










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versión impresa ISSN 1609-9117
Rev. investig. vet. Perú v.22 n.3 Lima jul./sep. 2011
Evaluación de dos métodos de criopreservación de embriones de llamas sobre las tasas de supervivencia in vivo e in vitro
Evaluation of two embryo cryopreservation methods in llama on the in vivo e in vitro embryonic survival rates
Martha Vásquez E.1, Sergio Cueva M.1, Aida Cordero R.2, Mario Lino Gonzales C.3, Wilfredo Huanca L.4,5
1 Laboratorio de Fisiología Animal, 4 Laboratorio de Reproducción Animal, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima
2 Departamento de Nutrición, Universidad Nacional Agraria La Molina, Lima
3 Instituto Nacional de Innovación Agraria, Puno
5 E-mail: whuanca2002@yahoo.com
RESUMEN
El objetivo del estudio fue evaluar el efecto de dos métodos de criopreservación sobre la supervivencia in vivo e in vitro de embriones de llama. Se recuperaron 73 embriones en estadio de blastocisto eclosionado mediante una técnica no quirúrgica a los 6.5 días post servicio en llamas superestimuladas. Las llamas receptoras se distribuyeron aleatoriamente en grupo Control (n = 14), de vitrificación (n=30) y de congelación lenta (n=29). Para la vitrificación, los embriones fueron expuestos a la solución de vitrificación (SV) conteniendo 20% Glicerol + 20% Etilenglicol + 0.5M Sucrosa + 10% suero fetal bovino (SFB) + 50 μg/ml sulfato de gentamicina, y sumergidos en nitrógeno líquido dentro de pajillas de 0.25 ml. Para la congelación lenta, los embriones fueron expuestos a fosfato buffer salino (PBS) con 1.5 M de Etilenglicol + 10% de SFB + 50 μg/ml de sulfato de gentamicina, aspirados en pajillas de 0.25 ml, y enfriados a una tasa de descenso de 0.12 °C/min hasta 5 °C y, luego, en la boca del tanque de nitrógeno, se continuó el descenso a una tasa de 5 °C /min hasta -20 °C por 5 min, y luego se sumergieron en el nitrógeno líquido. En la descongelación se utilizaron soluciones de dilución conteniendo dos concentraciones de sucrosa: 0.5 M y 0.2 M para congelación lenta y 0.25 M y 0.12 M para vitrificación. Se realizó una evaluación in vivo a todos los embriones del grupo control y al 50% de los embriones de los dos grupos experimentales, mediante transferencia directa a hembras receptoras previamente sincronizadas. El diagnóstico de preñez se llevó a cabo por ecografía transrectal a los 20 y 30 días. La preñez fue de 4/13, 2/12 y 0/11 para las receptoras de los grupos Control, Vitrificados y Congelación lenta, respectivamente, sin diferencia estadística. Para la evaluación in vitro, los embriones criopreservados fueron cultivados en PBS + 20% SFB, en una atmósfera compuesta por 5% de CO2, 20% de O2 y 75% de N2, durante 1 h a 39 °C, y se observó su re-expansión mediante la observación de su morfología. Se obtuvo 75.0% ƙ/12) de re-expansión en embriones vitrificados y 57.1% Ɣ/7) en embriones congelados lentamente Ɣ/7), sin diferencia significativa. Los resultados permiten concluir que la vitrificación podría ser el método adecuado para la criopreservación de embriones de llama.
