










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versión impresa ISSN 1609-9117
Rev. investig. vet. Perú v.19 n.1 Lima ene./jun. 2008
ARTÍCULOS PRIMARIOS
Efecto de fracciones del plasma seminal, según su peso molecular, sobre la inducción de la ovulación en llamas (Lama glama)
Effect of plasma seminal fractions, based on molecular weight, on the induction of ovulation in llamas (Lama glama)
Martha Vásquez E.1 ; Wilfredo Huanca L.1,2 ; Teodosio Huanca M.3 ; Marcelo Ratto F.4 ; Gregg Adams4
1 Laboratorio de Reproducción Animal, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima
2 E-mail: whuanca2002@yahoo.com
3 Instituto Nacional de Investigación y Extensión Agraria, Puno
4 Departamento de Ciencias Biomédicas Veterinarias, Universidad de Saskatchewan, Saskatoon, Canadá
RESUMEN
El presente estudio fue realizado con el propósito de evaluar el efecto de varias fracciones de plasma seminal de llamas, según su peso molecular (PM), sobre la inducción de la ovulación. Se utilizó 54 llamas hembras sin cría y en óptimo estado reproductivo, con folículo dominante ≥7 mm de diámetro determinado por ecografía transrectal por tres días consecutivos. Los animales se distribuyeron al azar a uno de los seis tratamientos: A) fracción de plasma seminal con PM ≥30 kDa, B) fracción de plasma seminal con PM entre 10 y 30 kDa, C) fracción de plasma seminal con PM entre 5 y 10 kDa, D) fracción de plasma seminal con PM <5 kDa, E) plasma seminal completo, y F) PBS. Los animales fueron tratados con una solución de 1.5 ml de la fracción de plasma seminal o placebo correspondiente por vía intramuscular. Las llamas fueron evaluadas por ecografía en el día 2 y 9 post-tratamiento para determinar la tasa de ovulación y el tamaño del cuerpo lúteo. Los resultados indicaron una tasa de ovulación del 100% en los grupos A y E, 11.1% en B y 0% en los demás tratamientos. No se encontró diferencias estadísticas entre grupos respecto al diámetro luteal. Los resultados obtenidos permiten señalar que la fracción de plasma seminal con peso molecular mayor a 30 kDa induce la ovulación en llamas.
Palabras clave: llama, plasma seminal, ovulación inducida, factor inductor de ovulación
ABSTRACT
The present study was conducted to evaluate the effect of different fractions of llama seminal plasma, based on the molecular weight (MW), on the ovulation. Fifty four female adult llamas without calf at foot and in good reproductive conditions were used. The animals were bearing a ≥7 mm dominant follicle determined by ultrasound diagnosis during a three consecutive days. Females were randomly distributed in six groups: A, fraction of seminal plasma with MW ≥30 kDa; B, fraction of seminal plasma with MW ≥10 and <30 kDa; C, fraction of seminal plasma with MW ≥5 and <10 kDa ; D, fraction of seminal plasma with MW <5 kDa; E, complete plasma seminal; and F, PBS. Animals were treated with a 1.5 ml solution of the corresponding fraction or placebo via i.m. Ovarian ultrasound was performed at day 2 and 9 post-treatment to determine the ovulation rate and the size of the corpora lutea. Ovulation rate was 100% in groups A and E, 11.1% in group B and 0% in the other groups. No statistical differences were observed between groups on the size of the corpus luteum. The results indicated that the plasma seminal fraction with molecular weight ≥30 kDa induce ovulation in llamas.
Key words: llama, seminal plasma, induced ovulation, induce factor of ovulation
INTRODUCCIÓN
Los camélidos sudamericanos pertenecen al grupo de las especies de ovulación inducida, es decir, requieren de un estímulo para la liberación del óvulo (Sumar, 1993). Estudios realizados en estas especies, señalan que el estímulo requerido es la cópula (San Martín et al., 1968), la cual provoca la liberación de la hormona LH, y la consiguiente ovulación (Bravo et al., 1990).
Experimentos realizados en camellos bactrianos señalan que la aplicación de plasma seminal indujo la ovulación en el 87% de hembras (Chen et al., 1985), lo cual sugirió la presencia de un factor inductor de ovulación en el plasma seminal de esta especie, que posteriormente fue aislado y purificado, determinándosele un peso molecular de 19.4 kDa (Pan et al., 2001).
