










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versión impresa ISSN 1609-9117
Rev. investig. vet. Perú v.23 n.2 Lima abril./jun 2012
EVALUACIÓN DE LA INTEGRIDAD DE ADN MEDIANTE CITOMETRÍA DE FLUJO EN ESPERMATOZOIDES DE ALPACA CRIOPRESERVADOS CON ANÁLOGOS DE SUPERÓXIDO DISMUTASA
FLOW CYTOMETRY EVALUATION OF DNA INTEGRITY OF ALPACA SPERM AFTER CRYOPRESERVATION WITH ANALOGUES OF SUPEROXIDE DISMUTASE
Alexei Santiani A.1,5, Shirley Evangelista V.2, Carolina Cheuquemán A.3, Alejandra von Baer H.4, Jennie Risopatrón G.3, Raúl Sánchez G.3
1 Laboratorio de Reproducción Animal, Facultad de Medicina Veterinaria, 2 Laboratorio de Fisiología de la Reproducción Animal, Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Lima, Perú
3 Centro de Biotecnología de la Reproducción (CEBIOR), Facultad de Medicina, Universidad de La Frontera, Temuco, Chile
4 Criadero Llamas del Sur, Temuco, Chile
5 E-mail: asantiani@hotmail.com
6 Terminal deoxynucleotidyl transferase-mediated fluorescein–dUTP nick end labeling [TUNEL]
RESUMEN
Se evaluó la estabilidad del ADN en espermatozoides de alpaca mediante citometría de flujo en muestras congeladas con antioxidantes análogos de superóxido dismutasa (Tempo y Tempol). Doce muestras de semen de alpaca fueron congeladas utilizando un dilutor en base a leche descremada, fructosa, yema de huevo y etilenglicol. Cada muestra fue dividida en tres porciones: grupo control, grupo Tempo (1 mM) y grupo Tempol (1 mM). Los antioxidantes fueron agregados al dilutor durante la curva de enfriamiento (10 °C). Las muestras fueron descongeladas y fijadas en una solución de formaldehido al 2% y permeabilizadas utilizando una solución de Triton X-100 al 0.8%. La integridad del ADN espermático se evaluó con la técnica de TUNEL y una sonda fluorescente para determinar la viabilidad celular (Ioduro de propidio [PI]). Los espermatozoides marcados con la sonda del TUNEL y PI fueron considerados células permeabilizadas con el ADN fragmentado. Las muestras fueron analizadas mediante citometría de flujo utilizando un láser de 488 nm, contando un mínimo de 10 000 células por muestra y utilizando microscopía de fluorescencia. La frecuencia de espermatozoides con ADN fragmentado en el grupo control (38.8 ± 16.2%) fue significativamente mayor (p<0.05) al grupo Tempol (16.7 ± 8.4%), mientras que los resultados del grupo Tempo (25.4 ± 5.8%) fueron estadísticamente similares a los grupos Control y Tempol. Se concluye que la adición del análogo de superóxido dismutasa, Tempol (1 mM), durante el proceso de criopreservación de espermatozoides de alpaca puede prevenir parcialmente el daño al ADN espermático.
Palabras clave: criopreservación, espermatozoides de alpaca, superóxido dismutasa, citometría de flujo, ADN, antioxidantes
ABSTRACT
DNA integrity in alpaca spermatozoa was evaluated by flow cytometric analysis in sperm cryopreserved using antioxidants analogues of superoxide dismutase (Tempo and Tempol). Twelve alpaca semen samples were frozen using an extender based on skim milk, fructose, egg yolk, and ethylene glicol. Each sample was divided into three aliquots: Control group, Tempo group (1 mM), and Tempol group (1 mM). Antioxidants were added during cooling at 10 °C. After thawing, samples were fixed using 2% formaldehyde solution and permeabilizated using 0.8% Triton X-100 solution. TUNEL assay and Iodure propidium (PI) were used for evaluation of DNA integrity and cell permeability. Spermatozoa labeled with TUNEL and PI was classified as cells with DNA damaged. Samples were analyzed by flow cytometric using a 488 nm laser, counting at least 10 000 cells per sample, and by epifluorescene microscopy. Frequency of DNA damaged sperm in control group (38.8 ± 16.2%) was significantly higher (p<0.05) than in the Tempol group (16.7 ± 8.4%), whereas the results in the Tempo group (25.4 ± 5.8) were similar to the control and Tempol groups. It was concluded that the addition of superoxide dismutase analogue Tempol (1 mM) during cooling (10 °C) in alpaca sperm cryopreservation partially prevents sperm DNA damage.
