidasa de V. aspis. Asimismo, De Kok et al. 1 8, utilizando diferentes inhibidores competitivos realizó un análisis del mecanismo de unión de los inhibidores al sitio activo de la enzima de C. adamanteus, observándose que los ácidos benzoicos metasustituidos fueron los inhibidores más fuertes.Con respecto al ácido sulfanílico y al ácido salicílico, los valores de K
i son mucho mayores, lo que demuestra que son menos afines a la enzima. Este se debe probablemente a que exhiben una mayor acidez (valores de pKa menores), lo cual constituye un factor que podría incidir en la fuerza de unión del sustrato a la enzima. Además, estos ácidos exhibieron un patrón de inhibición del tipo mixto. Estos resultados se complementa con los reportados por Page y Vanetten20 para la L-AO de C. adamanteus, quienes indican que el mecanismo de catálisis general implicado en la reacción enzimática utilizando L-leucina como sustrato es de naturaleza básica.Diversas estructuras de inhibidores sugieren que los compuestos aromáticos que tienen un grupo carboxilo o hidroxilo son capaces de inhibir a las enzimas L-AO
1 8. La similar afinidad que presentan los ácidos grasos sustituidos y no sustituidos por la L-AO, sugiere que el efecto de adicionar el componente aromático resulta comparable a incrementar el tamaño de la cadena alifática. Aún cuando no se ha establecido, es razonable suponer que la interacción entre L-AO y los aminoácidos involucran tanto a los grupos amino como a los carbonilo. Del mismo modo, se esperaría que los grupos ácidos del inhibidor reaccionen en la misma porción de L-AO que las porciones carboxilo de los aminoácidos. Por estas razones se cree que los inhibidores que contengan tanto el grupo básico como ácido pueden ser más efectivos que los que sólo cuenten con uno de éstos. Otro factor estructural es el cofactor FAD, que tiene una importante interacción con los átomos de la proteína, siendo considerado inclusive como un dominio de ésta, por lo que se piensa que el incremento en el ambiente apolar alrededor del FAD puede perturbar su espectro de absorción, afectando así también la actividad enzimática. Además, el grupo FAD de la enzima es importante para establecer interacciones con los sustratos los residuos de aminoácidos básicos tales como la histidina 461, arginina 88 y lisina 324. Esto explica la afinidad a moléculas como los ácidos benzoicos que presentan como sustituyentes característicos un grupo carboxilo, grupo también presente en los sustratos L- aminoácidos de esta enzima.A partir de este análisis, se puede decir que la estructura farmacofórica de un inhibidor de la L- aminoácido oxidasa apropiado para especies del genero Bothrops debe poseer al menos un grupo aromático con un sustituyente con carga negativa y otro sustituyente con polaridad positiva, con el fin de que tenga una mayor fuerza de unión al centro activo de la enzima, inhibiendo de esta manera su actividad enzimática pudiendo reducir así los efectos tóxicos del veneno.
CONCLUSIONES
La presente investigación ha permitido establecer que la L-aminoácido oxidasa del veneno de la serpiente peruana Bothrops atrox es capaz de hidrolizar diversos L-aminoácidos; siendo más activo sobre los L-aminoácidos hidrofóbicos y aromáticos. Esta afinidad por determinados sustratos servirá de base para el diseño de moléculas sintéticas con residuos aromáticos y sustituyentes con carga negativa y positiva que puedan inhibir esta enzima, reduciendo así sus efectos tóxicos.
AGRADECIMIENTOS
El presente trabajo fue parte de la Tesis de Biólogo con mención en Biología Celular y Genética de la Blga. Nora Ruiz. Ademas, los autores del presente trabajo agradecen al Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica (CONCYTEC) por el apoyo financiero brindado para llevar a cabo esta investigación (Proyecto PROCYT Nº 161-2006- CONCYTEC-OAJ).
REFERENCIAS
1. Tu, A.T. Venoms: chemistry and molecular biology. Ed. John Wiley & Sons, New York, 1977. p.560.
2. Suhr, S.M. y Kim, D.S. Identification of the snake venom substance that induces apoptosis.
Biochem Biophys Res Commun 1996; 224 (1): 134-139.3. Carrillo, N. e Icochea, J. Lista taxonómica preliminar de los reptiles vivientes del Perú. Publicaciones del Museo de Historia Natural Javier Prado. UNMSM. 1995; Serie A (49): 27.
