










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Neuro-Psiquiatría
versión impresa ISSN 0034-8597
Rev Neuropsiquiatr vol.81 no.2 Lima abr. 2018
http://dx.doi.org/https://doi.org/10.20453/rnp.v81i2.3335
ARTÍCULO DE REVISIÓN
Alteraciones cognitivas espaciales y no espaciales relacionadas al sistema vestibular: una entidad subdiagnosticada
Space and non-space-related cognitive disturbances related to the vestibular system: an underdiagnosed entity
Manuel A. Gallardo-Flores 1,a
1 Laboratorio de Otoneurología, Clínica Anglo Americana. Lima Perú.
a Médico Otorrinolaringólogo
RESUMEN
El sistema vestibular permite estabilizar las imágenes visuales en la retina durante el desplazamiento y mantener el balance postural. Hay proyecciones desde los núcleos vestibulares hasta la corteza cerebral, específicamente a nivel del hipocampo y la región témporo-parietal, relacionadas a la llamada cognición espacial y a algunos procesos cognitivos no espaciales como atención y cálculo numérico. La presente es una revisión bibliográfica de areas de estudio e investigación en relación a estos procesos. En los sistemas de búsqueda Pub Med y Google Académico, se encontraron 44 artículos, de los cuales dos fueron capítulos de libros de texto. Se elaboran conclusiones pertinentes.
PALABRAS CLAVE: Cognición espacial, cognición no espacial, sistema vestibular, corteza vestibular.
SUMMARY
The vestibular system allows the stabilization of visual images in the retina during displacement, thus contributing to the maintenance of postural balance. There are projections from the vestibular nuclei to the cerebral cortex, specifically hippocampus and the temporo-parietal region, related to the so-called spatial cognition and to some non- spatial cognitive processes such as attention and numerical calculation. A literature review on study and research areas related to these processes is presented. Academic Google and PubMed search engines were utilized with a total of 44 articles found, two of which were textbook chapters. Pertinent conclusions are elaborated.
KEY WORDS: Spatial cognition, non-spatial cognition, vestibular system, vestibular cortex.
INTRODUCCIÓN
El desarrollo del sistema vestibular sigue un planeamiento genético y la influencia de estímulos externos como la gravedad, la cual juega un papel fundamental para su desarrollo (1). El tamaño del laberinto dependerá del medio donde se desenvuelvan los seres vivos, de tal manera que aquellos animales que vivan en lugares altos, como los árboles, y de gran desplazamiento tendrá canales semicirculares de
mayor tamaño (2, 3). El desarrollo de la verticalidad, el cual llevará al desarrollo de la bipedestación depende del crecimiento de los canales semicirculares verticales (4). Los cambios evolutivos que finalmente dieron origen al Homo erectus se llevaron a cabo en el neocortex, cerebelo y el complejo vestibular (5,6). Este amplio desarrollo de estructuras del sistema nervioso, especialmente el neocortex, llevó a un sistema vestibular que a través de sus conexiones corticales permitiera establecer relaciones con el medio que rodeaba a los seres humanos, desarrollándose procesos cognitivos fundamentales como la memoria espacial, importante para el retorno del hombre primitivo hacia las cuevas después de un largo recorrido en busca de los alimentos.
Hemos observado en muchos pacientes con afección vestibular déficits cognitivos en la memoria, atención, velocidad del procesamiento mental y en cálculos matemáticos que antes del desarrollo del cuadro vestibular no tenían, los cuales no son usualmente identificados llevándonos a un sub- diagnóstico.
Esta revisión tiene por objeto poner al día algunos conocimientos no tomados en cuenta como las vías vestíbulo-corticales, la importancia del hipocampo en la memoria y el rol de las aferencias vestibulares hacia esta estructura, y sobre todo lo que se conoce hasta hoy como deterioro cognitivo por déficits vestibulares.
Sistema vestibular
Es un sistema filogenéticamente muy antiguo. La evolución cambió las características anatómicas y funcionales del laberinto, y el crecimiento del neocortex, cerebelo y sistema vestibular permitió la bipedestación y la estabilidad postural, esto a su vez permite la capacidad de exploración e interacción con el medio ambiente. Sin esto se disminuye las posibilidades de supervivencia en un ambiente hostil (7).
