










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versión impresa ISSN 1609-9117
Rev. investig. vet. Perú vol.25 no.3 Lima set. 2014
ARTÍCULOS PRIMARIOS
Expresión In vitro de las interleucinas 2 y 10 de linfocitos de alpacas (Vicugna pacos) en presencia de antígenos clostridiales
In vitro expression of interleukin-2 and -10 from alpaca (Vicugna pacos) leukocytes in presence of clostridial antigens
Raquel Watanabe W.1, Alberto Manchego S.1,2, Hermelinda Rivera G.1
1 Laboratorio de Microbiología y Parasitología Veterinaria, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima
2 E-mail: amanchegos@gmail.com
RESUMEN
El objetivo del presente estudio fue determinar, mediante la técnica de RT-PCR Tiempo Real y cuantificación relativa según el método 2-rrCt, los niveles relativos de expresión in vitro de ARN mensajero de IL-2 e IL-10 en leucocitos circulantes de alpacas en presencia de antígenos clostridiales, empleándose como control endógeno a GAPDH. Se colectó sangre entera de 10 alpacas adultas. Las muestras fueron centrifugadas para obtener la fracción leucocitaria en gradiente de Ficoll, colectándose la capa flogística. Se hizo un pool de leucocitos que fueron purificados con cloruro de amonio. Se centrifugó yla concentración se ajustó a 500 000 células vivas/ml de medio mínimo esencial (MEM). Fueron cultivados en placas para cultivo celular de 24 pocillos y enfrentados a suspensiones de extracto clostridial a las concentraciones de 400, 16, 0.8 y 0.2 µg/ml por 1, 12 y 24 h. Posteriormente, se realizó la RT-qPCR. La cinética de la expresión de IL-2 mostró una tendencia no significativa inversamente proporcional a la dosis de antígeno clostridial en los tres tiempos de incubación, a diferencia de IL-10 cuya expresión fue variable, alcanzando el máximo a las 12 h de incubación (p<0.05) y mostrandoun patrón similar al de IL- 2 en la dosis de 16 µg/ml de antígeno clostridial. La expresión de IL-10 superó hasta en 40 veces lo expresado por el calibrador respecto a IL-2, evidenciándose una marcada res- puesta hacia la polarización del subtipo Th2.
Palabras clave: interleucina 2, interleucina 10, Clostridium perfringens, RT-PCR tiempo real, cuantificación relativa
ABSTRACT
The aim of the study was to determine the relative in vitro ARNm expression levels of IL-2 and IL-10 in peripheral blood leukocytes by real time RT-PCR and relative quantification using the 2-rrCt method in presence of clostridial antigens and GAPDH as an endogenous control. Whole blood was collected from 10 adult alpacas. Samples were centrifuged to obtain leukocytes using the Ficoll reagent, purified with ammonium chloride and cultured in 24-well plates at a concentration of 500 000 live cells/ml minimal essential medium (MEM) with clostridial extract suspensions at concentrations of 400, 16, 0.8 and 0.2 µg/ml. Subsequently, the RT-qPCR test was performed. IL-2 kinetic expression patterns were inversely proportional to the clostridial antigen dose in the three incubation times (p>0.05), unlike IL-10 which its kinetic expression patterns were variable, reaching its maximum at 12 h (p<0.05) while showing a similar trend like IL-2 in the 16 µg/ml clostridial antigen dose. IL-10 expression exceeded 40-fold the control expression respect to IL-2. A strong polarization towards the Th2 subtype was shown.
Keywords: interleukin-2, interleukin-10, Clostridium perfringens, real time RT-PCR, relative quantification
INTRODUCCIÓN
La enterotoxemia, denominada también diarrea bacilar, es ocasionada por Clostridium perfringens y es la enfermedad infecciosa responsable de las mayores pérdidas económicas que afligen a los productores alpaqueros del Perú. La enfermedad afecta, principalmente, a las crías de las alpacas (Vicugna pacos) entre la segunda y novena semana de edad, incluyendo complicaciones con cuadros de colibacilosis (dia- rrea y septicemia) y de neumonía aguda. El cuadro patológico es consecuencia de un cuadro tóxico primario a nivel intestinal (yeyuno e íleon) que deriva en un cuadro de toxemia generalizada por acción de las toxinas de C. perfringens tipo A que ocasionan daños irreversibles en el endotelio vascular y el sistema nervioso. Generalmente, los signos clínicos son de curso agudo y finalizan con la muerte súbita del animal. La enfermedad se observa como brotes o epizootias, con mortalidades que pueden alcanzar el 50% de las crías (Ramírez et al., 1985) o más si se trata de casos severos (FAO, 2005a,b).