Palabras clave: llama, vitrificación, congelación lenta, preñez, reexpansión
ABSTRACT
The aim of the study was to evaluate in llama embryos the effect of two cryopreservation methods on the in vivo and in vitro survival rate. Seventy three hatched blastocysts were recovered by a non-surgical technique at day 6.5 after mating from superstimulated llamas. Receptors were randomly allocated to a control group (n=14), vitrification (n=30) and slow freezing (n=29). On vitrification, embryos were exposed to a vitrification solution (VS) containing 20% Glycerol + 20% Ethylene glycol + 0.5 M Sucrose + 10% fetal calf serum (FCS) + 50 μg/ml gentamicin sulfate, and then plunged into liquid nitrogen in 0.25 ml straws. On the slow freezing, embryos were exposed to phosphate buffer saline (PBS) with 1.5 M Ethylene glycol + 10% FCS + 50 μg/ml gentamicin sulfate, loaded in 0.25 ml straws, and cooled at a rate of 0.12 °C/min to 5 °C. Then, further temperature decrease at 5 °C /min rate, to -20 °C, for 5 min at the mouth of the nitrogen tank; finally straws were plunged into liquid nitrogen. For thawing, two dilution solutions were used composed of two sucrose concentrations: 0.5 M and 0.2 M for slow freezing, and 0.25 M and 0.12 M for vitrification. An in vivo evaluation was performed in all embryos of the control group and in 50% of the experimental groups by direct transfer into previously synchronized female recipients. Pregnancy diagnosis was carried out by transrectal ultrasound evaluation at 20 and 30 days. Pregnancy was in 4/13, 2/12, and 0/ 11 in recipients from control, vitrification and slow freezing groups respectively, without significant difference. For the in vitro evaluation cryopreservated embryos were cultured in PBS + 20% FCS under atmosphere compose of 5% CO2, 20% O2, and 75% N2 at 39 °C for 1 h, then reexpansion was recorded by morphological characteristics. Embryo reexpansion was 75% ƙ/12) in vitrified embryos and 57.1% Ɣ/7) in slow freezing embryos, and without significant difference. It was concluded that vitrification could be a suitable method for llama embryo cryopreservation.
Key words: llama, vitrification, slow freezing, pregnancy, reexpansion
INTRODUCCIÓN
Los avances en la criobiología, la biología celular y la embriología de animales domésticos han hecho capaz el desarrollo de metodologías de preservación de embriones de varias especies domésticas (Dobrinsky, 2002) y, recientemente, en llamas (Palasz et al., 2000; Aller et al., 2002). La criopreservación, junto a la transferencia de embriones e inseminación artificial, es una de las técnicas que puede contribuir al incremento de la productividad animal, ya que permite la preservación y transporte de germoplasma (Dobrinsky, 2002) y, por lo tanto, se incrementa la posibilidad de incrementar el número de crías por hembra a lo largo de su vida reproductiva.
En la criopreservación de embriones se tiene el método convencional, que consiste en una congelación lenta, y el método de vitrificación, que consiste en un enfriamiento muy rápido. Existen reportes de embriones de dromedarios (Camelus dromedarius) crioservados con el método convencional con resultados de preñez relativamente bajos Ƙ%, Nowshari et al., 2005; 32% Skidmore et al., 2004). Kaidi et al. ) compararon el efecto de ambos métodos sobre la morfología y metabolismo del blastocisto bovino producido in vitro, observando disminución en el consumo de glucosa, piruvato y oxígeno en la congelación lenta, además de un incremento en la actividad glucolítica, indicando un efecto de estrés.
La vitrificación permite la conservación de tejidos y células sin formación de cristales de hielo, y ha mostrado ser un método apropiado para la criopreservación de embriones, lográndose obtener crías vivas de varias especies domésticas después de transferir embriones criopreservados por esta técnica; por ejemplo 50% de preñez en ovejas (Papadopoulos et al., 2002).
Se han realizado pocos estudios sobre los efectos de la criopreservación en la viabilidad embrionaria en llamas (Palasz et al., 2000; Aller et al., 2002; Lattanzi et al., 2002), de allí que el objetivo del estudio fue evaluar la supervivencia embrionaria in vivo e in vitro de embriones de llama, criopreservados mediante los métodos de vitrificación y congelación lenta.
MATERIALES Y MÉTODOS
Lugar de Estudio
La fase de colección y evaluación de embriones in vivo se realizó en el C.I.P. Quimsachata, del Instituto Nacional de Innovación Agraria (INIA), ubicado en el departamento de Puno, durante los meses de febrero y marzo de 2008. La evaluación in vitro se realizó en la sección de Biotecnología Reproductiva del Laboratorio de Reproducción Animal de la Facultad de Medicina Ve - terinaria, Universidad Nacional Mayor de San Marcos, Lima.
Animales y Embriones
Se utilizaron 59 llamas hembras adultas ྷ donadoras y 36 receptoras) de 3 a 6 años de edad, con pesos promedio de 115 kg. Los animales presentaron un excelente historial reproductivo y buenas condiciones fisiológicas. Todas recibieron el mismo manejo (ausencia de macho) y fueron alimentadas sin restricciones sobre pasturas naturales.