Un estudio realizado en alpacas con la aplicación de plasma seminal de machos de la misma especie, vía intravaginal, reportó una tasa de ovulación de 7.6% (Ríos, 1989); sin embargo, un estudio posterior reportó el 100% de ovulación utilizando plasma seminal de llama y alpaca, vía intramuscular (López, 2004). El plasma seminal contiene diferentes fracciones y su efecto sobre la inducción de la ovulación en camélidos sudamericanos aún no ha sido bien estudiado; de allí que el presente estudio tuvo como objetivo evaluar el efecto del plasma seminal, separado en base a pesos moleculares, sobre la inducción de la ovulación en llamas.
MATERIALES Y MÉTODOS
El experimento se realizó en el Anexo Quimsachata de la Estación Experimental ILLPA-INIA, ubicado a 4200 msnm, distrito de Santa Lucía, provincia de Lampa, departamento de Puno, durante los meses de febrero y marzo del año 2004.
Se seleccionaron 54 llamas hembras sin cría al pie, con historial reproductivo de haber tenido al menos un parto anterior. Todas las hembras seleccionadas recibieron las mismas condiciones de manejo (ausencia de macho) y fueron alimentadas con pastura natural. Así mismo, se colectó semen a cinco llamas, utilizando una vagina artificial adaptada de ovino, envuelta en una frazadilla eléctrica y colocada dentro de un maniquí. El semen colectado fue centrifugado a 3000 rpm durante 20 minutos para separar la fracción celular del plasma seminal.
Se hizo una primera separación de componentes no deseables (posibles residuos contaminantes) del plasma seminal utilizando filtros de 0.45 y 0.22 µ de diámetro. Los 15 ml de plasma seminal clarificado se colocaron en el primer tubo que tenía un filtro con capacidad de retener moléculas mayores a 30 kDa. Se centrifugó a 4000 g durante 20 minutos. Se obtuvo 150 µl de muestra retenida en el filtro, la cual fue resuspendida con PBS hasta completar 15 ml y constituyó la primera fracción (≥30 kDa). El plasma seminal que pasó este primer filtro contenía moléculas menores a 30 kDa y sirvió para obtener las siguientes fracciones, utilizando tubos con filtros que retenían moléculas ≥10 kDa, ≥5 kDa. La última fracción la constituyó el plasma seminal que pasó el último filtro (<5 kDa), debido a que no existen filtros con capacidad de retener moléculas menores. A cada una de las fracciones se les adicionó 50 µg ml de kanamicina, y se almacenaron a –70 ºC hasta su utilización.
En la fase pre-experimental, las hembras fueron evaluadas en diferentes oportunidades por tres días consecutivos mediante ecografía para realizar el seguimiento de la actividad ovárica y el desarrollo de las ondas foliculares, a fin de identificar aquellas que presentaban folículos en fase de crecimiento y con un tamaño ≥7 mm. Las hembras fueron distribuidas aleatoriamente en los grupos experimentales, recibiendo una dosis de 1.5 ml por vía i.m. de las siguientes fracciones de plasma seminal o placebo:
-
Fracción ≥30 kDa
-
Fracción ≥10 y <30 kDa
-
Fracción ≥5 y <10 kDa
-
Fracción <5 kDa
-
Plasma seminal completo
-
PBS
Se realizó una evaluación ecográfica de los ovarios en el día del tratamiento (Do), registrándose el tamaño del folículo dominante, otra evaluación en el día 2 (D2) para determinar la tasa de ovulación y una tercera en el día 9 (D9) para determinar la presencia y tamaño del cuerpo lúteo.
Se utilizó la diferencia de proporciones para evaluar la tasa de ovulaciones, el análisis de varianza para determinar diferencias en el tamaño del folículo dominante y del cuerpo lúteo, y la prueba de "t" Student para determinar diferencias entre promedios.
RESULTADOS Y DISCUSIÓN
Los resultados del presente estudio se encuentran en el Cuadro 1. La tasa de ovulación fue del 100% en los grupos A y E, que incluyen a los animales con la fracción de plasma seminal mayor a 30 kDa y con plasma seminal completo. Estos resultados concuerdan con otros estudios que reportaron altas tasas de ovulación con la aplicación de plasma seminal en camellos y llamas (Pan et al., 2001; López, 2004). Los resultados, no obstante, difieren de aquellos encontrados por Ríos (1989), quien obtuvo un bajo porcentaje de ovulación (8%) al aplicar plasma seminal por vía vaginal en alpacas. Es posible que la cantidad de plasma seminal tenga una relación directa con la vía de aplicación para que ocurra la ovulación en los camélidos sudamericanos.