Key words: cryopreservation, alpaca sperm, superoxide dismutase, flow cytometry, DNA, antioxidants
INTRODUCCIÓN
Los estudios sobre criopreservación de espermatozoides de alpaca han utilizado diversos protocolos de congelamiento/ descongelamiento, así como dilutores y crioprotectores; no obstante, los resultados no han sido los esperados. La motilidad post descongelamiento en espermatozoides de semen de alpaca (Santiani et al., 2005) y espermatozoides epididimarios (Morton et al., 2007, 2010; Banda et al., 2010) varía entre 15 a 20%. Del mismo modo, la viabilidad celular e integridad acrosomal post descongelamiento ha sido inferior al 30% (Santiani et al., 2005; Banda et al., 2010). Las razones para la escasa supervivencia de los espermatozoides de alpaca al proceso de criopreservación no están bien entendidas; sin embargo, algunos factores como la susceptibilidad inherente de cada especie, los componentes de los dilutores y la producción de especies reactivas de oxígeno son factores sugeridos en otras especies, donde tampoco ha sido posible obtener elevados porcentajes de supervivencia espermática (Medeiros et al., 2002).
Las especies reactivas de oxígeno (ROS) participan en una serie de eventos fisiológicos del espermatozoide, como la capacitación espermática, hiperactivación de la motilidad y reacción acrosomal (Aitken y Fisher, 1994; De Lamirande et al., 1997). Sin embargo, se ha demostrado que niveles elevados de ROS pueden causar un efecto nocivo en los espermatozoides, como disminuir la motilidad y viabilidad espermática, inducir una indeseable capacitación espermática prematura (Peris et al., 2007; Ruiz et al., 2007; Santiani, 2003), aumentar la peroxidación de ácidos grasos insaturados de la membrana espermática (Aitken et al., 1989) y afectar la integridad del ADN espermático (Aitken et al., 1998).
Se ha demostrado en espermatozoides de ovinos (Santiani, 2003; Peris et al., 2007), bovinos (Bilodeau et al., 2000) y humanos (Wang et al., 1997) que los niveles de ROS aumentan significativamente durante el proceso de criopreservación. Estos niveles elevados podrían tener un rol importante en la disminución de la función espermática durante el proceso de criopreservación (Sue-Hee et al., 2010). Uno de los cambios observados en espermatozoides criopreservados es la inducción de la apoptosis celular y daño del ADN espermático (Agarwal y Said, 2005). Incluso, existen evidencias que el daño del ADN espermático tiene una correlación positiva con mayores niveles de ROS (Baumber et al., 2003). En ese sentido, la evaluación del ADN espermático mediante TUNEL6 por citometría de flujo, podría constituir una alternativa para determinar el efecto nocivo de ROS durante la criopreservación (Cordelli et al., 2005). La citometría de flujo tiene la ventaja que incluye un proceso automatizado que evalúa más de diez mil células en forma objetiva, en comparación con la microscopía tradicional que evalúa en forma subjetiva no más de 200 células (Muratori et al., 2008).
Una alternativa para prevenir el efecto negativo de los ROS durante la criopreservación espermática puede ser la adición de antioxidantes análogos a superóxido dismutasa al medio dilutor (Santiani, 2003; Ruiz et al., 2007). No obstante, no existen trabajos publicados sobre la producción de especies reactivas de oxígeno ni sobre la utilización de antioxidantes en este evento durante el proceso de criopreservación en la alpaca; por lo tanto, el objetivo del presente trabajo fue evaluar el efecto de dos antioxidantes (Tempo y Tempol) análogos de superóxido dismutasa durante los procesos de criopreservación de espermatozoides de alpaca sobre la integridad del ADN espermático utilizando la citometría de flujo.
MATERIALES Y MÉTODOS
Lugar de Estudio
La colección de semen de alpaca y criopreservación espermática fueron realizadas en la estación experimental del Instituto Veterinario de Investigaciones Tropicales y de Altura (IVITA), ubicada en Maranganí, Cusco, a una altura de 3550 msnm. El descongelamiento y fijación de muestras se hicieron en el Laboratorio de Reproducción Animal, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima, Perú. La evaluación de la integridad del ADN espermático mediante citometría de flujo fue realizada en el Centro de Biotecnología de la Reproducción (CEBIOR), Facultad de Medicina Humana, Universidad de La Frontera, Temuco, Chile.