El sistema vestibular permite estabilizar la imagen del mundo que nos rodea en la retina durante el desplazamiento, y mantener el balance postural. El órgano receptor se encuentra en el hueso temporal rodeado por la cápsula ótica. El laberinto posterior o vestibular está constituido por tres canales semicirculares dispuestos perpendicularmente entre sí en forma ortogonal. Cada canal semicircular tiene en uno de sus extremos una dilatación en la que se encuentra un acelerómetro angular, sensible a las aceleraciones angulares de la cabeza. Estos canales están unidos a una estructura sacular, el utrículo, en el cual se encuentra la mácula utricular, cuya función es percibir aceleración lineal y la gravedad. Estos acelerómetros angulares y lineales son estimulados continuamente durante el desplazamiento, y el impulso nervioso generado va hacia los núcleos vestibulares que se encuentran en el piso del cuarto ventrículo. Hacia estos núcleos también llega información visual y somatosensorial. Desde aquí, a través del fascículo longitudinal medio, asciende un impulso hacia los núcleos oculomotores para el reflejo vestíbulo-ocular y estabilizar la imagen en la retina. En forma descendente va hacia las neuronas motoras que se encuentran en las astas anteriores de la médula y mantener un tono muscular de reposo que permite la estabilidad postural o balance (8, 9).
La corteza vestibular
Se ha establecido a través de estudios con trazadores neuronales, electrofisiológicos, inmunohistoquímicos, con resonancia funcional y conductuales, áreas corticales vestibulares ubicados en la región temporal, parietal e hipocampal (10-13).
Desde los núcleos vestibulares hay proyecciones hacia el tálamo, específicamente los núcleos ventral posterolateral, ventral posteromedial y ventral posteroinferior. Desde los núcleos talámicos hay vías que van hacia la corteza cerebral. Las áreas corticales vestibulares son: el área 2v en la punta del surco intraparietal, área 3v en el surco central, la corteza vestibular parietoinsular y el área 7 en el lóbulo parietal inferior (8, 9, 14) (tabla 1).
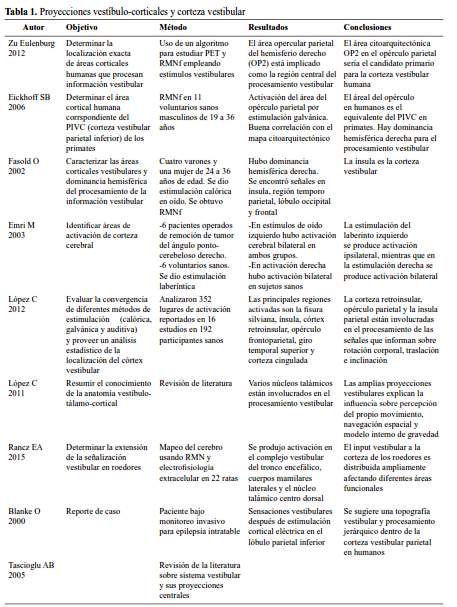
En roedores las aferencias vestibulares se distribuyen ampliamente por la corteza en diferentes áreas funcionales, no existiendo un área cortical vestibular primaria y la integración de la información ocurre en áreas que median diversas funciones sensori- motoras (15). Previa a la llegada a la corteza, las aferencias que parten desde los núcleos vestibulares superior y medial, vía fascículo longitudinal medio y pedúnculo cerebelar superior, llegan al núcleo ventral postero lateral del tálamo, así como al núcleo ventral intermedio. Estos núcleos son los principales relevos de información propioceptiva y cutánea a áreas primarias y corteza somatosensorial secundaria (16).