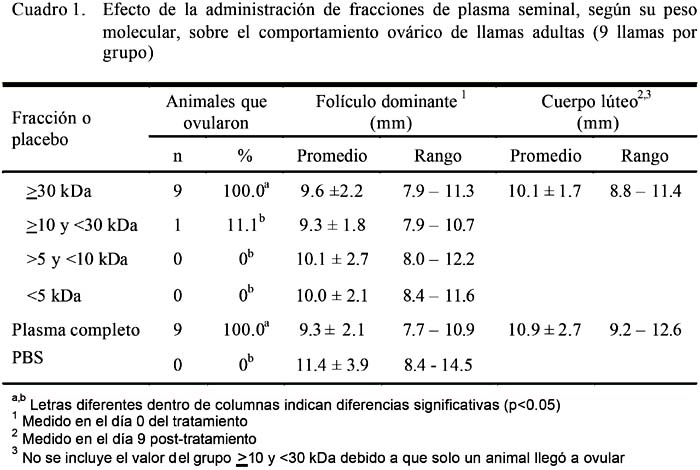
Solo llegó a ovular uno de los nueve animales que recibieron la fracción de plasma seminal con peso molecular entre ≥10 y <30 kDa. Esta ovulación podría haberse debido a un caso de ovulación espontánea y no al afecto del plasma seminal. La ovulación espontánea ocurre con una frecuencia de 5 a 15% en llamas, probablemente debido a estímulos visuales, auditivos u olfatorios (Novoa y Leyva, 1996). No obstante, Pan et al. (2001) reportaron altas tasas de ovulación al aplicar una fracción de plasma seminal con un peso molecular de 19.4 kDa en camellos bactrianos. Estas diferencias podrían explicarse por la variación entre especies, lo cual podría establecer características propias para cada especie sobre el plasma seminal, a pesar que ambas especies son filogenéticamente cercanas.
El tamaño mínimo del folículo dominante utilizado como criterio de inclusión en este estudio fue de 7 mm. Este criterio se sustenta en el reporte de Bravo et al. (1990), quién indicó que la ovulación no ocurre en hembras servidas con folículos menores de 7 mm, debido a que no llegan a inducir una suficiente secreción de LH (Bravo, 2002). Todos los folículos dominantes en el momento del tratamiento se encontraban con un tamaño ≥7 mm (Cuadro 1).
No se encontraron diferencias significativas en el tamaño del cuerpo lúteo en el día nueve post-tratamiento (día siete de la ovulación) entre los grupos A y E. Es en este momento donde el cuerpo lúteo alcanza su mayor desarrollo (Fernández-Baca et al., 1970) en la alpaca. Este resultado confirma que el factor inductor de ovulación presente en el plasma seminal de la llama es capaz de contribuir en la formación de un cuerpo lúteo de características similares al que se forma por efecto de la cópula.
Si bien es cierto que el factor inductor de ovulación parece tener similar efecto que la GnRH, no puede afirmarse que las características bioquímicas sean similares a esta hormona (Paolicchi, 1996; Pan et al., 2001), ya que en el presente estudio se obtuvo el 100% de ovulaciones con la fracción de plasma seminal con un peso molecular ≥30 kDa, peso que es significativamente mayor al de GnRH (1.2 kDa) (Mori, 2002). Por consiguiente, se podría descartar la similitud bioquímica entre ambos compuestos. Esto es, además, apoyado por el estudio de Paolicchi (1996), en el cual la exposición de células gonadotropas de la hipófisis de ratas con plasma seminal de alpaca más un anti-GnRH no modificó la secreción de LH. Tampoco podría ser de origen esteroideo, ya que los pesos moleculares de estos compuestos son inferiores a 1 kDa (Mori, 2002). El factor inductor de la ovulación en el plasma seminal tiene un peso molecular similar a LH (29 kDa), que es una glicoproteína (Mori, 2002).
El mecanismo de acción de este factor que induce la ovulación aún es desconocido, pero se puede proponer que su actividad biológica lo podría estar ejerciendo en forma directa a nivel del ovario o a nivel de tejido encefálico. En el primer caso estaría emulando la acción de la LH.
Un segundo mecanismo de acción podría ser a nivel de neuronas noradrenérgicas del cerebro medio, similar a como sucede normalmente en ovuladores inducidos después del estimulo de la cópula (Bakker y Baum; 2000), o como en el caso de los ovuladores espontáneos, donde es el estrógeno el que promueve la liberación de la noradrenalina hacia el área preóptica y eminencia media; lugares donde se localizan los cuerpos celulares y terminales de las neuronas productoras de GnRH (Demling et al., 1985; Mohankumar et al., 1994). Luego de ser estimuladas las neuronas que sintetizan GnRH, sería liberada al hipotálamo mediobasal, y desde aquí conducida a la hipófisis para inducir la secreción de LH y la posterior ovulación (Bakker y Baum; 2000). A este nivel, entonces, el factor inductor de ovulación también podría estar estimulando la secreción de GnRH como lo hacen los neurotransmisores norepinefrina, glutamato, neuropéptido y óxido nítrico, entre otros. Estos neurotransmisores, al parecer, actúan mediante un mecanismo esteroide dependiente, debido a que estudios anteriores, han demostrado que las neuronas de GnRH no poseen receptores para esteroides (Bhat et al., 1998); por tanto, estos últimos no podrían estimular la síntesis de GnRH directamente, sin la participación de los neurotransmisores. Es por esta razón que el plasma seminal podría estar comportándose como un neurotrans-misor a nivel encefálico.