Animales y Semen
Se colectaron muestras de semen de nueve alpacas Huacaya, pertenecientes al IVITA-Maranganí. El entrenamiento de los machos y la colección seminal se hizo en los meses de enero y febrero, que corresponden a la temporada reproductiva de los camélidos sudamericanos. El semen fue colectado utilizando el método de vagina artificial con hembras receptiva (Dávalos y Olazábal, 2002) y las muestras fueron transportadas de inmediato al laboratorio. Todas las muestras de semen fueron sometidas a una aspiración continua a través de una jeringa de tuberculina con aguja de 21 x 1" para disminuir la viscosidad seminal
Manejo del Semen
De 30 eyaculados obtenidos con buena calidad seminal, se congelaron 12 eyaculados (provenientes de 6 machos) según el protocolo descrito por Santiani et al. (2005), que emplea el método de dos pasos y un dilutor en base a leche descremada (95 ml), yema de huevo (5 ml), fructosa (4.85 g) y etilenglicol (0.1 M). Además, se utilizaron dos antioxidantes análogos de superóxido dismutasa: Tempo (2,2,6,6 tetrametil-1 Piperidiniloxil; Aldrich 214000) y Tempol (4- hidroxi 2,2,6,6 tetrametil-1-piperidiniloxil; Sigma H-8258).
Cada muestra de semen fue dividida en tres porciones para formar los siguientes grupos: grupo control, grupo Tempo (1 mM) y grupo Tempol (1 mM). Los antioxidantes fueron añadidos durante la curva de enfriamiento, cuando las muestras llegaron a los 10 °C. La segunda fracción del dilutor se agregó después del proceso de enfriamiento (de 35 a 5 °C en 90 minutos), y se dejó equilibrar por 30 minutos. Las muestras fueron almacenadas en pajillas de 0.25 mL, expuestas a vapores de nitrógeno líquido por 15 minutos, y finalmente depositadas en nitrógeno líquido. El descongelamiento se realizó en baño María a 38 °C por 1 minuto.
Integridad del ADN Espermático
Las muestras fueron fijadas y permemabilizadas previamente al protocolo de TUNEL. Para la fijación, el contenido de cada pajilla (250 μL) fue lavado mediante centrifugación (500 x G) por 5 minutos en 10 ml de medio Tris base (27.1 g Tris, 14 g ácido cítrico, 10 g fructosa, csp 1 L). Los pellets fueron resuspendidos y fijados agregando 5 ml de formaldehido al 2% diluido en PBS (24 g ClNa, 0.6 g ClK, 0.78 g KH2PO4, 3.3 g Na2HPO4, csp 1 L, pH 7.2), por 40 minutos a temperatura ambiental. Se retiró el fijador, mediante 2 lavados por centrifugación. Para la permeabilización, el pellet fue resuspendido en 100 L de solución Triton X-100 (Sigma T-9284) al 0.8%, diluido en 0.1% de citrato de sodio, por 15 minutos a 5 °C. Luego, se volvió a lavar 2 veces mediante centrifugación.
Se evaluó la integridad del ADN espermático mediante la técnica de TUNEL, utilizando el producto comercial In Situ Cell Death Detection Kit, Fluorescein (Roche Diagnostics, Alemania), siguiendo las indicaciones del fabricante. Los pellets obtenidos luego del procesamiento de la técnica de TUNEL fueron resuspendidos en 300 μL de PBS y colocados en tubos de 6 mL para lectura en citómetro de flujo. Finalmente, 1 L de solución stock (5 mg/mL) de ioduro de propidio (Live/Dead® Sperm Viability Kit, Molecular Probes L-7011, EEUU) fue agregado a cada tubo, con la finalidad de marcar las células permeabilizadas.
La lectura de los tubos, colocados a 4 °C en cámara oscura, se hizo mediante citometría de flujo y microscopía de fluorescencia. Los espermatozoides positivos a TUNEL presentaron una intensa coloración verde bajo microscopía de fluorescencia, mientras que los espermatozoides positivos a ioduro de propidio (PI) presentaron una coloración roja intensa. Asimismo, para definir las poblaciones en función de su fluorescencia a PI/TUNEL, se realizaron los siguientes controles: autofluorescencia (espermatozoides vivos incubados sin TUNEL y sin PI), control TUNEL (espermatozoides vivos tratados con ADNasa e incubados sólo con TUNEL) y control PI (espermatozoides muertos incubados sólo con PI).