El criterio de selección para las hembras donadoras fue la presencia de un folículo >7 mm determinado mediante ultrasonografía transrectal con un ecógrafo ALOKA SSD 500 y transductor lineal Modo B de 7.5 MHz. Estos animales fueron sometidos a un protocolo de estimulación ovárica, descrito por Huanca ), y servidos dos veces, a intervalos de 12 horas, con machos de fertilidad comprobada. Se aplicó una dosis de 0.4 mg de acetato de buserelina vía i.m. para asegurar la ovulación.
Los embriones fueron recuperados por un método no quirúrgico a los 6.5 días de la monta (Huanca, 2005). Se colectaron 73 embriones en estadio de blastocisto. La calidad de los embriones fue de excelente o buena (Grado 1) y regular (Grado 2), según el criterio de clasificación de la International Embryo Transfer Society (IETS, 1998), adaptado para el estudio. Asimismo, se realizó una medición del diámetro de los embriones, previo al procedimiento experimental de criopreservación, empleando una escala micrométrica en el objetivo del estereoscopio.
Diseño Experimental
Los embriones fueron aleatoriamente distribuidos en tres grupos experimentales, bajo un diseño completamente randomizado: G1, Grupo control: embriones frescos (n=14) G2, embriones vitrificados (n=30) y G3, embriones congelados lentamente (n=29). Para la evaluación in vivo (transferencia mbrionaria), los embriones fueron transferidos a las receptoras de la siguiente manera: G1, 14 embriones frescos en 13 receptoras (n=13) G2, 16 embriones vitrificados en 12 receptoras (n=12) G3, 11 embriones congelados lentamente en 11 receptoras (n=11). Los embriones restantes (G2, n=12; G3, n=7) se utilizaron en la evaluación in vitro. Para efectos del estudio, solo se utilizaron los embriones con un diámetro menor a 280 m.
Criopreservación
En el método de vitrificación se utilizó el protocolo reportado por Von Baer et al. ) con modificaciones. Se utilizó dos soluciones, una solución de equilibrio ƕ% glicerol + 5% etilenglicol + 0.2 M sucrosa + 10% suero fetal bovino + 50 m/ml gentamicina) a 25 ºC, donde el embrión permanece por 5 min, y una solución de vitrificación ྴ% glicerol + 20% etilenglicol + 0.5 M sucrosa + 10% suero fetal bovino + 50 ug/ml gentamicina) donde se mantiene al embrión por 1 min. El embrión se coloca en pajuelas de 0.25 ml Ƒ embrión por pajuela) y es sumergido de inmediato en el nitrógeno líquido.
En el método de congelación lenta se usaron los protocolos descritos por Palasz et al. ) y Lattanzi et al. ). Los embriones se colocaron en solución de congelación ྪ% etilenglicol + 10% SFB + 50 m/ml de gentamicina) a 26 ºC durante 5 min y envasados en pajuelas de 0.25 ml. El descenso de temperatura se realizó a una tasa de 0.12 °C/ min en 3 horas, luego a 5 °C hasta -20 °C, y finalmente se sumergieron las pajuelas en el nitrógeno líquido.
Descongelación de Embriones
Las pajuelas fueron extraídas del tanque de nitrógeno, mantenidas al ambiente por 10 s y colocadas a 37 °C por 1 min. De allí, los embriones vitrificados fueron vertidos en una solución de 100 μL de sucrosa 0.5 M e, inmediatamente, a otra de igual concentración por 5 min y después a una tercera solución de sucrosa 0.2 M por 5 min. Posteriormente, fueron lavados 3 veces con PBS + 20% Suero Fetal (Von Baer et al., 2002). En el caso de los embriones congelados en forma lenta, las primeras dos soluciones de sucrosa fueron 0.25 M y la última fue 0.12 M.
Cultivo de Embriones Descongelados
Para la evaluación in vivo, los embriones se cultivaron por 2 h en 100 μL de PBS + 20% de Suero Fetal a 35 °C. Para la evaluación in vitro se cultivaron por 1 h a 39 °C a 5% de CO2, 20% de O2 y 75% de N.