Las interleucinas o citoquinas desempeñan un rol importante en la defensa del organismo de humanos y animales frente a in- fecciones o lesiones; por lo tanto, el desarrollo de las enfermedades puede ser afectado o condicionado por sus respuestas. Su ac- ción en la mediación de la respuesta inmune ha sido ampliamente investigada, especial- mente, el patrón de respuesta de las subpoblaciones de linfocitos T CD4+ Th1 (IL- 2, IFN-γ, IL-12) y Th2 (IL-4, IL-10, IL-13). La dinámica de la expresión del ARN men- sajero (ARNm) de estas interleucinas ha sido estudiada en camélidos del Viejo Mundo (Camelus dromedarius y C. bactrianus) y del Nuevo Mundo como la llama (Lama glama) (Odbileg et al., 2005a,b). Asimismo, han sido clonadas, secuenciadas (Odbileg et al., 2006) y evaluadas en condiciones in vivo frente a antígenos de Brucella abortus (Odbileg et al., 2008).
Estudios a nivel molecular de perfiles de interleucinas de subpoblaciones linfocitarias Th1 y Th2 de llamas y camellos bactrianos, especies filogenéticamente emparentadas con las alpacas, han logrado examinar la expresión de los genes de las interleucinas involucradas en el balance de la respuesta Th1/Th2, revelando respuestas funcionalmente complejas en procesos infecciosos sistémicos. No obstante, son escasos los estudios sobre la inmunidad de la alpaca, habiéndose extrapolado el diseño de los oligonucleótidos empleados en base a las secuencias conservadas y homólogas con otras especies. La cinética de la expresión in vitro de algunas interleucinas (IL) de alpacas se ha estudiado en los últimos años, especialmente en la respuesta inmune innata (IL-1a, IL-1b, IL-6, TNF-a) (Bardález et al., 2012; Tambillo et al., 2012) y adaptativa (IL-2, IFN- γ, IL-4, IL-10) (Chiok, 2012).
El presente estudio tuvo como objetivo evaluar la expresión in vitro de las IL-2 e IL-10 en leucocitos circulantes de alpacas incubados con antígenos clostridiales, estableciendo la cinética de producción de los ARNm de estos dos elementos y, de esta manera, brindar una aproximación al com- portamiento in vivo de los leucocitos sanguíneos en los casos de toxemia generalizada causada por C. perfringens en crías de alpacas.
MATERIALES Y MÉTODOS
Obtención y Procesamiento de Muestras
Se tomaron muestras de sangre con EDTA de 10 alpacas adultas mediante punción de la vena yugular. Las muestras se centrifugaron a 1000 g por 5 min para obtener la fracción leucocitaria en gradiente de Ficoll. Se hizo un pool de leucocitos y se purificaron mediante lisis de eritrocitos con cloruro de amonio, y luego fueron resuspendidos en medio mínimo esencial (MEM) libre de antibióticos.
Antígenos Clostridiales
Los antígenos de C. perfringens se obtuvieron de cepas de campo procedentes de crías de alpacas muertas con cuadros de enterotoxemia. La cepa fue cultivada en caldo tioglicolato en anaerobiosis e inactivada con formalina 0.05%. Se precipitaron las proteínas totales con ácido tricloroacético (ATC) y se lavaron los restos con acetona. La dosis de antígeno se ajustó a una solución madre de 400 µg/ml empleando un fluorómetro Qubit (Invitrogen, EEUU). Posteriormente, se rea- lizaron diluciones con PBS para obtener concentraciones de 80, 16, 0.8 y 0.2 µg/ml.
Activación Leucocitaria
Las suspensiones de antígenos clostridiales en las cinco concentraciones (so- lución madre más cuatro diluciones) fueron incubadas con leucocitos de alpaca (500 000 células/ml) a 37 °C con 5% de CO2 en microplacas de poliestireno para cultivo celular de 24 pocillos por 1, 12 y 24 h. Adicionalmente, y por cada periodo de tiem- po, se empleó un control de células o calibrador inoculado con suero fisiológico e incubado bajo las mismas condiciones de los tratamientos. Luego de la incubación, se to- maron alícuotas de 300 µl y se separaron las células mediante centrifugación a 1000 g por 10 min. Se hizo un recuento de leucocitos de cada tratamiento empleando azul de tripán 0.5% para determinar su viabilidad. La acti- vación de los leucocitos en cada caso fue detenida mediante congelación a -20 °C.