Los espermatozoides fueron clasificados en cuatro categorías:
- Permeabilizados con ADN intacto (positivos PI/ negativos TUNEL)
- Permeabilizados con ADN fragmentado (positivos PI/positivos TUNEL)
- No permeabilizados con ADN intacto (negativos PI/negativos TUNEL)
- No permeabilizados con ADN fragmentado (negativo PI/positivo TUNEL).
Citometría de Flujo
La fluorescencia fue detectada utilizado un citómetro de flujo FACSCalibur (Becton Dickinson, CA, EEUU) equipado con óptica estándar. Las células fueron excitadas con un láser argón de longitud de onda de 488 nm, empleando 10 000 espermatozoides como mínimo en cada análisis. Para definir los parámetros de la población de espermatozoides, se realizó un análisis comparando el tamaño (forward-Scatter-Height FSC-H) y la complejidad celular (side-Scatter-height SSC-H). Los datos fueron trabajados en escala logarítmica y analizados utilizando el software Cell-Quest Pro (Becton Dickinson, CA, EEUU). Los resultados se presentaron en gráficos Dot Plots de dos parámetros (PI vs TUNEL, leídos a través de los filtros PE y FITC, respectivamente) y en porcentajes de espermatozoides TUNEL+.
Análisis Estadístico
