










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAnales de la Facultad de Medicina
versão impressa ISSN 1025-5583
An. Fac. med. v.68 n.4 Lima dez. 2007
TRABAJOS ORIGINALES
Caracterización de las proteínas totales de tres ecotipos de maca (Lepidium peruvianum G. Chacón), mediante electroforesis unidimensional y bidimensional
Characterization of three maca (Lepidium peruvianum G. Chacon) echotypes total proteins by unidimensional y bidimensional electrophoresis
Mario Monteghirfo 1, Armando Yarleque-Chocas 2
1 Centro de Investigación de Bioquímica y Nutrición. Facultad de Medicina, Universidad Nacional Mayor de San Marcos. Lima, Perú.
2 Laboratorio de Biología Molecular. Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Lima, Perú.
Resumen
Objetivo: Caracterizar las proteínas solubles que se encuentran en la raíz del Lepidium peruvianum G. Chacón, maca, mediante electroforesis unidimensional y electroforesis bidimensional. Diseño: Estudio de tipo observacional y transversal. Lugar: Centro de Investigación de Bioquímica y Nutrición Alberto Guzmán Barrón. Facultad de Medicina, Universidad Nacional Mayor de San Marcos. Lima, Perú. Materiales: Raíces de Lepidium peruvianum G. Chacón maca de los ecotipos blanco, amarillo y morado, procedentes de Junín que fueron obtenidas a través de la Universidad Nacional del Centro del Perú. Métodos: La extracción de las proteínas totales solubles se realizó con una solución antioxidante, seguida de electroforesis unidimensional y bidimensional para su caracterización. Principales medidas de resultados: Número de proteínas solubles, peso molecular de las proteínas y puntos isoelectricos de las proteínas más abundantes. Resultados: El análisis electroforético unidimensional mostró predominio de 2 proteínas (72% de las proteínas solubles totales). Una de 22,5 kDa, denominada en el presente trabajo macatina (51% de la proteína total); la otra de 17,0 kDa (21% de la proteína soluble total). El mapa electroforético bidimensional mostró que tanto la macatina como la proteína de 17,0 kDa son básicas y presentan 3 isómeros de carga que se distribuyen en un rango de punto isoeléctrico (pI) de 7,1 a 8,2. Conclusiones: Las proteínas solubles mostraron un patrón electroforético complejo, siendo la macatina la proteína más abundante.
Palabras clave: Lepidium; electroforesis; electroforesis en gel de poliacrilamida; electroforesis gel bidimensional.
Abstract
Objective: To characterize soluble proteins present in Lepidium peruvianum G. Chacon maca root by both unidimensional and bidimensional electrophoresis. Design: Observational and cross-sectional type study. Setting: Alberto Guzman Barron Biochemistry and Nutrition Research Center, Faculty of Medicine, Universidad Nacional Mayor de San Marcos, Lima, Peru. Materials: Roots of Lepidium peruvianum G. Chacon maca white, yellow and purple ecotypes, collected in Junin and obtained from Universidad Nacional del Centro del Peru. Methods: Soluble total proteins extraction with an antioxidant solution and characterization by both unidimensional and bidimensional electrophoresis were done. Main outcome measures: Soluble protein number, molecular weight and most abundant proteins isoelectrical points. Results: The unidimensional electrophoretic analysis showed predominance of two proteins (72% of total soluble proteins): one with 22,5 kDa denominated in the present work as macatina (51% of the total protein) and another with 17,0 kDa (21% of total soluble protein). The bidimensional electrophoretic map sample showed that both macatina and the 17,0 kDa protein are basic and display three charge isomers distributed in an isoelectric point (pI) ranging from 7,1 to 8,2. Conclusions: The soluble proteins ecotypes studied showed a complex electrophoretical pattern, being macatina the most abundant protein.
Key words: Lepidium; electrophoresis; electrophoresis, polyacrylamide gel; electrophoresis, gel, two-dimensional.