Evaluación in vivo
Dieciséis embriones vitrificados y 11 de congelación lenta fueron transferidos a 12 y 11 receptoras, respectivamente; mientras que 14 embriones frescos (control) fueron transferidos a 13 receptoras. Para la transferencia, los embriones se colocaron en pajuelas de 0.25 ml y se depositaron en el cuerno uterino ipsilateral al cuerpo lúteo a los 6.5 días post-inducción de la ovulación. El diagnóstico de gestación se realizó por ecografía a los 14 y 24 días después de la transferencia, determinándose la presencia o ausencia de la vesícula embrionaria.
Evaluación in vitro
Doce embriones vitrificados y 7 de congelación lenta fueron lavados tres veces en solución de 500 μl de PBS + 20% Suero Fetal, para luego del cultivo, verificar si la morfología fue similar a la morfología previa a la criopreservación. Luego, se realizó la tinción de los embriones para determinar la viabilidad de las células embrionarias mediante las alteraciones a nivel de membrana plasmática. Se utilizó ioduro de propidio (PI, Sigma) y bisbenzimida (BIS, Hoechst 3342, Sigma). El ioduro de propidio es un marcador de ácido nucleico que ingresa solo a células con alteraciones en la integridad de membrana, mientras que el bisbenzimida ingresa a todas las células y se une al DNA celular. En células con alteraciones de membrana, la fluorescen cia del BIS es disminuida por el PI, el cual absorbe esta energía y emite fluorescencia roja.
El procedimiento de tinción consistió en incubar los embriones en PBS con 10 μg/ml PI a 39 °C por 15 min, luego fijados en 70% de etanol helado ƕ °C) por 5 min. Después, se les transfirió a etanol con 10 μg/ml BIS a temperatura ambiente por 5 min; y, finalmente, fueron colocados en una gota de glicerol en una lámina portaobjetos, cubierto con un cubreobjetos, y observados en un microscopio de fluorescencia modelo Zeiss KB-L8 (Kaidi et al., 2001). La viabilidad embrionaria se determinó mediante la observación de núcleos fluorescentes color azul.
Análisis Estadístico
Los datos fueron analizados mediante Chi-Cuadrado y la prueba de Fisher Exacta para determinar posibles asociaciones entre los tratamientos con la tasa de preñez, y entre la calidad embrionaria antes y después con la tasa de preñez durante la evaluación in vivo.
La prueba de Fischer exacta fue empleada para determinar la probable asociación entre los tratamientos con la tasa de reexpansión y asociación entre los tratamientos y la calidad embrionaria in vitro. Los análisis estadísticos fueron realizados empleando el programa estadístico STATA v. 10.0 (StataCorp, Texas USA).
RESULTADOS Y DISCUSIÓN
La frecuencia de animales gestantes, luego de la transferencia, no fue estadísticamente diferente entre los embriones frescos Ɣ/13), vitrificados ƒ/12) y congelados lentamente Ɛ/11). El diámetro embrionario fue estadísticamente similar en las vacas que recibieron los embriones frescos 鶑 ± 35 μm), vitrificados 鵢 ± 35 μm), y congelados lentamente 鵿 ± 29 μm). Asimismo, el diámetro del cuerpo lúteo de las receptoras fue de 10 ± 1 mm, sin diferencia entre grupos.
Aller et al. ), reportaron una mayor tasa de preñez con embriones vitrificados en comparación con los resultados del presente estudio; sin embargo, solo trabajaron con 4 animales. Por otro lado, en congelación lenta solamente se ha reportado 32% de preñez en dromedarios (Camelus dromedarius) (Skidmore et al., 2004). Las diferencias pueden ser atribuidas al tiempo de exposición y tasa de descenso de temperatura. La menor tasa de descenso de temperatura ocasiona una entrada de los crioprotectores permeables a la célula y una deshidratación muy lenta, lo que conlleva a la formación de hielo intracelular (Fair et al., 2001). Ello explicaría el posible daño ocasionado en los embriones de este grupo y su fracaso en la viabilidad in vivo.
El diámetro embrionario también podría estar relacionado al éxito de la criopreservación, ya que embriones más grandes, de mayor volumen y líquido intracelular, tendrían mayor probabilidad de obtener una deshidratación incompleta durante la criopreservación, con el consiguiente daño celular por formación de cristales de hielo. Se reporta una falla en la transferencia de embriones en equinos cuando los embriones fueron mayores de 300 m de diámetro (Eldridge-Panuska et al., 2005). En el presente estudio, las receptoras que mantuvieron los embriones recibieron embriones vitrificados de un diámetro de 280 m, siendo el diámetro promedio de todos los embriones del experimento de 302 ± 17 m.