Se calculó el porcentaje de espermatozoides con ADN fragmentado, en función del total de células permeabilizadas. Estos porcentajes fueron transformados (raíz cuadrada del arcseno) para aproximar los datos a la distribución normal. El efecto de los grupos (Control, Tempo y Tempol) fue analizado utilizando la prueba de ANOVA mediante el software Prism version 3.0 (San Diego, EEUU), y se empleó la prueba de Tukey para determinar diferencias entre grupos.
RESULTADOS
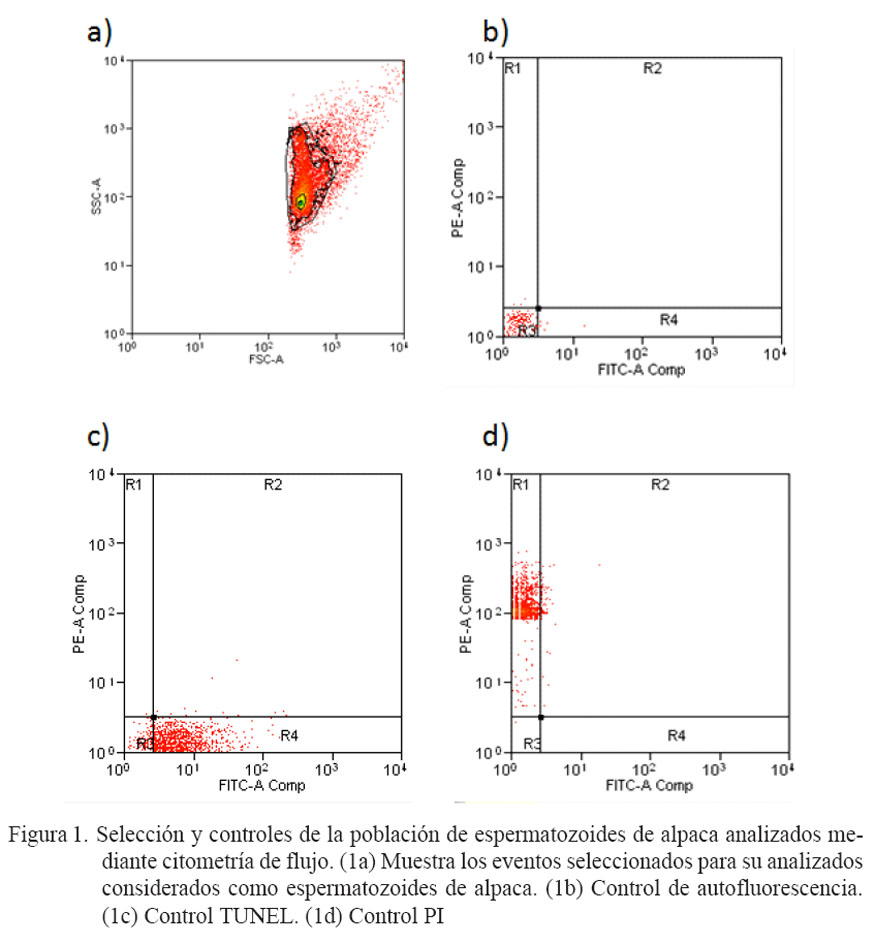
En la Fig. 1a se presenta la población de eventos que fue seleccionada (dentro de la línea negra continua) como espermato zoides de alpaca, de acuerdo a su tamaño (FSC) y complejidad (SSC). Las Figs. 1b, 1c y 1d representan los controles de autofluorescencia, control TUNEL y control PI, respectivamente, que fueron utilizados para definir los cuadrantes (R1, R2, R3 y R4) de las poblaciones espermáticas. Se observa que el control de autofluorescencia (Fig. 1b) no muestra excitación ante la exposición a los filtros para FITC y PE, quedando la mayoría de eventos localizados en el cuadrante R3. En la Fig. 1c, el control TUNEL presenta un distribución hacia el cuadrante R4 (inferior derecho), indicando una mayor excitación con el filtro FITC y ninguna con el filtro PE, donde estas células fueron observadas de