En animales se ha identificado a la corteza vestibular parieto insular (CVPI) como la “corteza vestibular”, la cual correspondería a la corteza temporo-parieto- insular y retroinsular, corteza parietal, corteza frontal, corteza cingular y tres estructuras subcorticales (tálamo, ganglio basal y cerebelo) en los seres humanos (16). En un estudio realizado en sujetos voluntarios sanos y pacientes que serían sometidos a cirugía de schwannoma vestibular derecho, encontraron que ante estimulación del oído izquierdo hubo gran activación bilateral del giro precentral y post-central, la úvula, el área SII y la corteza retrorinsular, mientras que la activación del oído derecho de los sujetos sanos generó activación de las mismas áreas, pero con gran activación del hemisferio izquierdo (17). Esta región constituye un área sensorial multimodal (18). Otro estudio atribuye al área OP2 como el equivalente de la CVPI de los primates no humanos (19).
La corteza insular posterior constituiría entonces el homólogo humano del CVPI de los animales, y que además estaría involucrada en la percepción de verticalidad y del movimiento propio. En la activación del cortex vestibular, de las áreas visuales sensibles al movimiento y áreas oculo-motoras se muestra una significativa dominancia del hemisferio cerebral derecho (20).
Por lo tanto, no existe un área vestibular primaria que solo procese información vestibular, es más bien una región multimodal donde se procesa además de información vestibular aferencias somatosensoriales y visuales.
Rol del hipocampo
En 1957 Scoville reportó el caso de un hombre que sufría de epilepsia intratable y con la finalidad de reducir sus ataques fue sometido a una resección temporal medial bilateral con el objeto de interrumpir el flujo eléctrico de los ataques a través de su cerebro (21); este paciente experimentó una severa amnesia anterógrada, esto es no podía formar memoria de nuevos eventos, experiencias o conocimiento semántico, sin embargo su memoria de trabajo estaba intacta y podía mantener nueva información por períodos cortos. Se pensó que la pérdida de la memoria estaría relacionada con la pérdida del área medial del lóbulo temporal, sugiriendo que esta zona era importante para un tipo particular de memoria, sin otro déficit mayor en funciones perceptuales, cognitivas o intelectuales. Por más de treinta años continuaron los estudios en este paciente, los cuales revelaron diferencias entre la memoria declarativa y procedimental. También elaboraron distinciones entre memoria a corto y largo plazo (22).
Con este reporte se dio inicio al desarrollo de una teoría neural de la memoria, en el cual “el hipocampo ha mantenido una posición central en el desarrollo de las teorías psicológicas de la memoria humana, normal y anormal, y animal” (23).
Un trabajo que resultó crucial en la comprensión de la memoria espacial fue el que realizó O’Keefe et al., en el cual determinaron que ratas con daño en
el hipocampo tenían pobre desempeño en tareas de memoria espacial. Asumieron que se debía a pérdida del sistema neural que provee al animal de un mapa cognitivo o espacial de su ambiente (24). Poncet et al., afirman que estas “place cells” hipocampales forman parte de un sistema neural integrado para la navegación espacial y parte importante de este sistema son las “head direction cells” (25). Sugiere además que se necesita de un elemento esencial que es la representación de la meta, lo cual es necesario para la planificación de una tarea, y esto se codificaría a nivel de la corteza prelímbica/infralímbica de la corteza prefrontal medial. Propone una red neural responsable de la navegación espacial en el cual el hipocampo permite el reconocimiento del lugar, la corteza prefrontal sería la responsable de codificar la localización de la meta y la planificación de la ruta se realizaría en la región infralímbica/límbica de la corteza prefrontal medial.
Se ha establecido que la estimulación vestibular ya sea eléctrica o por rotación puede generar actividad ritmo teta en el electroencefalograma, y que la inactivación del sistema vestibular reduce esta actividad eléctrica significativamente lo que a su vez afectaría el funcionamiento de las “place cells” deteriorando el aprendizaje espacial y la memoria (26,27). Aparentemente la estimulación eléctrica de alta frecuencia del laberinto derecho generaría respuestas hipocampales bilaterales (28).
La región dorsal del hipocampo recibe información polimodal altamente procesada y que procede de diferentes áreas corticales, mientras que la región ventral está ligada a estructuras subcorticales como la amígdala y el eje hipotálamo-pituitaria-adrenal. Es por esto que el papel más importante del hipocampo dorsal son las tareas de memoria y navegación espacial, mientras que la el hipocampo anterior está asociado al procesamiento de las emociones (29).