INTRODUCCIÓN
Las raíces y tubérculos andinos constituyen un reservorio de variabilidad genética de gran biodiversidad para el mundo. Crecen en los Andes, a altitudes de 2 400 a 4 800 metros, y por consiguiente tienen gran potencial para su introducción en otras áreas montañosas donde los cultivos del viejo mundo no se adaptan ( 1 ). Sin embargo, las raíces y tubérculos andinos no han sido tomados en cuenta en su real magnitud y han sido poco estudiados a nivel biológico y bioquímico ( 2,3 ).
Los cultivos andinos son muy diversos e incluyen granos, leguminosas, tubérculos, raíces, frutos y constituyen una fuente importante de proteínas, principalmente en la dieta vegetariana de los pobladores andinos ( 4 ). Sin embargo, actualmente la producción y consumo de muchas de esas especies están disminuyendo considerablemente, debido a la influencia de los hábitos alimenticios de los países desarrollados y a la falta de conocimientos de la calidad nutritiva de estos cultivos andinos.
Varios estudios han sido realizados, que han contribuido a la investigación del valor nutritivo de las proteínas, principalmente de los granos andinos Chenopodium quinoa quinua, Chenopodium pallidicauli kañiwa, Amaranthus caudatus kiwicha y Lupinus mutabilis tarwi ( 5 ). Existen muchos estudios químico-nutricionales en diferentes ecotipos de maca, en su gran mayoría con la denominación botánica de Lepidium meyenii Walp, para conocer el porcentaje de nutrientes de esta planta ( 6 ). En el presente trabajo se utilizará la denominación Lepidium peruvianum G. Chacón, debido a que los ecotipos analizados provienen del Departamento de Pasco. Otros trabajos etnobotánicos y fitoquímicos encuentran que la maca tiene propiedades fertilizante, afrodisíaca, revitalizante y reguladora ( 7 ). Sin embargo, es muy escasa la literatura científica en la cual se ha estudiado las características bioquímicas de las proteínas solubles presentes en la raíz de Lepidium peruvianum G. Chacón maca, a pesar de que su consumo ha aumentado considerablemente en los últimos años ( 8 ). Asimismo, se ha caracterizado las proteínas de otros cultivos andinos, como Ullucus tuberosus olluco y Tropeolum tuberosum mashua, mediante técnicas de electroforesis unidimensional y bidimensional ( 9 ).
En cuanto a estudios realizados con las proteínas de maca, estos son escasos. Existe un estudio en cual se comunica la composición química de los ecotipos amarillo, rojo y negro de Lepidium meyenii Walp ( 10 ). Se señala un contenido de proteínas totales para el ecotipo amarillo de 17,9 g%, el ecotipo rojo 17,2 g% y el ecotipo negro 16,3 g%. En otro estudio, encuentran un contenido de 13 g% para el ecotipo amarillo y 12,8 g% para el ecotipo morado ( 11 ). La fluctuación del contenido de proteína depende de la variedad y de la fertilidad del suelo ( 12 ). El presente trabajo tiene como objetivo caracterizar las proteínas solubles que se encuentran en la raíz de la maca, mediante electroforesis unidimensional y electroforesis bidimensional.
MÉTODOS
Se realizó un estudio de tipo observacional y transversal, utilizando para ello las raíces de Lepidium peruvianum G. Chacón maca de los ecotipos blanco, amarillo y morado procedentes de Junín. Las raíces fueron obtenidas a través de la Universidad Nacional del Centro del Perú.
Para la extracción de las proteínas solubles totales de las raíces de maca se utilizó una máquina extractora y, para evitar la oxidación, se adicionó una solución antioxidante compuesta de sulfito de sodio al 20% y bisulfito de sodio al 15%. El extracto fue centrifugado a 3 000 rpm, por 10 minutos, con la finalidad de remover el almidón y demás restos y el sobrenadante se guardó en congelación a una temperatura de –20°C hasta el momento de su procesamiento.