Extracción del ARN Total y Transcripción Inversa
Las muestras fueron descongeladas y centrifugadas a 1500 g por 5 min. Se eliminó el sobrenadante y al pellet se le agregó Trizol (InvitrogenTM Life Technologies, EEUU), si- guiendo las instrucciones del fabricante, y clo- roformo absoluto frío. El ARN fue precipita- do con alcohol isopropílico frío y lavado con etanol frío al 70%, luego fue resuspendido con 60 µl de agua libre de nucleasas y conge- lado a -70 °C hasta su uso.
El ARN obtenido de cada muestra fue tomado como templado para la síntesis del ADN complementario (ADNc) utilizándose el kit SuperScript™ III First-Strand Synthesis SuperMix for qRT-PCR (InvitrogenTM, EEUU) usando oligos dT y hexámeros al azar como cebadores.
PCR Tiempo Real para Genes GAPDH, IL-2 e IL-10
Se emplearon los oligonucleótidos dise- ñados por Odbileg et al. (2006, 2008), basa- dos en secuencias genéticas para bovino y camello para los genes de IL-2 e IL-10 (Cuadro 1). Asimismo, se incluyeron oligonu-cleótidos específicos para la amplificación específica de transcritos del gen GAPDH (gliceraldehído 3 – fosfato deshidrogenasa) (Patil et al., 2004) como control endógeno para la normalización de la RT-PCR. Se empleó el kit SuperScriptTM III First-Strand Synthesis SuperMix for qRT-PCR (Invitrogen, EEUU), siguiendo las recomendaciones del fabricante.
Las condiciones del PCR Tiempo Real para GADPH, IL-2 e IL-10 en el termociclador PTC 200 Engine Chromo IV (MJ Research, EEUU) fueron las siguientes: 50 °C por 2 min (incubación con UDG), 95 °C por 10 min (inactivación del UDG y activación de la ADN polimerasa), seguido de 40 ciclos de 94 °C por 30 s, 55 °C por 30 s y 72 °C por 30 s (extensión final) y lectura de placa, curva de disociación de 65 hasta 95 °C con lectura de placa cada 0.3 °C, y luego a 4 °C indefinidamente para mantener los productos. Los tubos fueron refrigerados a 4 °C a la finalización del proceso (stop).