Las tareas espaciales, de memoria allocéntrica que tiene su asiento en el hipocampo, están relacionadas directamente con la edad. En sujetos sanos sin deterioro vestibular ni deterioro cognitivo mayores de 70 años hay deficiencia en memoria espacial, dependiente de disminución en el volumen del hipocampo (30,31).
Cognición espacial y sistema vestibular
Como se ha descrito anteriormente el sistema vestibular no solo tiene como funciones estabilizar la mirada y mantener el balance postural sino que también participa en cognición espacial, representación corporal y procesos afectivos (32,33), aunque también algunos procesos cognitivos no espaciales también se ven afectados (34) (tabla 2).
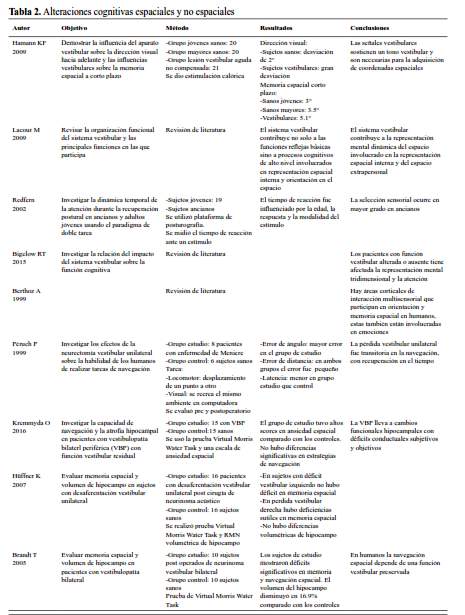
Están descritos dos tipos de memoria espacial: egocéntrico, esto es el individuo con las señales vestibulares y propioceptivas como punto de referencia y allocéntrica, en la cual las señales externas son el punto de referencia (29).
Se han realizado una serie de estudios con la finalidad de evaluar el impacto de las lesiones vestibulares periféricas sobre cognición espacial. En sujetos con lesión vestibular aguda no compensada se encontró deterioro en memoria espacial y alteración en la dirección visual hacia adelante (35). En pacientes con enfermedad de meniere que fueron sometidos a neurectomía vestibular se encontró deterioro de la memoria espacial, pero esta se recuperó parcialmente un mes después lo que hace pensar que los déficits vestibulares unilaterales pueden generar deterioro transitorio en navegación espacial (36). En sujetos con déficit vestibular periférico bilateral debido a neurectomía vestibular bilateral por schwannoma vestibular, se encontró además de déficit en tareas cognitivas espaciales disminución en el volumen de hipocampo hasta en 16,9% (37). En deterioros vestibulares periféricos bilaterales con función vestibular residual se encontró déficits sutiles en tareas de memoria y navegación espacial, así como atrofia del hipocampo medio y del parahipocampo posterior (38). Estos dos últimos estudios utilizaron una prueba virtual de memoria y navegación espacial que es una adaptación de la tarea de Morris en agua, utilizada en roedores. También se ha demostrado que en pacientes con deterioro vestibular periférico hay alteración en las tareas de transformación mental de los objetos (39). Hüfner reportó que en lesiones vestibulares unilaterales del laberinto derecho había peor desempeño en la prueba de memoria y navegación espacial virtual de Morris que en los déficits de laberinto izquierdo (40).
Cognición no espacial y sistema vestibular
Se ha demostrado que lesiones hipocampales producen, además de afecciones en tareas de memoria espacial, deterioros en tareas no espaciales tales como aspectos emocionales especialmente ansiedad (29). Como se ha mencionado anteriormente la región dorsal del hipocampo está relacionada a procesos espaciales, mientras que la región anterior está asociada al procesamiento de las emociones. Entonces existe una
superposición de las funciones de estructuras cerebrales involucradas en memoria y orientación espacial y las involucradas en las emociones, encontrándose ansiedad, agorafobia y pánico en sujetos con vértigo. Esto es considerado como una manifestación de la distorsión de la habilidad integradora multisensorial (41).