Es posible que embriones de llama, en estadios más tempranos, y con menor diámetro, serían más adecuados para ser criopreservados, ya que en un estudio previo con alpacas, blastocistos liberados de 7 días, con un diámetro promedio de 450 μm, vitrificados y transferidos a receptoras no produjo ninguna preñez (Vásquez et al., 2007). Por el contrario, Skidmore et al ) reportaron preñeces utilizando 21 blastocistos de 6 días. El protocolo empleado en ese estudio fue distinto al utilizado en el presente trabajo; sin embargo, incluyó las mismas concentraciones de los crioprotectores permeables.

Otro factor determinante que no puede dejarse de lado es el estado nutricional de las receptoras. Whitaker et al. ) encontraron menor liberación de LH en respuesta a inyecciones de GnRH, y Pereyra y Linares , citado por Rutter, 2008) encontraron menores niveles de progesterona en vacas subalimentadas. Los animales experimentales fueron alimentados con pasturas naturales y este factor nutricional podría haber ocasionado una menor funcionalidad del cuerpo lúteo y, en consecuencia, mortalidad embrionaria temprana.
Se observó variación estadística de la calidad embrionaria post descongelación en ambos métodos de criopreservación, aunque esto se observó mayormente en los embriones que fueron destinados a la evaluación in vivo (Cuadro 1). Es importante hacer notar, que en la evaluación in vitro, durante el proceso de dilución del crioprotector (salida del crioprotector), se registraron las características morfológicas embrionarias normales y anormales.
La calidad del embrión parece estar influenciada por el tratamiento superestimulatorio de la donadora, debido probablemente a los cambios hormonales en los fluidos foliculares y estructurales en los oocitos, los cuales podrían afectar la fertilización y desarrollo embrionario temprano (Callensen et al., 1986). También existe una estrecha relación entre la calidad embrionaria con la futura tasa de preñez (Abe et al., 2002).
Entre los cambios morfológicos anormales que se observaron en los embriones congelados por ambos métodos está la pérdida celular, vista como una descamación de células, la cual se podría atribuir a la pérdida de las conexiones intercelulares en las células trofoblásticas, lo cual ha reportado en embriones bovinos vitrificados, y sería también, la causa del colapso del blastocele (Vajta et al., 1997). Además, se observó que algunos embriones permanecieron contraídos, debido probablemente a la destrucción irreversible de los filamentos de actina, constituyentes del citoesqueleto de todas la células eucariotas; causado posiblemente por daño osmótico, como ha sido reportado en embriones porcinos (Lang et al., 1998).
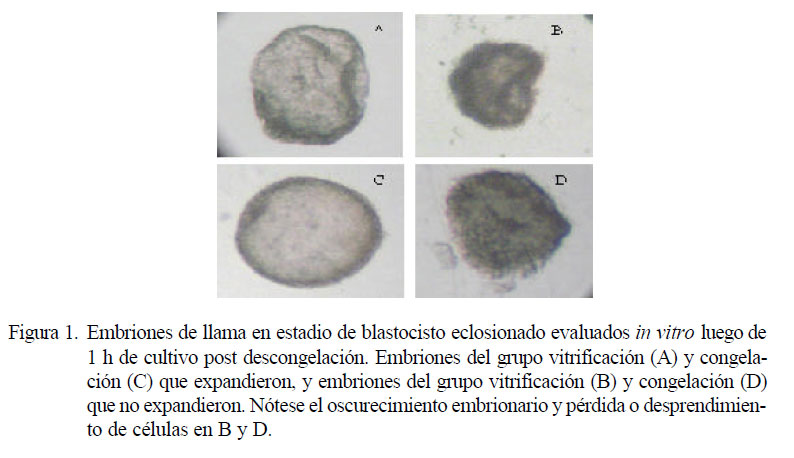
La reexpansión embrionaria es una indicación de la viabilidad embrionaria, ya que muestra la capacidad que tiene el embrión para poder eliminar el crioprotector, rehidratarse y recuperar su forma inicial previa a la criopreservación. Durante la evaluación in vitro, el 75% ƙ/12) de los embriones vitrificados se reexpandieron, en tanto que el 57.1% Ɣ/7) de los embriones sometidos a congelación lenta presentaron reexpansión, no siendo significativa esta diferencia (Fig. 1).