En base a que se ha aislado y purificado el factor inductor de ovulación del plasma seminal de camellos bactrianos, observándose que está compuesto por una secuencia de péptidos y aminoácidos (Pan et al., 2001), y con los resultados obtenidos con el tratamiento A del presente estudio, se podría suponer que el factor inductor de ovulación podría ser de naturaleza proteica, pero esto queda por dilucidar.
CONCLUSIONES
-
El plasma seminal de llamas contiene un factor inductor de ovulación de un peso molecular mayor de 30 kDa.
-
No se registraron diferencias en el tamaño del cuerpo lúteo en los tratamientos con plasma seminal completo y plasma seminal con la fracción mayor a 30 kDa.
LITERATURA CITADA
1. Bakker Y, Baum M. 2000. Neuro-endocrine regulation of GnRH, release in induced ovulators. Neuroendocri-nology 21: 220-262.
2. Bhat K, Mahesh B, Ping L, Chorich L, Wiedmeier V, Brann D. 1998. Opioid-glutamate-nitric oxide connection in the regulation of luteinizing hormone secretion in the rat. Endocrinology 139: 955-960.
3. Bravo W, Fowler M, Stabenfeldt G, Lasley B. 1990. Ovarian follicular dynamics in the llama. Biol Reprod 43: 579-585.
4. Bravo W, Stabenfeldt G, Lasley B. Fowler M. 1991. The effect of ovarian follicle size on pituitary and ovarian responses to copulation in domesticated South American Camelids. Biol Reprod 45: 553-559.
5. Bravo W. 2002. Female reproduction. In: Bravo W (ed). The reproductive process of South American Camelids. Salt Lake City, USA: Seagull Printing. p 1-31.
6. Chen B, Yuen Z, Pan G. 1985. Semen-induced ovulation in the Bactrian camel (Camelus bactrianus). J Reprod Fert 73: 335-339.
7. Demling J, Fuchs E, Baumert M, Wuttke W. 1985. Preoptic catecholamine, GABA, and glutamate release in ovariectomised and ovariectomised estrogen-primed rats utilizing a push-pull cannula technique. Neuroendocrinology 41: 212-218.
8. Fernández-Baca S, Madden D, Novoa C. 1970. Effect of different mating stimuli on induction of ovulation in the alpaca. J Reprod Fertil 22: 261-267.
9. López A. 2004. Inducción de la ovulación en llamas mediante la administración intramuscular del plasma seminal proveniente de llama, alpaca y bovino. Tesis de Médico Veterinario. Lima: Facultad de Medicina Veterinaria, Univ. Nacional Mayor de San Marcos. 41 p.
10. Mohankumar P, Thyagarajan S, Quadri S. 1994. Correlations of catecholamine release in the medial preoptic area with proestrous surges of luteinizing hormone and prolactin: effects of aging. Endocrinology 135: 119-126.
11. Mori J. 2002. Reproductive physiology of dairy cattle. In: Manual for diagnosis and treatment of reproductive disorders in dairy cattle. Japan: Japan Livestock Technology Association. p 1-49.
12. Novoa C, Leyva V. 1996. Reproducción en alpacas y llamas. Publicación científica IVITA N° 26. 30 p.
13. Pan G, Chen Z, Liux L, Li D, Xie Q, Ling F, Fang L. 2001. Isolation an purification of the ovulation – inducing factor from seminal plasma in the bactrian camel (Camelus bactrianus). Theriogenology 55: 1863-1879.
14. Paolicchi F, Urquieta B, Del Valle L, Bustos O. 1996. Actividad biológica del plasma seminal de alpaca: Estímulo para la producción de LH por células gonadotrofas. Rev Arg Prod Anim 16: 351-356.
15. Ríos M. 1989. Presencia de un factor de inducción de la ovulación en el semen de alpacas y toro. Tesis de Médico Veterinario. Lima: Facultad de Medicina Veterinaria, Univ. Nacional Mayor de San Marcos. 30 p.
16. San Martín M, Copaira M, Zúñiga J. 1968. Aspects of reproduction in the alpaca. J Reprod Fert 16: 395-399.
17. Sumar J. 1993. Efectos de los estímulos de inducción en la ovulación de alpacas y llamas. Rev Inv Pec, IVITA 6(1): 17-21.
