











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
Pendimethalin [N-(1-etilpropinil)-2,6-dinitro-3,4-xilidina] es un herbicida pre-emergente que pertenece a la familia de las dinitroanilinas (Vera et al., 2023). Este herbicida actúa sobre la división celular y su acción se produce porque impide la formación de microtúbulos durante la división mitótica de las células (Appleby & Valverde, 1989). Por lo general, es usado antes de la siembra en cultivos de ciclo corto (Onwuchekwa-Henry et al., 2023). Sin embargo, el constante uso y las altas dosis generan contaminación a los acuíferos a través de escorrentía, lixiviación, evaporación y deriva. Además, ocasionan reducción de la microbiología del suelo, necrosis y disminución de la biomasa foliar y radicular en plantas (Juraeva et al., 2023; Khaliq & Matloob, 2012; Pinilla, 2020). Se han reportado contaminación de pendimentalina y afectación a los habitantes en afluentes de aguas de Francia (840 ng/L); Brasil (0,06 y 0,38 μg/L), así mismo en Ecuador (0,170 - 0,557 μg/L) en la zona arrocera de la cuenca del rio Guayas, por tal motivo, la constante aplicación de este herbicida es una amenaza para la salud humana (Deknock et al., 2019; Le Du-Carree et al., 2021; Qiao et al., 2021).
Una alternativa para transformar los plaguicidas en compuestos más simples y reducir contaminantes en el ecosistema es mediante el uso del potencial metabólico de los microorganismos (Dubey, 2021; Hernández et al., 2017; Neamat‐Allah et al., 2021; & Trivedi). Las rizobacterias promotoras de crecimiento vegetal por sus siglas en inglés Plant Growth-Promoting Rhizobacteria (PGPR) son conocidas por incrementar el desarrollo de las plantas, reducir el estrés biótico y abiótico, incluso mostrar propiedades tolerantes/degradantes a los pesticidas (Oleńska et al., 2020; Hkudaygulov et al., 2022).
Las PGPRs producen enzimas hidrolasa, oxidorre-ductasa, deshalogenasa, oxigenasa y transferasa, que funcionan mediante reacciones catalíticas para descomponer y reducir los contaminantes ambien-tales dañinos (Jeong & Choi, 2020; Saravanan et al., 2021). Además, estos microorganismos tienen varios mecanismos para tolerar los herbicidas, como la bioaumentación. Se considera una forma eficiente de remediar sitios contaminados con pendimentalina mediante la introducción de microorganismos degradantes específicos (Wang et al., 2018). Singh & Singh, (2020) demostraron que ciertas rizobacterias toleraban 1000 mg/L de pendimentalina en condiciones in vitro. Moyano (2015) señala que la degradación de pendimetalina por rizobacterias sucede existiendo una relación entre la temperatura y el pH. Rosado et al. (2020) reportaron que las bacterias Acinetobacter pitii, Atlantibacter hermanii y Pseudomonas plecoglossicida presentaron un crecimiento con 2.0 g/L con el pendimethalin, por lo tanto, son consideradas como tolerantes.
En la actualidad, se han realizado métodos y procesos como técnicas de enriquecimiento, detección funcional, metagenómica, transferencia horizontal de genes y estrategias de ingeniería genética, para identificar microorganismos degradadores de pesticidas en los procesos de biodegradación y biomineralización (Verma et al., 2014). Sin embargo, en el Ecuador existen pocos reportes sobre rizobacterias tolerantes al pendimentalina como posible alternativa para su uso en suelos contaminados. En este contexto, como alternativa para prevenir la contaminación ambiental de pendimentalina, se planteó como objetivo evaluar el efecto de la pendimentalina sobre el crecimiento celular de las PGPR.
METODOLOGÍA
Ubicación
La investigación se realizó en la Universidad Técnica Estatal de Quevedo, km 7½ de la Vía Quevedo, El Empalme, cantón Mocache, Los Ríos, coordenadas geográficas 1°04'51,7 latitud Sur 79°30'10,0" longitud Oeste, altitud de 66 m.s.n.m.