El porcentaje de embriones que reexpandieron en el grupo vitrificación concuerda con el estudio realizado por Men et al. ), donde un alto porcentaje de embriones vitrificados y que presentaron cambios morfológicos normales se restablecieron luego de un cultivo in vitro. Por otro lado, la tasa de reexpansión embrionaria in vitro de embriones congelados del presente estudio ࿙.1%) es menor a lo reportado en otros estudios %, Palasz et al., 2000; 63.2%, Lattanzi et al., 2002) a las 12 y 24 horas del cultivo, respectivamente. Aunque la reexpansión de los embriones del estudio no fue total, debido posiblemente al corto periodo de cultivo Ƒ h), los resultados indican que la vitrificación fue el método que produjo menos daño a las células embrionarias y permitió mayor viabilidad in vitro, en comparación a la congelación lenta.
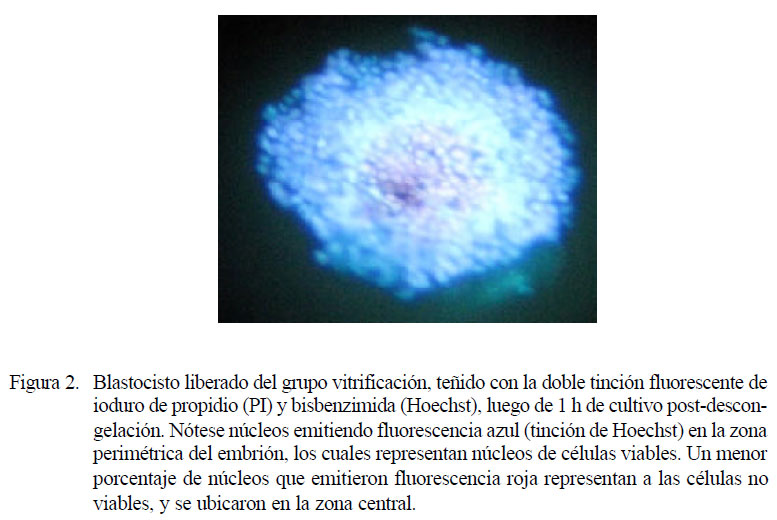
El porcentaje de células viables y no viables de tres embriones vitrificados post-descongelación fue medido a través de una tinción fluorescente (doble tinción de ioduro de propidio/ bisbenzimida, Hoechst). Se contó un promedio de 919 núcleos, de los cuales 633.7 .0%) emitió fluorescencia azul (Fig. 2). Esto permite reafirmar que el método de vitrificación dañó a un menor porcentaje de células embrionarias, lo que se traduce en un mejor pronóstico de la futura viabilidad del embrión.
Los embriones de llamas, al igual que los embriones de porcinos y bovinos, parecen ser especialmente sensibles al enfriamiento (von Baer et al., 2002), por poseer mayor cantidad de lípidos intracelulares (Nagashima et al., 1999; Pereira y Marques, 2008). Esta característica podría disminuir su tolerancia a los daños causados por la criopreservación, lo que haría que los embriones de camélidos sudamericanos necesiten de un procedimiento de criopreservación que disminuya el tiempo de exposición a temperaturas críticas, como sucede en la vitrificación.
Los resultados permiten concluir que el método de vitrificación en embriones de llama produjo mayor viabilidad in vitro que el método de congelación lenta.
LITERATURA CITADA
1. Abe H, Matsukasi S, Hoshi H. 2002. Ultraestructural differences in bovine morula classified as high and low qualities by morphological evaluation. Theriogenology 57: 1273-1283.
2. Aller J, Rebuffi G, Cancino A, Alberico R. 2002. Successful transfer of vitrified llama (Lama glama) embryos. Anim Reprod Sci 73: 121-127.
3. Callensen S, Greve T, Hytell P. 1986. Preovulatory endrocrinology and oocyte maduration in superovulated cattle. Theriogenology 25: 71-78.