El sistema vestibular tiene también influencia sobre atención y procesos cognitivos, como la habilidad visuo-espacial, de tal manera que una hipofunción vestibular genera incremento en la atención dirigida a mantener la estabilidad con la consiguiente disminución de los recursos disponibles para tareas cognitivas (42). En pruebas de doble tarea en las cuales se pedía mantener la postura sobre una plataforma de posturografía a la vez que se realizaba cálculos aritméticos se encontró interferencias entre realizar tareas cognitivas y control postural, debido a un incremento en el proceso de atención (43).
En 1989 ya se había reportado que en lesiones periféricas como fístula perilinfática, había síntomas cognitivos como emocionales, déficits de memoria y atención, ansiedad y depresión (44). También se ha demostrado que el cálculo aritmético estuvo afectado en situaciones de doble tarea (45), asumiéndose que esta discalculia en sujetos con trastornos vestibulares estaría relacionada a déficits de memoria espacial (46).
CONCLUSIONES
El sistema vestibular permite estabilizar la mirada durante el desplazamiento y el mantenimiento de la postura, lo cual permite la supervivencia en un ambiente hostil en los animales. Existen además proyecciones hacia centros corticales superiores, fundamentalmente temporo-parietales, desde los núcleos vestibulares. Estas áreas corticales son fundamentalmente de integración multimodal de información visual, somatosensorial, cervical y vestibular. Estas regiones están involucradas principalmente en procesos cognitivos espaciales, de memoria egocéntrica y allocéntrica, esta última asentada en el hipocampo. También hay manifestaciones de compromiso emocional como ansiedad y fobias; alteraciones en la atención y cálculos de números, esto último debido a déficits espaciales.
Se nos abre entonces un campo muy grande de estudio donde encontraremos explicación a muchas manifestaciones, que si bien es cierto son algo sutiles si no les prestamos la debida atención, que afectan la calidad de vida de nuestros pacientes.
REFERENCIAS BIBLIOGRAFICAS
1. Jamon M. The development of vestibular system and related function in mammals: Impact of gravity. Front Integr Neurosci. 2014; 8: 11. Doi:10.3389/fnint.2014.00011 [ Links ]
2. Solntseva GN. Early embryogenesis of the vestibular apparatus in mammals with different ecologies. Aquatic Mammals. 2002; 28(2):159-169. [ Links ]
3. Boistel R, Honel A, Lebrun R, Daghfous G, Tafforean P. Shake rattle and roll: the bony labyrinth and aerial descent in squamates. Integr Comp Biol.2011; 51(6):957-68. doi: 10.1093/icb/icr034 [ Links ]
4. Spoor F. The semicircular canal system and locomotor behavior, with special reference to hominin evolution. Courier Forschungs institut Senckenberg. 2003; 243:93-104. [ Links ]
5. Barton RA, Harvey PH. Mosaic evolution of brain structure in mammals. Nature. 2000; 405 (29): 1055-1058. [ Links ]
6. Whiting B, Barton RA. The evolution of the cortico- cerebellar complex in primates: anatomical connections predict patterns of correlated evolution. J Hum Evol. 2003; 44(1):3-10. [ Links ]
7. Wade MG, Jones G . The role of vision and spatial orientation in the maintenance of posture. Phys Ther.1997; 77: 619-628. [ Links ]
8. Highstein SM. How does the vestibular part of the inner ear works? In: Baloh RW, Halmagyi GM. (Edit). Disorders of the vestibular system. New York: Oxford University Press; 1996. [ Links ]
9. Leigh RJ, Zee DS. A survey of eye movements: characteristics and teleology. In: Leigh RJ, Zee D. (Edit). The neurology of eye movements. New York: Oxford University Press; 1999. [ Links ]