Selección y preparación del inóculo bacteriano
Del Banco de Germoplasma del Laboratorio de Microbiología Molecular de la UTEQ, fueron seleccionadas las rizobacterias provenientes de cultivares endémicos de musáceas con su clasifica-ción a metabolitos secundarios (Tabla 1). Las rizobacterias se incubaron en 100 mL de King B (KB) líquido selectivo [(g/L): peptona de carne 20,0; glicerol, 15,0 mL; fosfato dipotásico; 1,5; sulfato de magnesio heptahidratado, 1,5; agua destilada (pH 7,2)], suplementado con chloram-phenicol (13 μg/mL), ampicilina (40 μg/mL) por 48 h (King et al., 1954).
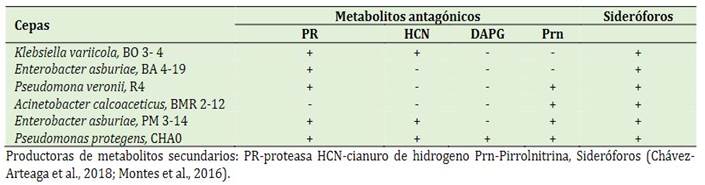
Adaptabilidad de las rizobacterias a concentra-ciones de pendimentalina en condiciones in vitro
Las cepas fueron tomadas por individual, y se ubicaron en un Erlenmeyer de 50 mL con 40 mL de medio de cultivo King B líquido (KBL) g/L: (peptona 10 g, sulfato de magnesio 1,5 g, fosfato dipotásico 1,5 g y glicerina 15 ml suplementado con cloranfenicol 40 mg) a concentraciones de 0, 50, 100 y 150 mg/L de GRAMILAQ (pendimentalina 400 g/L). Las muestras se incubaron por 180 rpm a 28 °C durante 72 horas. A las 24, 48 y 72 horas se tomaron muestras para determinar las siguientes variables: pH mediante el potenciómetro OHAUS modelo STARTER y la absorbancia a 600 nm con el espectrofotómetro "UNICO" modelo 1205.
Niveles de tolerancia de las rizobacterias a la Pendimentalina bajo diferentes condiciones abióticas in vitro
Se seleccionaron 3 rizobacterias por su tolerancia a la pendimetalina. Las cepas se incubaron en KBL suplementado de 150 mg/L de GRAMILAQ® 400 en 40 mL. Los cultivos se ajustaron a diferentes pH, considerando que 5,0 y 6,5 se agregó ácido clorhídrico (HCl) al 0,1% y 8,0 hidróxido de sodio (NaOH) al 0,1 M. Las muestras se dejaron en incubación a 180 rpm durante 72 horas. Cada 24, 48 y 72 horas se tomaron las siguientes variables: Las UFC/ml se evaluaron con la metodología descrita por (Dickson, 1993). Los valores de pH mediante el potenciómetro OHAUS modelo STARTER y la absorbancia a 600 nm con el espectrofotómetro "UNICO" modelo 1205.
Reducción de estrés por rizobacterias en suelos contaminado con Pendimentalina
Se colectó muestra de suelo sobre los primeros 0,20 m de profundidad procedente de una finca produc-tora de arroz, de la provincia de Los Ríos, Cantón “Mocache”. El suelo se solarizó con la metodología descrita por Navarro (1991). Después de 10 días, se homogenizó y se tamizó con una malla < 2 mm. El suelo se mezcló con turba y perlita en relación 3:1:1. El sustrato se llevó a Capacidad de Campo (CC), se agregó dosis de 0, 75 y 150 mg/kg de pendimentalina. Las muestras de suelo se incubaron durante 15 días, manteniendo en CC el sustrato. Las semillas del material de arroz INIAP FL - ÉLITE se esterilizaron de forma super-ficial con alcohol (70%) durante 30 s y NaOCl (1% V/V) durante 60 s, luego se enjuagaron tres veces con agua destilada estéril. Se colocaron 10 semillas en cajas de Petri estériles que conteniá 40 g de sustrato. Las rizobacterias del anterior experi-mento crecieron en KBL a 170 rpm durante 24 horas. De forma individual, se homogenizaron en relación v/v al 30% y se obtuvo una concentración celular de (8,6E+7 UFC/ml) aplicando 2 mL de la solución bacteriana alrededor del suelo. Las cajas de Petri se sellaron con papel parafilm y se incu-baron a temperatura ambiente (25 °C) bajo un fotoperiodo de 12 h luz-12 h oscuridad durante 96 h. Se tomaron las siguientes variables: semillas germinadas, crecimiento temprano (longitud de la raíz y del hipocótilo). El índice de vigor (IV) y el porcentaje de germinación se calculó con lo descrito por Aparicio et al. (2018) y Zucconi et al. (1981).