El ensayo fue desarrollado por triplica- do y los resultados fueron evaluados mediante el software Opticon Monitor v. 2.1, obteniéndose los valores de las curvas de amplificación (Ct, cycle treshold) y disocia- ción (Tm, temperature melting). La cuantificación relativa de los amplicones de ARNm de las interleucinas 2 y 10 se realizó fabricante.
Los datos fueron analizados con el software STATA SE 12 utilizando las pruebas no paramétricas de Kruskal Wallis y Compara- ción Múltiple de Medianas. Adicionalmente, se realizó un análisis de efecto simple para determinar diferencias estadísticas entre tratamientos (dosis de antígeno clostridial) según el tiempo (periodos de incubación) y viceversa.
RESULTADOS
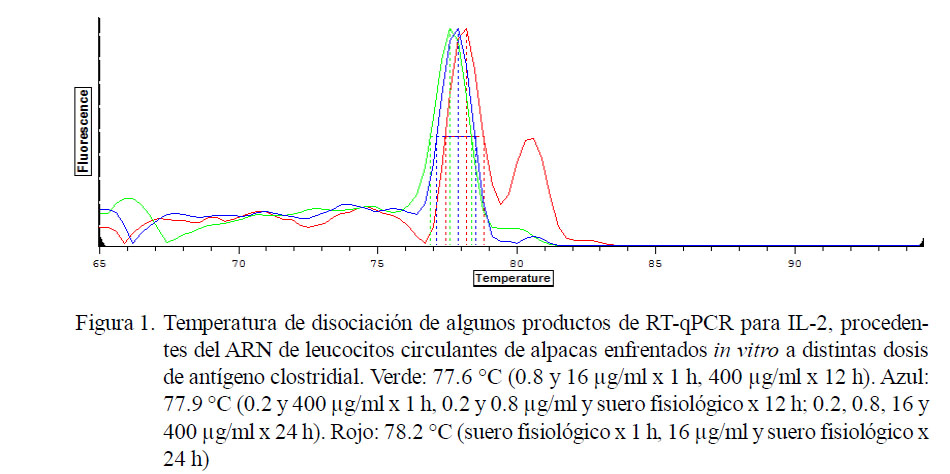
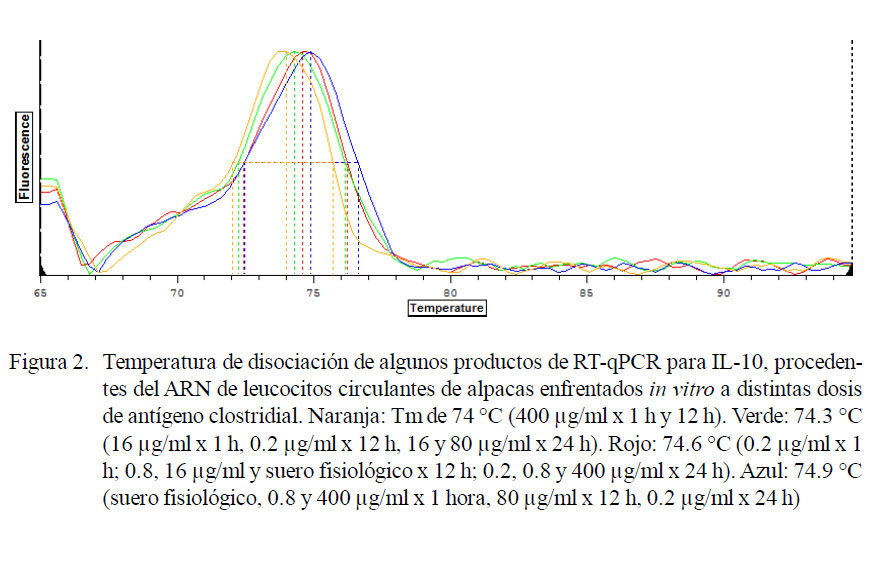
Se encontraron cinco temperaturas de disociación para GAPDH (82.1. 82.7, 84.2, 84.5, 84.8 °C). Asimismo, tres temperaturas de disociación con leve ruido de fondo para IL-2 e IL-10 de 77.6, 77.9 y 78.2 °C (Fig. 1) y 74.0, 74.3, 74.6 y 74.9 °C (Fig. 2), respectivamente.
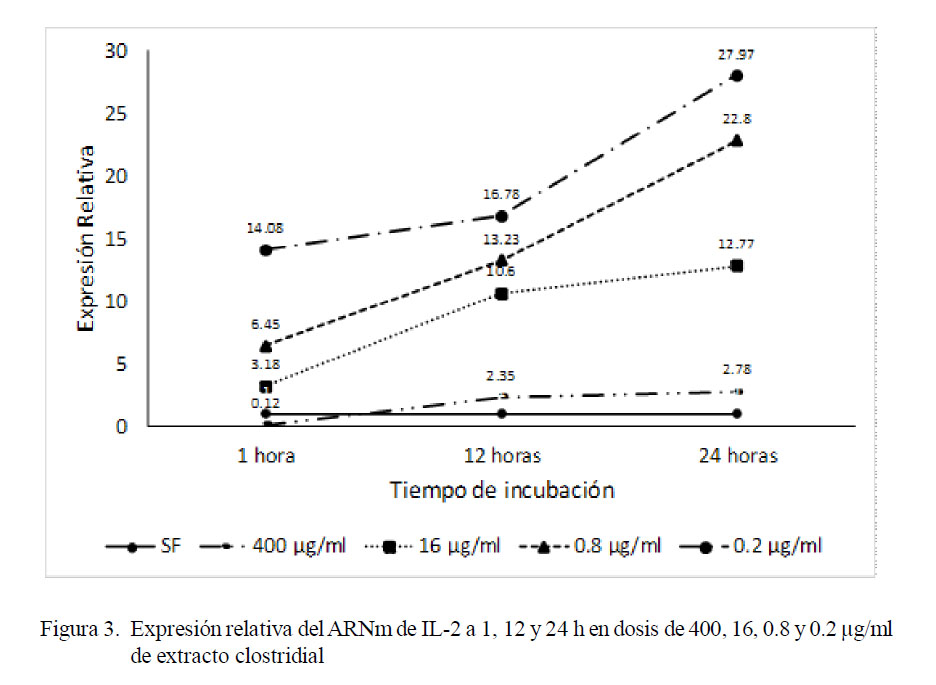
El análisis realizado para comparar la expresión de IL-2 en los tres periodos de incubación de cada tratamiento reveló que todas las dosis de antígeno clostridial favore- cieron el incremento de la expresión relativa del ARNm de IL-2 superando al control, a excepción de la dosis máxima de 400 µg/ml. En la Fig. 3 se puede notar la tendencia progresiva en incremento de la expresión de Il-2, conforme aumenta la dosis de antígeno clostridial y el tiempo de incubación. El análisis de normalidad mediante la prueba de Shapiro-Wilk estableció que las variables se- guían una distribución normal. La prueba de Kruskal Wallis no determinó diferencia esta- dística entre tratamientos según la dosis de antígeno clostridial y los tiempos de incubación; sin embargo, el análisis factorial encontró diferencia estadística entre trata- mientos a las 24 h de incubación y a las dosis de 0.8 y 0.2 ìg/ml.