10. Zu-Eulenburg P, Caspers S, Roski C, Eickhoff SB.Meta-analytical definition and functional connectivityof the human vestibular cortex. Neuro Image. 2012;60: 162-169. [ Links ]
11. Blanke O, Perrig S, Thut G, Landis T, Seeck M. Simple and complex vestibular responses induced by electrical cortical stimulation of the parietal cortex in humans. J Neurol Neurosurg Psychiatry. 2000; 69:553-556 [ Links ]
12. López C, Blanke O. The talamocortical vestibular system in animals and humans. Brain Research Reviews. 2011; 67: 119-146. [ Links ]
13. Hitier M, Besnard S, Smith PF. Vestibular pathways involved in cognition. Front Integr Neurosci.2014;8:59. doi: 10.3389/fnint.2014.00059 [ Links ]
14. Tascioglu AB. Brief review of vestibular system anatomy and its higher order proyections. Neuroanatomy. 2005; 4:24-27. [ Links ]
15. Rancz EA, Moya J, Drawitsch F, Brichta AM, Canals S, Margrie TW. Widespread vestibular activation of the rodent cortex. J Neurosci. 2015; 35 (15): 5926-5934. [ Links ]
16. Lopez C, Blanke O, Mast FW. The human vestibular cortex revealed by coordinate-based activation likelihood estimation meta-analysis. Neuroscience.2012; 212: 159-179. [ Links ]
17. Emri M, Kisely M, Lengyel Z, Balkay L, Marian T. Cortical proyection of peripheral vestibular signaling. J Neurophysiol. 2003; 89: 2639-2646. [ Links ]
18. Fasold O, Von Brevem M, Kuhberg M, Ploner ChJ, Villringer A, Lempert T, et al. Human vestibular cortex as identified with caloric stimulation in functional resonance imaging. Neuroimage. 2002; 17: 1384-1423. Doi: 10.1006/nimg.2002.1241. [ Links ]
19. Eickhoff SB, Weiss PH, Amunts K, Fink GR, Zilles K. Identifying human parieto-insular vestibular cortex using fMRI and cytoarchitectonic mapping. Human Brain Mapping. 2006; 27: 611-621. [ Links ]
20. Brandt T, Dieterich M. The vestibular cortex: its locations, functions and disorders. Ann NY Acad Sci.1999; 871: 293-312. Doi: 10.1111/J.1749-6632.1999.tb09193.x [ Links ]
21. Scoville WH, Milner B. Loss of recent memory after bilateral hippocampal lesions. J Neurol Neurosurg Psychiat. 1957; 20: 11-21. [ Links ]
22. Pickren WE, Rutherford A. A history of modern psychology in context. New Jersey: John Wiley & Sons Inc; 2010. [ Links ]
23. Good M. Spatial memory and hippocampal function:where are we now? Psicologica. 2002, 23: 109-138. [ Links ]
24. O’Keefe J, Dostrovsky J . The hippocampus as a spatial map: Preliminary evidence from unit activity in the freely-moving rat. Brain Research. 1971; 34:171-175. [ Links ]
25. Poucet B, Lenck-Santini PP, Hok V, Lave E, Banquet JP, Gaussier P, Muller RU. Spatial navigation and hippocampal place cell firing: The problem of goal encoding. Rev Neurosci. 2004;15(2):89-107. [ Links ]
26. Aitken P, Zheng Y, Smith PF. The modulation of hippocampal theta rhythm by the vestibular system. J Neurophysiol. 2018; 119 (2): 548-562. Doi: 10.1152/jn.00548.2017 [ Links ]
27. Stackman RW, Clark A, Taube JS. Hippocampal spatial representations require vestibular input. Hippocampus. 2002; 12: 291-303. [ Links ]
28. Cuthbert PC, Gilchrist DP, Hicks SL, Mac Dougall HG, Curthoys IS. Electrophysiological evidence for vestibular activation of the guinea pig hippocampus. Neuro Report. 2000; 11: 1443-1447. [ Links ]
29. Bannerman DM, Sprengel R, Sanderson DJ, Mc Hugh SB, Rawlins JN, Monyer H , et al. Hippocampal synaptic plasticity, spatial memory and anxiety. Nat Rev Neurosci. 2014;15(3):181-92. doi: 10.1038/nrn3677 [ Links ]