Las muestras de proteínas solubles totales de los tres ecotipos de maca fueron analizadas utilizando el sistema discontinuo SDS-PAGE ( 13 ). Se tomó 1 mL de sobrenadante de maca y se precipitó con 4 volúmenes de acetona fría, luego se centrifugó a 3000 rpm por 5 minutos; se descartó el sobrenadante y el sedimento se resuspendió en buffer de muestra (Tris – HCl pH 6,8 , 0,5M, SDS 10%, glicerol 20% y 2–mercaptoetanol 10%); se llevó a un volumen final de 250 µL, se homogenizó completamente y luego se centrifugó a 12 000 rpm, durante 5 minutos. Se calentó el sobrenadante en baño maría, por 3 minutos. Se enfrió con agua, luego se agregó 5 µL de azul de metileno. Las muestras fueron aplicadas en un gel de poliacrilamida al 15% (SDS – PAGE). La electroforesis se realizó en una cámara electroforética de tipo vertical Migthy Hoefer durante 45 minutos, para lo cual se usó una corriente constante de 20 miliamperios. Una vez terminada la corrida electroforética los geles fueron teñidos por 20 minutos con una solución colorante de azul de Coomassie R-250 al 0,1% metanol: agua: ácido acético (5:5:1) y se decoloró por difusión en una solución decolorante de metanol: ácido acético: agua (5:7:88). Como marcadores de peso molecular se usó albúmina de bovino 66 kilodaltons (kDa), ovoalbúmina (45 kDa), gliceraldehido-3-fosfato deshidrogenasa (36 kDa), anhidrasa carbónica (29 kDa), tripsinógeno de bovino (24 kDa), inhibidor de tripsina (20,1 kDa) y lacto albúmina (14,2 kDa). Los geles teñidos fueron analizados en un densitómetro, para la determinación de las proporciones de las proteínas de cada uno de los ecotipos de maca.
Las muestras de proteínas solubles totales de raíz de maca fueron analizadas utilizando el sistema de IEF/SDS-PAGE ( 14 ). Se pesó 3 mg del precipitado de acetona y se resuspendió en 100 µL de buffer de solubilización 9M úrea, 5% NP40, 1% 2- mercaptoetanol, 2% anfolitos pH 3-10 y se los separó en la primera dimensión mediante isoelectroenfoque en tubo con gradiente de pH, para lo cual se utilizó anfolitos con rango de pH de 3 a 10. Las condiciones de corrida fueron de 300 voltios durante 40 minutos. En la segunda dimensión, se empleó el método electroforético SDS/PAGE. La concentración del gel de apilamiento fue de 5% y la concentración del gel de separación de 15%. Las condiciones de corrida fueron de 20 mA para el gel de apilamiento y 40 mA para el gel de separación. Los geles fueron coloreados con azul de Coomassie R-250.
RESULTADOS
Respecto al análisis electroforético, la Figura 1 muestra los patrones electroforéticos unidimensionales obtenidos mediante sistema de gel denaturante SDS-PAGE, de la proteína soluble total de los 3 ecotipos de maca; dichos patrones electroforéticos son muy similares. La línea 1 determina el estándar de peso molecular para proteínas, la línea 2 muestra el patrón electroforético de las proteínas del ecotipo amarillo, las líneas 3 y 4 exhiben los patrones electroforéticos de los ecotipos morado y blanco, respectivamente. Para todos los ecotipos, se observa que los polipéptidos más abundantes son los de peso molecular bajo. El polipéptido más prominente en los 3 ecotipos analizados tiene un peso molecular aproximado de 22,5 kDa; el segundo polipéptido más abundante presenta un peso molecular de 17,0 kDa; el tercer polipéptido más abundante tiene un peso molecular de 15,5 kDa y luego le siguen los polipéptidos de 10,2 kDa y 5,3 kDa, que se encuentran en menor cantidad (Figura 2).