color verde intenso bajo microscopía de fluorescencia (Fig. 2d). Por el contrario, el control PI (Fig. 1d) presenta una mayor distribución hacia el cuadrante R1, indicando una mayor excitación con el filtro PE y muy poca con el filtro FITC; asimismo, estas células fueron observadas de color rojo intenso bajo microscopía de fluorescencia (Fig. 2b).
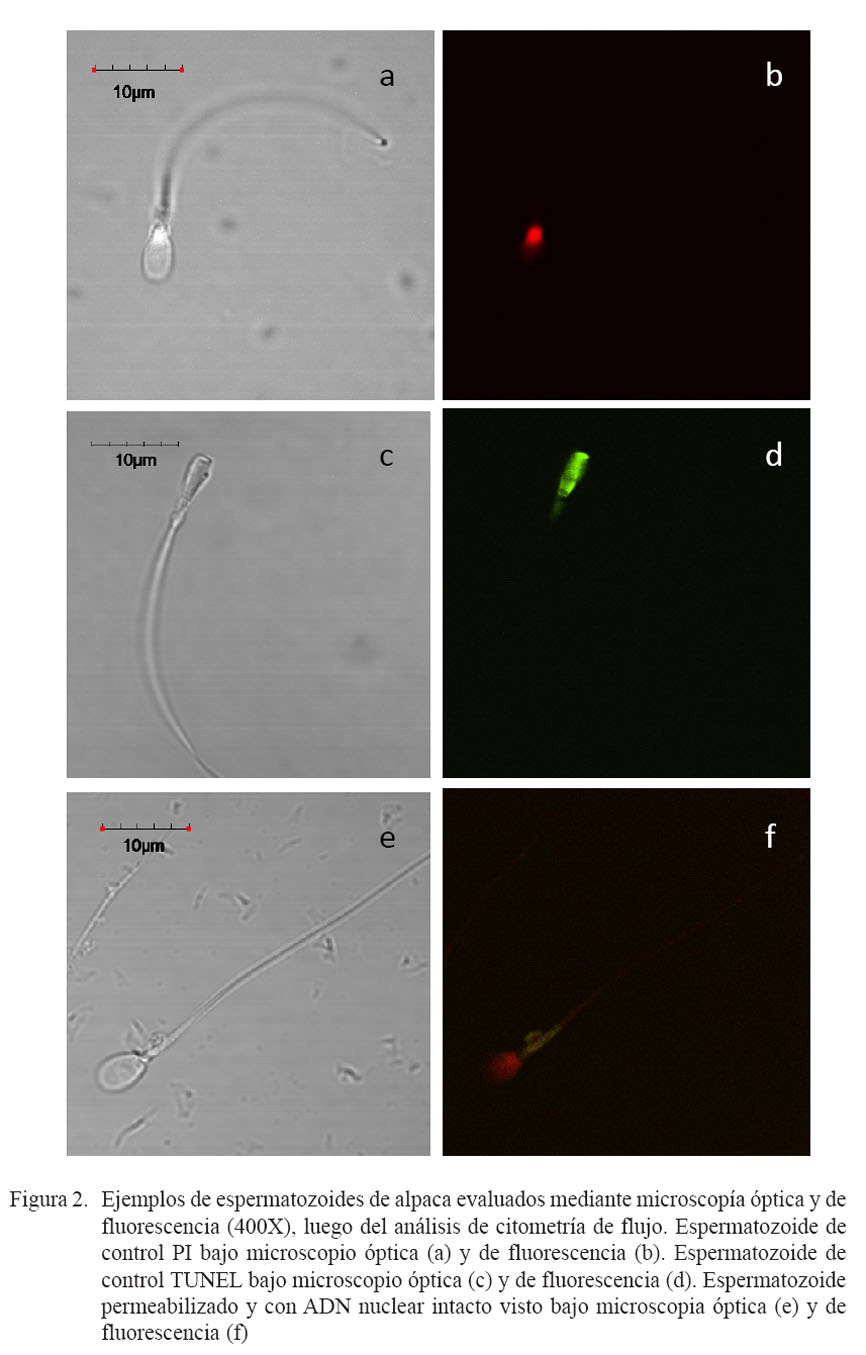
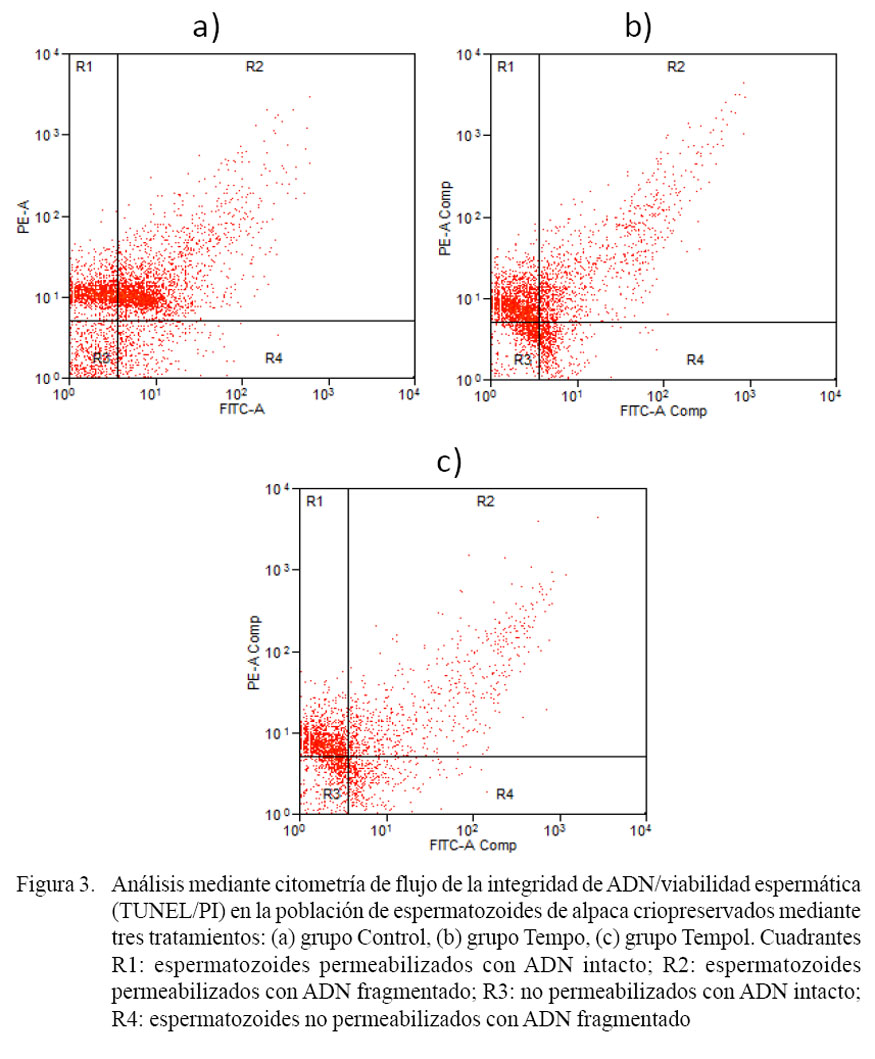
En el análisis de la integridad de ADN, sólo se consideraron los espermatozoides dentro de los cuadrantes R1 y R2, por representar a las células permeabilizadas, las cuales tuvieron la mayor probabilidad de exponerse al reactivo del TUNEL. La frecuencia de espermatozoides con ADN fragmentado fue de 38.8 ± 16.2, 25.4 ± 5.8 y 16.7 ± 8.4% para los grupos Control, Tempo y Tempol, respectivamente. La frecuencia de espermatozoides con ADN fragmentado fue estadísticamente superior en el grupo Control con relación al grupo Tempol (p<0.05). Por otro lado, el grupo Tempo no presentó diferencias significativas en porcentaje de ADN fragmentado en comparación con los grupos Control y Tempol. En la Fig. 2f se puede observar un ejemplo de espermatozoide de alpaca permeabilizado y que presenta el ADN nuclear intacto.