Diseño experimental y análisis de datos
Para los ensayos in vitro se aplicó un diseño completamente al azar (DCA) con factorial, para cepa y dosis de herbicida. Con tres réplicas y dos unidades experimentales. A nivel invernadero, se empleó un DCA con factorial, para cepas y dosis de pendimentalina. Cada tratamiento contenía tres repeticiones, con tres unidades experimentales, se evaluaron nueve plantas por tratamiento. Para el análisis se utilizó el software estadístico Infostat 2020 y la versión 19.1.0 del software Minitab (Di Rienzo et al., 2020; Minitab, 2021).
RESULTADOS Y DISCUSIÓN
Densidad óptica de las rizobacterias a diferen-tes dosis de pendimentalina
Se presentó significancia estadística en los factores en estudio y las interacciones (p < 0,0001) durante las 24, 48 y 72 h. A las 24 h, las cepas CHA0 y BA4-19 con 150 mg/L mostraron mayor densidad óptica (2,28 y 2,27). A las 48 h se presentó un ligero incremento con 2,33 y 2,32. Esto evidencia que las rizobacterias en estudio tienen una rápida adapta-bilidad al herbicida. Sin embargo, a las 72 h se evidenciaron que las cepas bacterianas (CHA0, BA4-19, B03-4, PM3-14 y R4) con aplicación de 150 mg/L tuvieron mayor absorbancia (2,38 y 2,51) a diferencia del sin aplicación (1,74 y 0,99) (Tabla 2). Esto indica que las rizobacterias, además de tolerar el herbicida puede seguir con sus actividades metabólicas para su desarrollo. La biodegradación por microorganismos comienza con oxidación, reducción, hidrólisis y desalquilación; así como, diversas rutas metabólicas como el metabolismo energético mitocondrial, el metabolismo de los ácidos grasos y lípidos, el metabolismo de los aminoácidos, las vías oxidativas e hidrolíticas y la metilación (Singh & Singh, 2016; Guerrero Ramírez et al., 2023). El análisis de la dinámica poblacional de Bacillus subtilis, Pseudomonas fluorescens y Escherichia coli muestran un aumento celular al consumir el herbicida como única fuente de carbono después de siete días después de la inoculación en el medio modificado con 125 mg/L pendimetalina (Avarseji et al., 2021). Esto concuerda con Kulshrestha et al., (2000) donde realizaron cuatro aplicaciones por dos años de pendimetalina en el cultivo de maíz observaron que a una superficie del suelo (0-15 cm) muestra mayor degradación del herbicida del 54%.
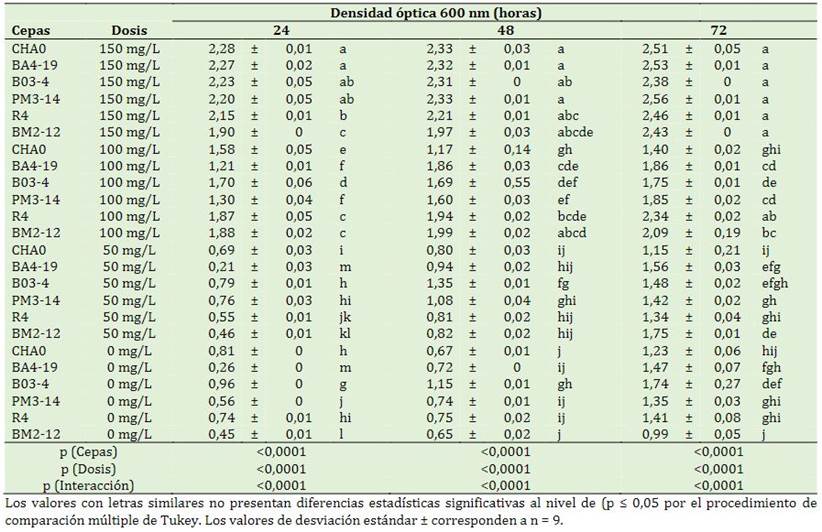
pH de las rizobacterias a diferentes dosis de pendimentalina
Se presentó significancia estadística en los factores en estudio y las interacciones (p < 0,0001) a las 24, 48 y 72 h. A las 24 horas, la cepa BM2-12 tuvo un mayor incremento del pH cuando no existió la aplicación de pendimentalina (7,55). A las 48 h, la cepa PM3-14 con 100 mg/L demostró un incremento en el pH (7,92) en comparación a los demás tratamientos. Sin embargo, a las 72 h la cepa BM2-12 sometida a 50 mg/L aumentó el pH (8,29) (Tabla 3).