4. Loja, D., Avilés, R., Necochea, Y., Vilca, M. y Castro, J. Ofidismo por Bothrops atrox: estudio clínico-epidemiológico. Diagnóstico 2000; 39(5):261-5.
5. Lazo, F., Málaga, O., Yarlequé, A. et al. Algunas propiedades bioquímicas de una L- aminoácido oxidasa aislada del veneno de la serpiente Bothrops atrox. Rev. Soc. Quím. Perú, jul./set. 2007, vol.73, no.3, p.131-141.
6. Cisneros, Y.; Lazo, F., Gutiérrez, S., Yarlequé, A. Características bioquímicas de una proteína antibacteriana aislada del veneno de Lachesis muta "Shushupe". Rev. Soc. Quím. Perú, oct./dic. 2006, vol.72, no.4, p.187-196.
7. Solis, C., Escobar, E., Yarlequé, A. y Gutiérrez, S. Purificación y caracterización de la L- aminoácido oxidasa del veneno de la serpiente Bothrops brazili "Jergón shushupe" Rev Peru Biol, 1999; 6(1): 75-84.
8. Lazo, F., Málaga, O., Yarlequé, A. et al. Actividad antimicrobiana de una flavoproteína aislada del veneno de la serpiente peruana Bothrops atrox ("jergón").
Rev. Soc. Quím. Perú, oct./dic. 2007, vol.73, no.4, p.197-207.9. Warburg, O. and Christian, W. Isolierung and cristallisation del Gärungs ferments enolase. Biochemische Zertschrift. 1941; 31: 384-421.
10. Lowry, O.H., Rosebrough, N.J., Farr, A.L. y Randall, R. Protein measurement with the folin phenol reagent.
J Biol Chem 1951; 193: 265-275.11. Loayza, S., Morante, Y., Campos, S., y Yarlequé, A. Enzimas proteolíticas en el veneno de las serpientes peruanas Lachesis muta y Bothrops atrox.
Bol Soc Quim del Perú 1985; LII (3): 151-163.12. Worthington Biochemical Corporation. L-Amino Acid Oxidase. New Jersey: The Worthington Enzymes Manual. Enzymes Related Biochemicals; 1993 p. 34-35.
13. Izidoro, L.F., Ribeiro, M.C., Souza, G.R., Sant`ana, C.D., Hamaguchi, A., et al. Biochemical and functional characterization of an L-amino acid oxidase isolated from Bothrops pirajai snake venom. Bioorg. Med. Chem., 2006. v.14, p.7034-43.
14. Ponnudurai, G., Chung, M.C. y Tan, N.H. Purification and properties of the L-amino acid oxidase from Malayan pit viper (Calloselasma rhodostoma) venom. Arch Biochem Biophys 1994; 313(2): 373-378.
15. Sakurai, Y., Takatsuka, H., Yoshioka, A., Matsui, T., Suzuki, M. Titán, K. y Fujimura, Y. Inhibition of human platelet aggregation by L- amino acid oxidase purified from Naja naja kaouthia venom. Toxicon 2001; 39(12): 1827-1833.
16. Tan, N.H. y Swaminathan, S. Purification and properties of the L-amino acid oxidase from monocellate cobra (Naja naja kaouthia) venom. Internat J Biochem 1992; 24: 967-973.
17. Li, Z.Y., Yu, T.F. y Lian, E. Purification and characterization of L-amino acid oxidase from king cobra (Ophiophaqus hannah) Venom and its effects on human platelet aggregation. Toxicon 1994; 32(11): 1349 – 1358.
18. De Kok, A. y Veeger, C. Studies on L- amino acid oxidase I. Effects and the competitive inhibitors. Biochim Biophys Acta 1968; 33:345.
19. Du, X.Y. y Clemetson, K.J. Snake venom L- amino acid oxidases. Toxicon 2002; 40(6):659-665.
20. Page, D. y Van Etten, R. L- amino acid oxidase.
II. Deuterium isotope effects and the actino mechanism for the reduction of L- amino acid oxidase by L- leucine. Biochim Biophys Acta 1971; 227: 16-31.