En forma similar, el análisis realizado para comparar la expresión de IL-10 en los tres periodos de incubación de cada trata- miento reveló que los niveles de expresión fueron variables dependiendo de la dosis de antígeno clostridial y el tiempo de incubación. En la Fig. 4 se observan los picos (máxima expresión) a las 12 h de incubación en las concentraciones de 0.2, 0.8 y 400 µg/ml, tendencia que no se evidencia a la concentración de 16 µg/ml. Por otro lado, solo en la de 16 µg/ml se observa un incremento progresivo en la expresión de esta interleucina conforme aumenta el tiempo de incubación, mientras que en las dosis de 400, 0.8 y 0.2 µg/ml, la expresión de IL-10 aumenta a las 12 h pero disminuye marcadamente a las 24 h de incubación.
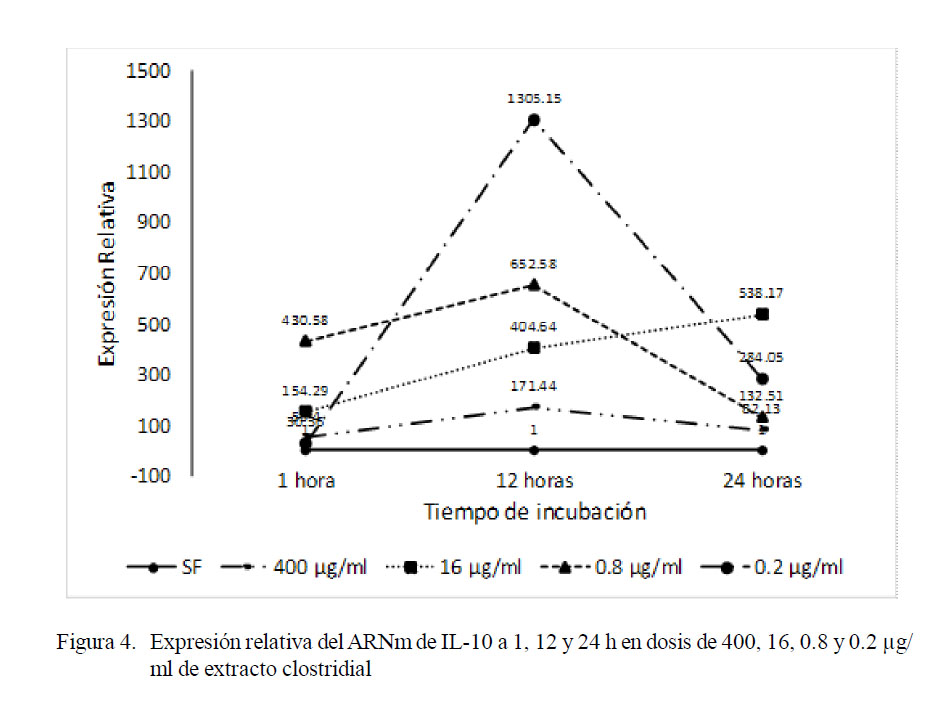
El análisis de normalidad mediante la prueba de Shapiro-Wilk estableció que las variables no seguían una distribución normal. La prueba de Kruskal Wallis determinó diferencia estadística entre los tratamientos a las 12 h (p=0.0444) y 24 h (p=0.0361) de incubación. No obstante, la prueba de Comparación Múltiple de Medianas solo determinó diferencia estadística entre 400 y 0.2 µg/ml de antígeno clostridial a las 12 h de incubación. Por otro lado, el análisis factorial indicó diferencia estadística entre tratamientos a las 12 y 24 h de incubación, y a las dosis de 16, 0.8 y 0.2 µg/ml.
La especificidad de los productos de la PCR tiempo real para IL-2 e IL-10 fue corroborada con la electroforesis de los productos en gel de agarosa mostrando el tamaño indicado por Odbileg et al. (2005, 2008).
DISCUSIÓN
El presente estudio cuantificó relativa- mente la expresión del ARN mensajero de las interleucinas 2 y 10 in vitro en leucocitos circulantes de alpacas incubados con cuatro dosis de extracto de C. perfringens por 1, 12 y 24 h, siendo la IL-2 una interleucina re- presentativa de la subpoblación linfoide Th1 y la IL-10 de la subpoblación Th2, ambas correspondientes al conjunto de linfocitos T CD4+.