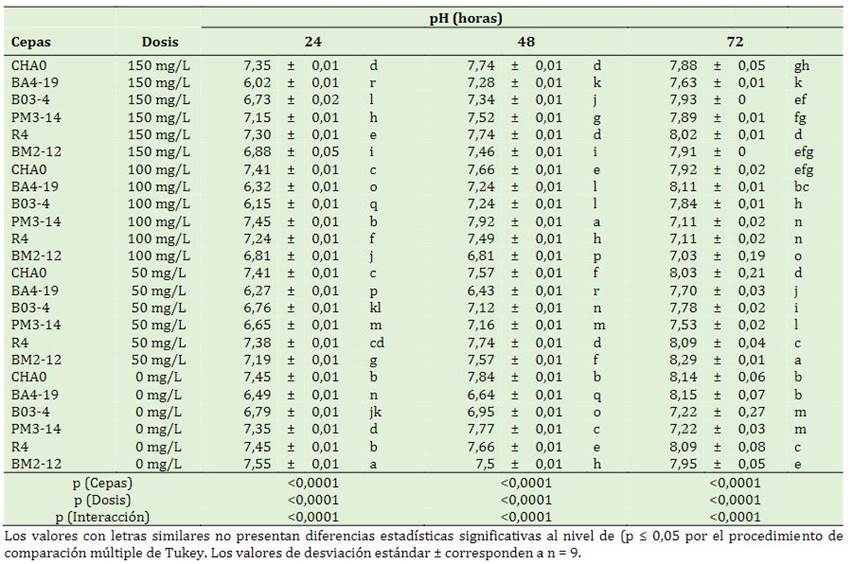
Es posible que la acción metabólica de las rizobac-terias produzca reacciones alcalino-reductoras en dosis específicas de contaminantes, a su vez, la respuesta de las cepas al disminuir el pH podría estar influenciado por los ácidos orgánicos. Estas bacterias modifican el pH por la producción de ácidos orgánicos, en cambio cuando se parte con pH ligeramente alcalino, las bacterias regulan comenzando a las 96 h hasta obtener un pH neutro esto se da por la segregación de amonio a partir de desaminación de aminoácidos (Lei et al., 2023). Clavispora lusitaniae reduce el pH del 7,51 inicial a 5,5 durante la degradación de la pendimetalina a las 96 h post-incubación (Han et al., 2019). El aumento del pH de la solución juega un rol clave en la biosorción de herbicidas, probablemente podría atribuirse a la acumulación en el estado de ionización de los grupos funcionales como carbo-xilato, fosfato y grupos amino de la pared celular, compiten con iones que coexisten en la solución (Baxter & Cummings, 2008; Ibrahim et al., 2023).