30. Gazova I, Laczo J, Rubinova E, Mokrisova I Hynsicova E, Andel R, et al. Spatial navigation in young versus older adults. Front Aging Neurosci.2013; 5:94. doi: 10.3389/fnagi2013.00094 [ Links ]
31. Konishi K, Bohbot VD . Spatial navigational strategies correlate with gray matter in the hippocampus of healthy older adults tested in a virtual maze. Front Aging Neurosci. 2013; 5: 1-8. Doi: 10.3389/fnagi.2013.00001 [ Links ]
32. Mast FW, Preuss N, Hartmann M, Grabherr L. Spatial cognition, body representation and affective processes: the role of vestibular information beyond ocular reflexes and control of postural. Front Aging Neurosci. 2014;8(11): 1-14. [ Links ]
33. Lacour M. Locomotion and spatial navigation in vestibular pathology. An Gerontol. 2009; 2 (2): e38-e45. [ Links ]
34. Smith PF, Zheng Y, Horii A, Darlington CL. Does vestibular damage cause cognitive dysfunction in humans? J Vestib Res. 2005;15(1):1-9. [ Links ]
35. Hamann KF, Weiss U, Ruile A. Effects of acute vestibular lesions on visual orientation and spatial memory, shown for the visual straight ahead. Ann N YAcad Sci. 2009; 1164: 305-308. Doi: 10.1111/j.1749-6632.2009.03867.x [ Links ]
36. Peruch P, Borel L, Gaunet F, Thinus-Blanc G, Magnan J, Lacour M. Spatial performance of unilateral vestibular defective patients in nonvisual versus visual navigation. J Vestib Res. 1999;9(1):37-47. [ Links ]
37. Brandt T, Schautzer F, Hamilton DA, Brüning R, Markowitsch HJ, Kalla R, et al. Vestibular loss causes hippocampal atrophy and impaired spatial memory in humans. Brain. 2005; 128: 2732-2741. [ Links ]
38. Kremmyda O, Hüfner K, Flanagin VL, Hamilton DA, Linn J, Strupp M, et al. Beyond dizziness: virtual navigation, spatial anxiety and hippocampal volumen in bilateral vestibulopathy. Front Hum Neurosci.2016; 10: 139. doi: 10.3389/fnhum.2016.00139 [ Links ]
39. Péruch P, Lopez C, Redon-Zouiteni C, Escoffier G, Zeitoun A, Sanjuan M,et al. Vestibular information is necessary for maintaining metric properties of representational space: evidence from mental imagery. Neuropsychologia. 2011;49(11):3136-44. doi: 10.1016/j.neuropsychologia.2011.07.026 [ Links ]
40. Hüfner K, Hamilton DA, Kalla R, Stephan T, Glasauer S, Ma J, et al. Spatial memory and hippocampal volume in humans with unilateral vestibular deafferentation. Hippocampus. 2007; 17:471-485. [ Links ]
41. Berthoz A, Viaud-Delmo I. Multisensory integration in spatial orientation. Curr Opin Neurobiol. 1999;9(6):708-12. [ Links ]
42. Bigelow RT, Agrawal Y. Vestibular involvement in cognition: visuospatial ability, attention, exectutive function, and memory. J Vestib Res. 2015;25(2):73-89. doi: 10.3233/VES-150544 [ Links ]
43. Redfern M, Müller MLTM, Jennings JR, Furman JM. Attentional dynamics in postural control during perturbations in young and older adults. J Gerontol A Biol Sci Med Sci. 2002;57(8):B298-303. [ Links ]
44. Grimm R, Hemenway WG, Lebray PR, Black FO. The perilymph fistula síndrome defined in mild head trauma. Acta Otolaryngol Suppl. 1989;464:1-40. [ Links ]
45. Yardley L, Papo D, Bronstein A, Gresty M, Garder M. Attentional demands of continuously monitoring orientation using vestibular information. Neuropsychologia. 2002; 40: 373-383. [ Links ]
46. Smith PF. Dyscalculia and vestibular function. Medical Hypotheses. 2012; 79: 493-496 [ Links ]
Correspondencia:
Manuel A. Gallardo-Flores
Correo electrónico: mgallardoflores@yahoo.es
Declaro no haber recibido financiamiento ni tener conflicto de intereses en la elaboración de este artículo.
Recibido: 18/03/2018
Aceptado: 08/05/2018