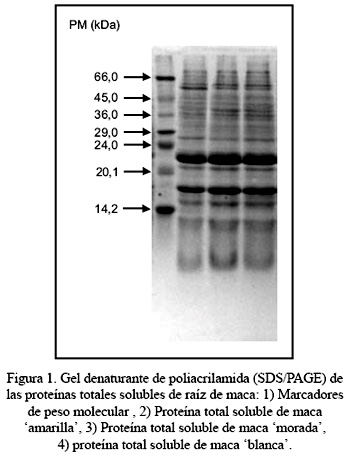
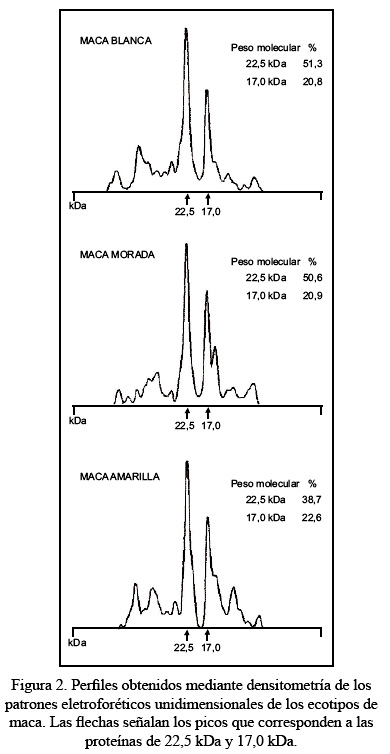
La Figura 2 además nos muestra los resultados del análisis densitométrico de los 3 ecotipos de maca. En el ecotipo blanco la proteína más abundante de 22,5 kDa constituye el 51,3% de la proteína soluble total; en el ecotipo morado, el 50,6%, y el ecotipo amarillo, el 38,7%. La segunda proteína más abundante de 17,0 kDa constituye el 20,8% en el ecotipo blanco, 20,9% en el ecotipo morado y 22,6% en el ecotipo amarillo.
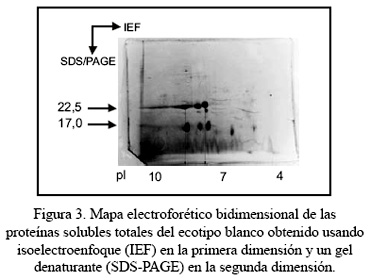
La Figura 3 muestra el mapa electroforético bidimensional de la proteína total del ecotipo blanco obtenido usando isoelectroenfoque (IEF) en la primera dimensión y un gel denaturante (SDS-PAGE) en la segunda dimensión. El mapa bidimensional no es muy complejo, pero revela características interesantes de las proteínas más abundantes de maca. El mapa bidimensional muestra que los puntos isoeléctricos de las proteínas más abundantes de maca con pesos moleculares de 22,5 kD y 17,0 kD se distribuyen en un rango de puntos isoeléctricos de 7,1 a 8,2. Ambas proteínas presentan microheterogeneidad de carga, con tres isómeros de carga cada una. También muestra que la proteína de 15,5 kDa es de carácter ácido y tiene dos isómeros de carga. Por último, muestra que el polipéptido de 5,3 kDa también es de carácter ácido. Los mapas bidimensionales de las proteínas totales de los ecotipos morado y amarillo fueron similares.
DISCUSIÓN
En el presente estudio se ha evidenciado que los patrones electroforéticos de las proteínas solubles de los ecotipos blanco, morado y amarillo de raíz de maca son muy similares, cuando son separados mediante electroforesis denaturante (SDS-PAGE). Similares resultados han sido encontrados en los estudios realizados en otros cultivos andinos, como la oca ( 15 ), mashua ( 16 ). Con respecto a la proteína más abundante de la raíz de maca, a la que hemos designado el nombre de macatina en el presente trabajo, se ha determinado que tiene un peso molecular aproximado de 22,5 kDa en los 3 ecotipos analizados y constituye entre 38% y 51% de la proteína soluble total de maca. Las proteínas de otros cultivos andinos han sido purificadas de los órganos vegetativos de almacenamiento y caracterizadas bioquímicamente. En Oxalis tuberosa, oca, se ha purificado la ocatina, que constituye 40% a 60% de la proteína total soluble de los tubérculos de oca; tiene un peso molecular aparente de 18 kDa y se ha demostrado que inhibe en crecimiento de varias bacterias fitopatogénicas y de hongos ( 15 ). Se ha sido purificado de los tubérculos Solanun tuberosum, papa, la patatina, con un peso molecular aparente de 40 kDa y que constituye el 40% de la proteína total soluble ( 17 ). Varias comunicaciones indican que la patatina es una glicoproteina que tiene actividad enzimática ( 18 ), propiedades antifúngicas ( 19 ) y antioxidantes ( 20 ). Por último, en las semillas de maca se ha demostrado péptidos con actividad antifúngica ( 21 ). Aún no se ha hallado si la macatina posee alguna de las propiedades anteriormente mencionadas, que es común a las proteínas más abundantes de los órganos vegetativos de almacenamiento de otros cultivos andinos.