Efecto del pH sobre la densidad óptica de las rizobacterias
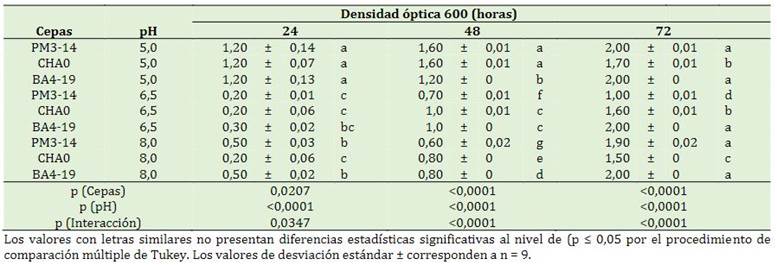
Se presentó significancia estadística en los factores en estudio y las interacciones (p < 0,0001) a las 48 y 72 h. A las 24 h el pH alcalino y neutro afectó la densidad óptica de las rizobacterias, en compa-ración al pH ácido (5,0) donde mostró un mayor incremento en las cepas PM3-14, CHA0 y BA4-19 con 1,23, 1,19 y 1,1, respectivamente. A las 48 h se presenció un desarrollo de células en el pH neutro y alcalino, sin embargo, existió mayor suspensión bacteriana en las rizobacterias PM3-14 y CHA0 con (1,6 y 1,6). Con respecto a las 72 h, se demostró una estabilidad en las células en pH acido, neutro y alcalino (Tabla 4).
La bacteria BA 4-19 posee un creciente crecimiento microbiano acelera la tasa de degradación de pendimetalina en condiciones ambientales desfa-vorable consume la fuente alternativa de carbono, fosfato y energía del herbicida. En esta etapa, las células se dividen mediante fisión binaria y dupli-can su número después de cada tiempo de genera-ción (Doolotkeldieva et al., 2018). La actividad metabólica es alta debido a la capacidad de las bacterias que utilizan el glifosato de carbono como fuente de energía para el crecimiento y la división celular (Li et al., 2021). La cinética de crecimiento celular de la bacteria GP-1 genera proceso de descontaminación del Glifosato de 1000 ppm a partir de octavo día (Malviya et al., 2015).
Efecto del pH sobre las rizobacterias
Se presentó significancia estadística en los factores en estudio y las interacciones (p < 0,0001) a las 24, 48 y 72 h. A las 24 h, el pH alcalino presentó una disminución en la cepa CHA0 con (7,7), sin embargo, en condiciones ácidas, se evidenció un incremento con las cepas PM3-14, CHA0, BA4-19 con (6,8, 7,0 y 6,7) respectivamente. A las 48 y 72 h se presenció una estabilidad en el pH desde ligeramente alcalino a casi neutro (Tabla 5).
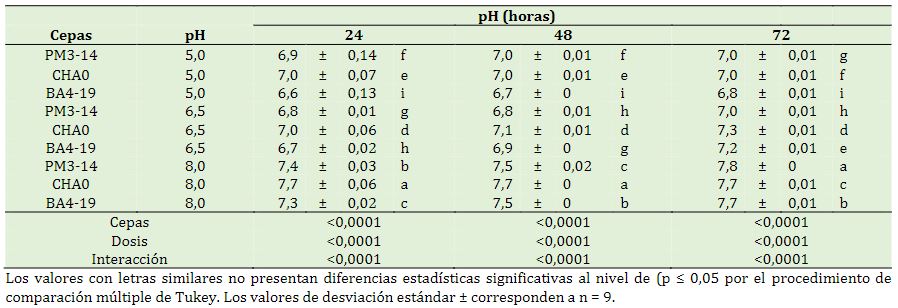
Las bacterias PM 3-14, BA 4-19 y CHA0 son conocidas por su versátil ruta metabólica, que les permite sobrevivir en ambientes contaminados y ajustar el pH mediante la producción de metabo-litos diversos. La respiración y fermentación bacteriana, que involucran la transformación de compuestos orgánicos, también pueden variar el estado del pH en medios de cultivo contaminados con herbicidas. La inoculación de Thermobifida cellulosilytica TB100 degrada el lindano en condiciones de pH 7 y 8. Sin embargo, el pH ácido y alcalino disminuyó la eficiencia de las especies bacterianas (Usmani et al., 2021). El medio de cultivo envenenado con glifosato influye en una disminución gradual del pH 6,8 a 5,3 para Bacillus tropicus GP1 y de 7,5 a 5,3 para Proteus mirabilis GP2 desde el día 0 al día 30 (Ibrahim et al., 2023).