4. Dobrinsky J. 2002. Advancements in cryopreservation of domestic animal embryos. Theriogenology 57: 285-602.
5. Eldridge-Panuska W, Caracciolo di Brieza V, Seider Jr G, Squires E, Carnevale E. 2005. Establishment of pregnancies after serial dilution or direct transfer by vitrified equine embryos. Theriogenology 63: 1308-1319.
6. Fair T, Lonergan P, Dinnyes A, Cotell D, Hytell P, Ward F, Boland M. 2001. Ultrastructure of bovine blastocysts following cryopresevation: Effect of method of blastocyst production. Mol Reprod Dev 58: 186-195.
7. Huanca W. 2005. Aplicación de biotecnologías reproductivas en especies domésticas y silvestres de camélidos sudamericanos. Agrociencia, Uruguay 9Ƒ- 2): 505-509.
8. International Embryo Transfer. 1998. Manual of the International ET Society. 3rd ed. Savoy IL. IETS: USA. 170 p.
9. Kaidi S, Bernard P, Lambert A, Massip A, Dessy F, Donnay I. 2001. Effect of conventional controlled-rate freezing and vitrification on morphology and metabolism of bovine blastocysts produced in vitro. Biol Reprod 65: 1127- 1134.
10. Lang F, Busch G, Ritter M, Volkl H, Waldegger S, Gulbins E. 1998. Functional significance of cell volume regulatory mechanism. Physiol Rev 78: 247-305.
11. Lattanzi C, Santos C, Chaves G, Miragaya M, Capdevielle E, Judith E, Agüero A, Baranao L. 2002. Cryopreservation of llama (Lama glama) embryos by slow freezing and vitrification. Theriogenology 57: 585 (Abstr).
12. Men H, Agca Y, Mullen S. 2005. Osmotic tolerant of in vitro produced porcine blastocysts assessed by their morphological integrity and cellular actin filament organization. Cryobiology 51: 119-129.
13. Nagashima H, Cameron R, Kuwayama M, Young M, Beebe L, BlackshawA, Nottle M. 1999. Survival of porcine delipated oocytes and embryos after cryopreservation by freezing or vitrification. J Reprod Dev 45: 167-176.
14. Nowshari M, Ali S, Saleem S. 2005. Offspring resulting from transfer of cryopreserved embryos in camel (Camelus dromedarius). Theriogenology 63: 2512-2522.
15. Palasz A, Adams G, Brogliatti G, Mapletofl R. 2000. Effect of collection and of permeating cryoprotectans on llama (Lama glama) embryos and trophoblastic vesicles. Theriogenology 53: 341 (Abstr).
16. Papadopoulos S, Rizos D, Duffy P, Wade P, Quinn K, Boland M, Lonergan P. 2002. Embryo survival and recipient pregnancy rates after transfer of fresh or vitrified, in vivo or in vitro produced ovine blastocysts. Anim Repro Sci 74: 35-44.
17. Pereira R, Marques C. 2008. Animal oocyte and embryo cryopreservation. Cell Tissue Banking 9: 267-277.
18. Rutter B. 2008. Programa de entrenamiento sobre manejo reproductivo de vacas lecheras. Red de información y capacitación agropecuaria. Lima: PERULACTEA. 204 p.
19. Skidmore J, Billah M, Loskutoff N. 2004. Developmental competence in vitro and in vivo of cryopreserved, hatched blastocysts from the dromedary camel (Camelus dromedarius). Reprod Fertil Dev 16: 605-609.
20. Vajta G, Hyttel H, Callensen D. 1997. Morphological changes of in vitro produced bovine blastocysts after vitrification, in straw direct rehydration, and culture. Mol Reprod Dev 48: 9-17.
21. Vásquez M, Cervantes M, Cordero A, Cárdenas O, Huanca T, Huanca W. 2007. Vitrificación de embriones de alpacas: estudio preliminar. En: XX Reunión de la Asociación Latinoamericana de Producción Animal. Cusco, Perú: ALPA.
22. Von Baer A, Del Campo M, Donoso X, Toro F, Von Baer L, Montecinos S, Rodríguez-Martínez, H, Palasz A. 2002. . Vitrification and cold storage of llama (Lama glama) hatched blastocysts. Theriogenology 57: 489 (Abstr).
23. Whitaker D, Smith E, Da Rosa G, Kelly M. 1993. Some effects of nutrition and management on the fertility of dairy cattle. Vet Rec 133: 61-64.