Se observó un incremento progresivo en la expresión de IL-2 inversamente proporcional a la concentración de antígeno clostridial y directamente proporcional al tiempo de incubación, a excepción de la dosis de 400 µg/ml en 1 h, donde la expresión equiva- le a la décima parte respecto al grupo control inoculado con suero fisiológico. Esto indicaría una menor producción de esta interleucina debido a la actividad leucotóxica del extracto (Tambillo et al., 2012), dado que la bacteria secreta grandes cantidades y tipos de exotoxinas con actividades enzimáticas y porinas. Esta actividad citotóxica no es tan marcada en las tres dosis menores debido a una mayor cantidad de células viables (datos no mostrados) y, por lo tanto, una mayor estimulación autocrina y paracrina entre las mismas, lo que refuerza la producción total de esta interleucina. No obstante, no se determinó la dosis mínima de extracto que estimularía la síntesis de IL-2 por los leucocitos periféricos.
En el caso de IL-10, los patrones en la cinética de la expresión del ARNm a distintas dosis de extracto de C. perfringens varían en los tres tiempos de incubación. Al igual que en IL-2, no se determinó la dosis mínima de extracto capaz de estimular la síntesis de IL-10 en los leucocitos periféricos.
En términos generales, los niveles de expresión de IL-2 superan en menos de 30 veces lo expresado por el calibrador, siendo el máximo de 27.97; mientras que, en IL-10, la mínima y máxima expresiones corresponden a 30.36 y 1305.15 veces lo expresado por el calibrador a la 1 y 12 h de incubación, respectivamente, a la mínima concentración de antígeno clostridial (0.2 µg/ml). La interpretación de estos eventos dispares debe ser analizada considerando el periodo de incubación para ambas interleucinas que radica en las interacciones inmunológicas que favorecen el predominio de un subtipo de linfocitos T CD4+ y, por lo tanto, de un perfil de interleucinas de la inmunidad adaptativa.
La interleucina más importante producida por los linfocitos T poco después de su activación, a menudo en las 2 a 4 h siguientes al reconocimiento del antígeno y de los coestimuladores, es la IL-2 (Abbas et al., 2012), hecho que explica la cinética uniforme de la expresión de la interleucina en los tres tiempos de incubación a medida que disminuye la concentración de antígeno clostridial.
Los macrófagos responden a los microorganismos secretando interleucinas y expresando coestimuladores que potencian la activación de los linfocitos T y la inmunidad celular. La IL-10 actúa sobre los macrófagos activados bajo dos formas. La primera es la inhibición de la síntesis de IL-12 por las célu- las dendríticas y macrófagos activados (Abbas et al., 2012). La IL-12 es un estímulo crítico para la secreción de IFN-γ y es un inductor de las relaciones inmunitarias innatas y celu- lares frente a los microorganismos intracelulares. En consecuencia, la IL-10 ac- túa inhibiendo estas reacciones, a la vez que favorece indirectamente la polarización del subgrupo Th2 sobre el Th1 por inhibición de esta interleucina. La segunda consiste en la inhibición de la expresión de coestimuladores y moléculas del CMH clase II sobre los macrófagos y células dendríticas (Abbas et al., 2012). Por lo tanto, la IL-10 actúa inhibiendo la activación de los linfocitos T y finalizando las reacciones inmunitarias celu- lares a través de los linfocitos T reguladores.
Por otro lado, la IL-10 inhibe la activi- dad de linfocitos NK (células citotóxicas na- turales) y la síntesis de las interleucinas de linfocitos Th1 (IL-1, IFN-γ y TNF-b), a la vez que actúa en macrófagos activados para suprimir la secreción de IL-1, IL-6, TNF-a y radicales reactivos de oxígeno (Tizard, 2009). Es probable que los resultados de Tambillo (2012) se hayan debido, fundamentalmente, a la acción de esta interleucina sobre la pro- ducción de IL-1, IL-6 y TNF-a, cuyas ex- presiones de ARN mensajero también fue- ron evaluadas a las mismas dosis de antígeno clostridial por los mismos tiempos de incubación.
Estos resultados contrastan con los ha- llazgos de Wallace et al. (1999) en un estu- dio in vitro e in vivo de la respuesta de cinco interleucinas (IL-1, IL-6, TNF-a, IFN-γ e IL-2) sometidas a la enterotoxina de C. perfringens (CPE), donde se observó la ac- tividad mitogénica de la enterotoxina al indu- cir la síntesis de IL-1, IL-6, TNF-a e IFN-γ por los macrófagos, mas no de IL-2 dentro de las 24 h de incubación y muerte celular total a las 48 h. Esos hallazgos difieren con aquellos del presente estudio en cuanto a la producción de Il-2, donde se evidencia am- plificación de ARNm de esta interleucina. No obstante, debe mencionarse que en ese estu- dio se empleó un kit de ELISA para la cuantificación absoluta y se basó, principal- mente, en la evaluación de la interacción de la respuesta inmuneinnata y no dela adquirida.