En la Fig. 3 se muestran algunos ejemplos de gráficos Dot Plot correspondiente a los grupos Control, Tempo y Tempol. En (a) se puede observar una mayor distribución de eventos entre los cuadrantes R1 y R2, lo cual representan espermatozoides con ADN intacto y espermatozoides con ADN fragmentados; mientras que en (b) y (c) se observa una mayor concentración de eventos localizados en el cuadrante R1, lo cual representa espermatozoides permeabilizados con ADN intacto.
DISCUSIÓN
Los resultados del presente estudio son los primeros que sugieren un efecto favorable de la utilización de antioxidantes como componentes de los dilutores para prevenir el daño al ADN espermático durante la criopreservación de semen de alpaca. También se presentan por primera vez los resultados del análisis de la fragmentación del ADN en espermatozoides de alpaca mediante el ensayo de TUNEL evaluado por citometría de flujo.
La fragmentación del ADN espermático en el grupo tratado con Tempol, antioxidante análogo de superóxido dismutasa, fue de 16.7%, mientras que se ha reportado una fragmentación de ADN de 13% en espermatozoides frescos de alpaca (Rodríguez, 2009). Por otro lado, la fragmentación del ADN espermático en el grupo de espermatozoides criopreservados sin antioxidantes llegó a 38.8%. Estos resultados indicarían que la utilización del antioxidante Tempol como parte del dilutor para criopreservación de semen de alpaca, evitaría en forma significativa el daño al ADN de espermatozoides de alpaca, alcanzando porcentajes muy similares a los encontrados en muestras de semen fresco.
El incremento significativo en el daño al ADN espermático en caso de no utilizar antioxidantes durante la criopreservación podría atribuirse al aumento significativo de la producción de ROS durante el proceso de criopreservación de los espermatozoides, lo cual ha sido demostrado en otras especies de mamíferos (Wang et al., 1997; Santiani, 2003). Estos posibles niveles elevados de ROS estarían relacionados con la fragmentación del ADN en espermatozoides de alpaca.
El método de TUNEL se basa en que durante la apoptosis o fragmentación del ADN espermático se producen fragmentos de ADN o cadenas simples de ADN, las cuales son factibles de ser detectados mediante la incorporación de nucleótidos marcados con fluoresceína (Gorczyca et al., 1993). Los espermatozoides marcados con la sonda del TUNEL presentan una fluorescencia de color verde, que podría ser evaluada mediante microscopia de fluorescencia o mediante citometría de flujo. En el caso de lecturas mediante microscopía de fluorescencia, se recomienda contar 200 espermatozoides por duplicado para disminuir la probabilidad de error (NAFA y ESHRE, 2002), lo cual puede constituir la evaluación en un proceso largo y tedioso. Por el contrario, la evaluación del ensayo de TUNEL mediante citometría de flujo se realiza en segundos y evaluando un población de 10 000 espermatozoides, constituyendo entonces un análisis más objetivo y replicable (Muratori et al., 2008).
El porcentaje de daño espermático encontrado mediante citometría de flujo puede ser el doble o hasta el triple que los detectados mediante microscopía del fluorescencia (Muratori et al., 2008). Esto podría explicar el 15-20% de ADN fragmentado encontrado en espermatozoides de alpaca por Rodríguez (2009), donde utilizaron TUNEL y microscopía de fluorescencia, en contraste el 16.8-38.8% encontrado en el presente estudio empleando TUNEL y citometría de flujo.
CONCLUSIONES
La adición de Tempol (1 mM) durante la curva de enfriamiento (10 °C) para la criopreservación de semen de alpaca reduce significativamente (p<0.05) la fragmentación del ADN espermático.
LITERATURA CITADA
1. Aitken R, Clarkson J, Fishel S. 1989. Generation of reactive oxygen species, lipid peroxidation and human sperm function. Biol Reprod 40: 183-197.
2. Aitken R, Fisher H. 1994. Reactive oxygen species generation and human spermatozoa: the balance of benefit and risk. Bioessays 16: 259-267.
3. Aitken R, Gordon E, Harkiss D, Twigg J, Milne P, Jennings Z, Irvine D. 1998. Relative impact of oxidative stress on the functional competence and genomic integrity of human spermatozoa. Biol Reprod 59: 1037-1046.