Efecto del pH sobre la UFC/mL de las rizobac-terias
Se presentó significancia estadística en los factores en estudio y las interacciones (p < 0,0001) a las 48 y 72 h. Durante las primeras 24 h, PM3-14 con pH de 6,5 evidenció un mayor número de UFC/ml con 1,4E+07. A las 48 y 72 h, la cepa CHA0 demostró una mayor cantidad de células viables con (1,0E+08 y 1,1E+09), respectivamente (Tabla 6). Esto demuestra que las rizobacterias tienen la capacidad de desarrollar su mayor concentración celular en condiciones de pH ligeramente neutro.
Inoculación de rizobacterias sobre la germina-ción de Oryza sativa L. en suelo contaminado con pendimentalina
El porcentaje de germinación se vio afectado con dosis creciente de herbicidas. Las cepas BA4-19, CHA0 y PM3-14 con dosis de 75 y 150 mg/kg de pendimentalina aumentaron la germinación en comparación al control. La longitud de la raíz presentó un incremento sin aplicación de herbicida y rizobacterias (7,2 cm), sin embargo, en presencia de herbicida, las rizobacterias PM3-14 con dosis de 75 mg/kg y 150 mg/kg disminuyó la toxicidad con (2,3 y 1,3 cm) en comparación al control (0 cm). Se evidencia una mayor longitud del hipocótilo e índice de vigor (IDV) en suelos sin contaminación y aplicación de rizobacterias (3,1 cm y 102,5). A pesar de la acción de la rizobacteria para reducir la contaminación de pendimentalina, no se presentó una mejoría en la longitud del hipocótilo e IDV (Tabla 7). La bacteria PM 3-14 ejerce interacciones simbióticas en los estadios tempranos de germi-nación de arroz en suelos contaminado con pendi-mentalina donde posee un efecto bioestimulador sobre el incremento de biomasa del sistema radicular. Estas bacterias generan protección a las semillas de arroz de los efectos tóxicos de los herbicidas al degradar los herbicidas presentes en el suelo y además liberan subproductos que son beneficiosos para el crecimiento de las plantas (Sowunmi et al., 2020).
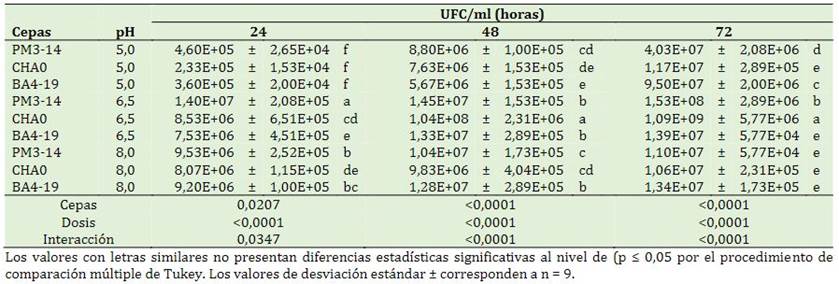
Tabla 7 Efecto de las rizobacterias sobre la dosis de pendimentalina sobre las variables morfológicas del arroz
Aplicaciones in vitro de Arthrobacter sp. AAC22 en semillas Avena sativa L. incrementa la longitud y mejora el índice de vigor a los 14 días post-incubación (Urseler et al., 2022). La inoculación con Bacillus amyloliquefaciens NRRU-TV11 ejerce un aumento sobre la altura de la planta, la longitud de las raíces, la biomasa y el índice de vigor de las plántulas de arroz en comparación con los controles no inoculados en suelos contaminados con clorpirifos (Saengsanga & Phakratok, 2023).
CONCLUSIONES
Las PGPR pueden tolerar 150 mg/L, presentando un mayor crecimiento celular en comparación a las demás dosis de pendimentalina. Se evidenció que las rizobacterias pueden reducir o aumentar su pH en dependencia de la condición que se encuentre, además la presencia de UFC/mL incrementa cuando existe un pH neutro. Las rizobacterias reduce levemente la toxicidad en germinación y crecimiento de arroz, sin dosis de pendimentalina mejora la germinación y las características morfológicas de las plantas. Es posible que las rizobacterias tengan un efecto especial, el estudio de bioprospección y diferentes condiciones abiótica a nivel de vivero podría ser una alternativa para seguir con futuros estudios sobre la descontaminación de la pendimentalina.