Es evidente que en el presente estudio existe un predominio en la polarización hacia el subtipo Th2 en comparación al Th1, ten- dencia que se observa en los tres periodos de incubación, no solo por el efecto de IL-10 sobre IL-2, sino además, por interacciones paralelas orquestadas de otras interleucinas específicas de ambos subgrupos como IL-4 e IFN-γ, y que no fueron evaluadas. Es sabi- do que estas dos interleucinas se inhiben mu- tuamente, aunque la ausencia de una no im- pida la regulación de la otra, como ha sido demostrado para IL-4 en relación a IFN-ã (Yagi et al., 2010). Estos hallazgos contras- tan con los resultados de estudios in vitro en mucosa intestinal de alpacas (Chiok, 2012; More, 2013), en los que se observa una pola- rización hacia el fenotipo Th1, lo que podría sugerir la influencia e interacción con com- ponentes intrínsecos inmunes del microambiente intestinal y la microflora.
Los linfocitos Th2 presentan recepto- res para IL-1 y pueden responder a la IL-1 coestimuladora procedente de los macrófagos o de las células dendríticas. La IL-1 puede secretarse a los fluidos tisulares (IL-1a) o permanecer unida a la superficie celular (IL- 1b), donde estimulará a los linfocitos T uni- dos (Tizard, 2009). Esta característica singu- lar puede sugerir una lógica en los hallazgos de Tambillo et al. (2012) con los del presente estudio, donde los niveles de expresión de ARNm de IL-1a e IL-1b se mantuvieron por debajo del calibrador a la 1 y 12 h de incubación (con excepción de las 24 h). Por lo tanto, puede asumirse que esta interleucina, aún en niveles pequeños, ejerció un efecto de retroalimentación positiva en las células Th2 reforzando sus acciones biológicas. Ade- más, si sus niveles de expresión no fueron tan marcados, pudo deberse a la acción pa- ralela ejercida por IL-4 al bloquear la síntesis de IL-12, suprimiendo la polarización hacia células Th1.
Los valores y variaciones de tempera- tura en las curvas de disociación de los pro- ductos amplificados se encontraron también en el estudio de Chiok (2012). Podría tratar- se de variaciones en la composición de bases en los productos amplificados como resulta- do del muestreo de alpacas procedentes de zonas geográficas distintas, así como a híbridos de camélidos sudamericanos con si- milar fenotipo pero diferente genotipo (Kadwell et al., 2001). Una explicación adi- cional para estos hallazgos podría ser atribui- da a los polimorfismos genéticos, específi- camente aquellos de nucleótido simple (SNPs) (Vignal et al., 2002). También puede ser el resultado de mecanismos que regulan la expresión génica en su etapa postranscripcional como es el splicing «corte y empalme» alternativo y la edición del ARNm (Black, 2003).
En el presente estudio no se pudo de- terminar fehacientemente la existencia de polimorfismos o isoformas de ARNm de IL- 2 ni de IL-10 en alpacas, lo que solo sería posible mediante el secuenciamiento nucleotídico de los productos de la RT-PCR Tiempo Real y análisis funcionales posterio- res de tales variaciones, considerando que la secuenciación del ADN es el gold standard para la detección de polimorfismos.
Existe una baja probabilidad de haberse cuantificado formas de ARNm inmaduro o pre-ARNm, llamado también ARNm precur- sor, debido a que tales formas suelen tener diversos tamaños en longitud pares base (pb) (Sánchez-Sánchez y Mittnacht, 2008). Tales formas hubieran sido visibles en los geles de agarosa realizados sobre los productos fina- les como bandas múltiples de diferentes ta- maños; sin embargo, todos los geles evalua- dos demostraron un solo producto de tamaño compatible con el cDNA del ARNm secuenciado por Odbileg et al. (2006 y 2008), descartando esta posibilidad (Chiok, 2012).
Los resultados obtenidos en el presente estudio usando los oligonucleótidos para el gen GAPDH evidenciaron la presencia de cinco productos amplificados en un rango de 82.1 a 84.8 °C, los cuales variaban en 0.3 °C, encontrándose una temperatura de disociación predominante de 82.7 °C (33.2%), seguida por una de 82.1 y otra de 84.2 °C (26.7%) y dos temperaturas de 84.5 y 84.8 °C (6.7%). Esto concuerda con otros resultados realizados en mucosa intestinal de crías de alpaca (More et al., 2011; Bardález et al., 2012). Es posible que esto se deba, al igual que en el caso de IL-2 e IL-10, a polimorfismos de nucleótido simple (SNPs) o a la regulación post-transcripcional del ARNm. El presente estudio no determina la existencia de relación entre las posibles variantes y factores geográficos o poblacionales, ni las consecuencias de la expresión de uno u otro polimorfismo de GAPDH.