4. Agarwal A, Said T. 2005. Oxidative stress, DNA damage and apoptosis in male fertility: a clinical approach. BJU Int 95: 503-507.
5. Banda J, Evangelista S, Ruiz L, Sandoval R, Rodríguez C, Valdivia M, Santiani A. 2010. Efecto de dilutores en base a Tris, Tes y leche descremada en la criopreservación de espermatozoides obtenidos del epidídimo de alpaca. Rev Inv Vet, Perú 21: 145-153.
6. Baumber J, Ball B, Linfor J, Meyers S. 2003. Reactive oxygen species and cryopreservation promote DNA fragmentation in equine spermatozoa. J Androl 24: 621-628.
7. Bilodeau J, Chatterjee S, Sirard M, Gagnon C. 2001. Levels of antioxidant defenses are decreased in bovine spermatozoa after a cycle of freezing and thawing. Mol Reprod Dev 55: 282-288.
8. Cordelli E, Eleuteri P, Leter G, Rescia M, Spano M. 2005. Flow cytometry applications in the evaluation of sperm quality: semen analysis, sperm function and DNA integrity. Contraception 72: 273-279.
9. De Lamirande E, Leclere P, Gagnon C. 1997. Capacitation as a regulatory event that primes spermatozoa for the acrosome reaction and fertilization. Mol Hum Reprod 3: 175-194.
10. Gorczyca W, Gong J, Darzynkiewicz. 1993. Detection of DNA strand breaks in individual apoptotic cells by the in situ terminal deoxynucleotidyl transferase and nick translation assays. Cancer Res 53: 945-951.
11. Medeiros C, Forell F, Oliveira A, Rodrigues J. 2002. Current status of sperm cryopreservation: why isn´t better?. Theriogenology 57: 327-344.
12. Morton K, Bathgate R, Evans G, Maxwell W. 2007. Cryopreservation of epididymal alpaca sperm: a comparison of citrate- Tris- and lactose-based diluents and pellets and straws. Reprod Fertil Dev 19: 792-796.
13. Morton K, Evans G, Maxwell W. 2010. Effect of glycerol concentration, Equex STM® supplementation and liquid storage prior to freezing on the motility and acrosome integrity of frozen-thawed epididymal alpaca sperm. Theriogenology 74: 311-316.
14. [NAFA y ESHRE] Nordic Association for Andrology y European Society for Human Reproduction and Embriology. 2002. Manual on basic semen analysis. Sweden: NAFA & ESHRESIGA. 33 p.
15. Muratori M, Forti G, Baldi E. 2008. Comparing flow cytometry and fluorescence microscopy for analizing human sperm DNA fragmentation by TUNEL labeling. Cytometry A 73: 785- 787.
16. Peris S, Bilodeau J, Dufour M, Bailey J. 2007. Impact of cryopreservation and reactive oxygen species on DNA integrity, lipid peroxidation and functional parameters in ram sperm. Mol Reprod Dev 74: 878-892.
17. Rodríguez C. 2009. Efecto del plasma seminal sobre la supervivencia de espermatozoides criopreservados de alpaca (Vicugna pacos). Tesis de Magíster. Lima: Universidad Nacional Mayor de San Marcos. 76 p
18. Ruiz L, Santiani A, Sandoval R, Huanca W, Coronado L, Alzamora C. 2007. Efecto de dos antioxidantes (tempo y tempol) en la criopreservación de semen ovino empleando un dilutor en base a tris. Rev Inv Vet, Perú 18: 99-106.
19. Santiani A. 2003. Criopreservación de semen ovino: efecto de la adición de antioxidantes al diluyente. Tesis de Magíster. Temuco, Chile: Univ La Frontera. 95 p.
20. Santiani A, Huanca W, Sapana R, Huanca T, Sepúlveda N, Sánchez R. 2005. Effects on the quality of frozenthawed alpaca (Lama pacos) semen using two different cryoprotectants and extenders. Asian J Androl 7: 303-309.
21. Sue-Hee K, Do-Hyeon Y, Yong-Jun K. 2010. Effects of cryopreservation on phosphatidylserine translocation, intracellular hydrogen peroxide, and DNA integrity in canine sperm. Theriogenology 73: 282-292.
22. Wang A, Shang H, Ikemoto I, Anderson D, Loughlin K. 1997. Reactive oxygen species generation by seminal cells during cryopreservation. Urology 49: 921-925.