Otra característica a destacar en el mapa electroforético bidimensional es la existencia de 3 isoformas, tanto en la macatina como en la proteína de 17,0 kDa, que presentan puntos isoeléctricos muy parecidos. Esta característica podría deberse a la existencia de una familia de genes relacionados de macatina ( 22 ), a un mecanismo de procesamiento alternativo a nivel pre-mRNA de la macatina ( 23 ) o por una modificación post-traduccional, que implicaría la pérdida de un fragmento peptídico de la macatina recientemente sintetizada ( 24 ). Esta característica es inusual y no se ha encontrado en los mapas electroforéticos bidimensionales de otros cultivos andinos, como Solanum tuberosum papa ( 16 ) y Oxalis tuberosa oca ( 15 ).
Desde el punto de vista nutricional, distintos trabajos han determinado que la maca muestra un perfil excelente de aminoácidos ( 25 ). Responsables de este excelente balance de aminoácidos esenciales son las proteínas que están presentes en la raíz de maca. Por consiguiente, es muy factible que la proteína macatina de 22,5 kDa y la proteína de 17,0 kDa, identificadas y parcialmente caracterizadas en el presente trabajo y que constituyen el 70% de la proteína total de maca, sean ricas en aminoácidos esenciales. Los genes que codifican para estas proteínas podrían ser excelentes candidatos para ser utilizados biotecnológicamente en el mejoramiento de la calidad nutricional de otras especies comestibles con contenido bajo de aminoácidos esenciales. Esto permitirá redescubrir el enorme potencial de la maca y su contribución a la biotecnología.
REFERENCIAS BIBLIOGRÁFICAS
1. King S, Gershoff S. Nutritional evaluation of three underexploited Andean tubers: Oxalis tuberosa (Oxalidaceae), Ullucus tuberosus (Basellaceae), and Tropaeolum tuberosum (Tropaeolaceae). Economic Botany. 1987;4:503-11. [ Links ]
2. Tapia ME. Avances en las investigaciones sobre tubérculos alimenticios de los Andes. En: Agricultura Andina. Lima: Proyecto PISCA IIICA/CIID; 1984. p. 35-49. [ Links ]
3. Flores H, Flores T. Biochemistry of plant storage organs. En: JT Romeo, ed. Recent Advances in Phytochemistry. Functionality of Food Phytochemicals. New York: Plenum Publishing Corporation; 1997. p. 113-32. [ Links ]
4. Ayala G, Vílchez W. Las repercusiones de los periodos agrícolas y los pisos agroecológicos del ande Peruano sobre el consumo de alimentos y estado nutricional de la población. Lima: Editorial Empresa Científica Mundial; 1996. [ Links ]
5. Junge I. Lupino y quinua, estado actual de los conocimientos y de las investigaciones sobre su empleo en alimentación humana. Concepción: Universidad de Concepción, Chile; 1975. [ Links ]
6. Baquerizo VG. Estudio Químico-Bromatológico del Lepidium meyenii Walp. (Maca) y del Aiphanes var. Deltoidea Burret (Shica - Shica). Tesis de Bachiller en Medicina. Facultad de Medicina, Universidad Nacional Mayor de San Marcos. Lima, Perú. 1968. p. 35. [ Links ]
7. Chacón RG. Estudio fitoquímico de Lepidium meyenii Walp. Tesis de Bachiller en Ciencias Biológicas, Facultad de Ciencias de la Universidad Nacional Mayor de San Marcos. Lima, Perú. 1961. p. 46. [ Links ]
8. Torres VR. Estudio Nutricional de la Maca (Lepidium meyenii Walp.) y su aplicación en la elaboración de una bebida base. Tesis para optar el grado de Ingeniero. Facultad de Ingeniería en Industrias Alimentarias, Universidad Nacional Agraria La Molina. Lima, Perú. 1984. p. 95. [ Links ]
9. Monteghirfo GM. Determinación de aminoácidos de proteínas de quinua y kañiwa por HPLC. Theorema. 1994;5. [ Links ]