CONCLUSIONES
-
Los leucocitos sanguíneos de alpacas estimulados por antígenos clostridiales expresaron ARN mensajero de IL-2 e IL-10.
-
Se observó un incremento gradual no significativo en la expresión del ARN mensajero de IL-2 inversamente proporcional a la concentración de antígeno clostridial en los tres tiempos de incubación y una cinética variable en IL- 10, observándose una expresión gradual a la dosis de 16 µg/ml (p<0.05) y la máxima expresión a la dosis mínima de extracto clostridial a las 12 h de incubación (p<0.05).
-
Se encontró una polarización de la respuesta inmune hacia el fenotipo Th2.
LITERATURA CITADA
1. Abbas AK, Lichtman AH, Pillai S. 2012. Inmunología celular y molecular. 7a ed. Madrid: Elsevier. 212 p. [ Links ]
[ Links ]
[ Links ]
[ Links ]
[ Links ]
[ Links ]
[ Links ]
[ Links ]
[ Links ]
10. More J. 2013. Efecto de antígenos clostridiales con ácido retinoico sobre la expresión de citoquinas de la respuesta inmune humoral y celular de la mucosa intestinal de crías de alpacas (Vicugna pacos). Tesis de Magíster. Lima: Univ Nacional Mayor de San Marcos. 104 p. [ Links ]
11. Obdileg R, Konnai S, Usui T, Ohashi K, Onuma M. 2005a. Quantification of llama inflammatory cytokine mRNAs by real-time RT-PCR. J Vet Med Sci 67: 195-198. [ Links ]
12. Obdileg R, Konnai S, Ohashi K, Onuma M. 2005b. Molecular cloning and phylogenetic analysis of immfla- matory cytokines of Camelidae (llama and camel). J Vet Med Sci 67: 921-925. [ Links ]
13. Odbileg R, Purevtseren B, Batsukh Z, Konnai S, Ohashi K, Onuma M. 2006. Complete cDNA sequences and phylogenetic analysis of the Th1 and Th2 cytokines of the bactrian camel (Camelus bactrianus). J Vet Med Sci 68: 941-946. [ Links ]
14. Odbileg R, Purevtseren B, Gantsetseg D, Boldbaatar B, Buyannemekh T, Galmandakh Z, et al. 2008. Cytokine responses in camels (Camelus bactrianus) vaccinated with Brucella abortus strain 19 vaccine. J Vet Med Sci 70: 197-201. [ Links ]
15. Patil A, Hughes AL, Zhang G. 2004. Rapid evolution and diversification of mammalian a-defensins as revealed by comparative analysis of rodent and pri- mate genes. Physiol Gen 20: 1-11. [ Links ]
16. Pfaffl, MW. 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29: 2002-2007. [ Links ]
17. Ramírez A, Huamán D, Ellis R. 1985. Enterotoxemia de la alpaca. INIPA-Programa colaborativo de apoyo a la investi- gación en rumiantes menores. Colorado State University. Serie de Reportes Técnicos 63: 1-40. [ Links ]
18. Rebrikov DV, Trofimov D. 2006. Real- Time PCR: A review of approaches to data analysis. Appl Biochem Microbiol 42: 455-463. [ Links ]
19. Sánchez-Sánchez F, Mittnacht S. 2008. Nonsense-mediated decay: paving the road for genome diversification. Bioessays 30: 926-928. [ Links ]
20. Tambillo L, Manchego A, Chiok KL, Sandoval N, More J, Rivera H. 2012. Evaluación in vitro de la respuesta leucocitaria de alpacas (Vicugna pacos) en presencia de antígenos clostridiales. Rev Inv Vet Perú 24: 510-523. [ Links ]
21. Tizard IR. 2009. Introducción a la inmunología veterinaria. 8ª ed. Barcelona: Elsevier Saunders. 592 p. [ Links ]
22. Vignal A, Milan D, SanCristobal M, Eggen A. 2002. A review on SNP and other types of molecular markers and their use in animal genetics. Genet Sel Evol 34: 275-305. [ Links ]
23. Yagi R, Junttila IS, Wei G. 2010. The Transcription Factor GATA3 actively represses RUNX3 protein-regulated production of interferon-γ. Immunity 32: 507-517. [ Links ]
24. Wallace FM, Mach AS, Keller AM, Lindsay JA. 1999. Evidence for Clostridium perfringens enterotoxin (CPE) inducing a mitogenic and cytokine response in vitro and a cytokine response in vivo. Curr Microbiol 38: 96-100. [ Links ]