10. Yllesca GM. Estudio químico fitoquímico comparativo de 3 ecotipos de Lepidium meyenii Walp Maca procedente de Carhuamayo (Junín). Lima: Facultad de Farmacia y Bioquímica, Universidad Nacional Mayor de San Marcos; 1994. p. 103. [ Links ]
11. Beltrán SH, Baldeón MS, Carrillo FE, Fuertes RC, Arroyo AJ, Sandoval MS y Obregón VL. Estudio Botánico y Químico de los ecotipos amarillo y morado de Lepidium Peruvianum: Maca. Evaluación de su toxicidad aguda. Lima: Universidad Nacional Mayor de San Marcos / Instituto de Fitoterapia Americano; 1997. p. 9. [ Links ]
12. Flores HE, Walker TS, Guimarães RL, Bais HP, Vivanco JM. Andean root and tuber crops: Underground rainbows. Horticultural Science. 2003;38(2):161-7. [ Links ]
13. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;257:680-5. [ Links ]
14. O´Farrell P. High resolution two dimensional electrophoresis of basic as well acidic proteins. Cell. 1977;12:1133-42. [ Links ]
15. Flores T, Alape GA, Flores DM, Flores HE. Ocatin. A novel tuber storage protein from the andean tuber crop oca with antibacterial and antifungal activities. Plant Physiology. 2002;128:1291-302. [ Links ]
16. Espinoza E, Monteghirfo GM, Alvarez PJ, Arnao SA. Análisis electroforético unidimensional y bidimensional de las proteínas de Tropaeolum tuberosum (MASHUA). Boletín de la Sociedad Química del Perú. 2002;68(1):14-22. [ Links ]
17. Park WD, Blackwood C, Mignery GA, Hermodson MA, Lister RM. Analysis of the heterogeneity of the 40,000 molecular weight tuber glycoprotein of potatoes by immunological methods and by NH2-terminal sequence analysis. Plant Physiology. 1983;71:156-60. [ Links ]
18. Jiménez M, Escribano J, Gandia-Herrero F, Chazarra S, Cabanes J, García-Carmona F, Pérez-Gilabert M. Characterization of patatin esterase activity in AOT-isooctane reverse micelles. Biotechnology Progress. 2002;18(3):635-40. [ Links ]
19. Sharma N, Gruszewski HA, Park SW, Holm DG, Vivanco JM. Purification of an isoform of patatin with antimicrobial activity against Phytophthora infestans. Plant Physiology Biochemistry. 2004;43(7-8):647-55. [ Links ]
20. Liu YW. Patatin, the tuber storage protein of potato (Solanum tuberosum L.), exhibits antioxidant activity in vitro. Journal of Agricultural and Food Chemistry. 2003;51(15):4389-93. [ Links ]
21. Pizarro AJ, Monteghirfo GM. Aislamiento de una fracción proteica de bajo peso molecular de semillas de maca (Lepidium peruvianum G. Chacón) con propiedades antifúngicas. Revista de la Sociedad Química del Perú. 2003;69(4):222-8. [ Links ]
22. Thornton JW, DeSalle R. Gene family evolution and homology: Genomics meets Phylogenetics. Annual Review of Genomics and Human Genetics. 2000; l:41-73. [ Links ]
23. Kazan K. Alternative splicing and proteome diversity in plants: the tip of the iceberg has just emerged. Trends in Plant Science. 2003;8(10):468-71. [ Links ]
24. Huber SC, Hardin SC. Numerous post-translational modifications provide opportunities for the intricate regulation of metabolic enzymes at multiple levels. Plant Biology. 2004;7:318-22. [ Links ]
25. Dini A, Migliuolo G, Rastelli L, Saturnino P, Schettino O. Chemical composition of Lepidium meyenii. Food Chemistry. 1994;49:347-9. [ Links ]
Manuscrito recibido el 17 de octubre de 2007 y aceptado para publicación el 13 de diciembre de 2007.
Correspondencia:
Mg. Mario Monteguirfo Gomero
Centro de Investigación de Bioquímica y Nutrición
Facultad de Medicina, UNMSM.
Av. Grau 755. Lima 1, Perú.
Correo-e: mmonteghirfo@gmail.com

