










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versão impressa ISSN 1609-9117
Rev. investig. vet. Perú vol.28 no.2 Lima abr./jun. 2017
http://dx.doi.org/10.15381/rivep.v28i2.13070
http://dx.doi.org/10.15381/rivep.v28i2.13070
ARTÍCULOS PRIMARIOS
Efecto de la testosterona exógena sobre las adherencias pene-prepuciales y la producción de fibra en Alpacas Huacaya
Effect of the exogenous testosterone on the penis-prepucial adherences and fiber production in Huacaya Alpacas
Alex Oscanoa M.1, Víctor Leyva V.1,4, Wilber García V.2, Rosario Gonzáles De La Cruz M.1, Virgilio Alarcón B.3
1 Laboratorio de Reproducción Animal, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima, Perú
2 Estación Experimental del Centro de Investigaciones IVITA-Maranganí, Universidad Nacional Mayor de San Marcos, Cusco, Perú
3 Centro de Investigación de Camélidos Sudamericanos, Universidad Nacional de San Antonio Abad del Cusco, Cusco, Perú
4 E-mail: vleyva@vet.unmsm.edu.pe
RESUMEN
Se evaluó el efecto de la testosterona sobre las adherencias pene-prepuciales y producción de fibra. Se usaron 30 alpacas de 18 meses de edad, distribuidas al azar en tres grupos: machos sin tratamiento (T0), machos tratados con enantato de testosterona (0.5 mg/kg/peso vivo) cada dos semanas durante 12 semanas (T2E), y machos castrados (C0). Las adherencias pene-prepuciales, tamaño testicular, comportamiento sexual y peso corporal se evaluaron cada dos semanas, y muestras de fibra se recolectaron al inicio del estudio y al final de tres periodos experimentales. Se tuvo una fase pre-experimental de cuatro semanas donde se evaluaron los parámetros en estudio para ajustar los datos de la fase experimental. La liberación de las adherencias pene-prepuciales fue mayor en T2E que en T0y C0 entre la semana 8 (p<0.001) y la semana 12 (0.006>p<0.001). El incremento testicular (largo, ancho y grosor) fue mayor en T0 que en T2E entre las semanas 4 y 12 (0.01>p<0.0001). El diámetro de fibra fue similar entre T2E y T0, pero en ambos fue mayor que en C0 (p<0.05). Los pesos de fibra sucia y limpia fueron similares en todos los grupos. El rendimiento de fibra limpia fue menor en T2E que en T0 y C0 en los periodos II y III (0.04>p<0.0001). El peso corporal fue mayor en T2E que en T0 y C0 (p<0.05). Se concluye que la administración de testosterona a alpacas machos jóvenes fue efectiva en la liberación de las adherencias pene-prepuciales e incremento del peso corporal, pero redujo el tamaño testicular y rendimiento de fibra limpia; sin embargo, no afectó los pesos de fibra sucia y limpia ni el diámetro de fibra.
Palabras clave: alpaca, testosterona, adherencias pene-prepuciales, fibra
ABSTRACT
The effect of testosterone on penis-prepucial adherences and production of fibre was studied in 30 alpacas. Animals were 18 months old and were distributed in three groups: without treatment (T0), treated with testosterone enanthate (0.5 mg/kg/body weight) every two weeks during 12 weeks (T2E), and castrated (C0). The penis-prepucial adherences, testicular size, sexual behavior and body weight were evaluated every two weeks, and fibre samples were collected at the beginning of the study and at the end of three experimental periods. In a 4-week pre-experimental phase all parameters were evaluated for adjusting the data of the experimental phase. Release of penis-prepucial adherences was greater in T2E than in T0 and C0 between weeks 8 (p<0.001) and week 12 (0.006>p<0.001). Higher testicular size (length, width, and thickness) occurred in T0 than in T2E between weeks 4and 12 (0.01>p<0.0001). Fibre diameter was similar in T2E and T0, but were greater than in C0 (p<0.05). Clean and greasy fibre weights were similar in all groups. The clean fibre yield was less in T2E than T0 and C0 in the periods II and III (0.04>p<0.0001). Body weight was greater in T2E than in T0 and C0 (p<0.05). It was concluded that the administration of testosterone to young male alpacas was effective in releasing penis-prepucial adherences and in body weight increase, but decreased the testicular size and clean fibre yield; however, did not affect the clean and greasy fibre weights, and fibre diameter.
Key words: alpaca, testosterone, penis-prepucial adherences, fibre
INTRODUCCIÓN
El inicio de la actividad sexual a una edad tempranaresultaimportanteen programasdemejoramiento genético (Cardellino y Rovira, 1995). La maduración sexual de la alpaca macho es usualmente tardío, y en muchoscasos, alos3 años(Fernández-Baca, 1971; Sumar, 1983),completándosecuando las adherencias pene-prepuciales desaparecen(Huanca,2000).Al respecto,sereporta queocurreenel 12%apartirdel añodeedad, en el 60-70% a los 2 años y en 96-100% de los animales a los 3 años (Sumar, 1983; Salas, 2003). Si bien algunos machos pueden presentarlibidosexual apartirdel añodeedad (Fernández-Baca, 1971; Sumar, 1983), sólo se puede emplear en el empadre a machos de dos años que hayan liberado el pene (Núñez et al., 1996; Huanca, 2000). Se ha demostrado que la desaparición de las adherencias del pene depende directamente de los niveles de testosterona; así mismo, tiene relación positiva con el peso corporal y volumen testicular (Chuna et al., 1999). Con esta base, San Miguel et al. (2002) administraron testosterona, pero solo hubo respuesta en el 57% de los machos al año de edad; por lo que se hace necesario obtener un protocolo más eficiente, a fin de maximizar tempranamente el uso de reproductores élite.
A pesar del efecto positivo de la testosterona endógena sobre la madurez sexual (separación pene-prepucio) (Mc Donald, 1991), y desarrollo corporal (anabólico) de las alpacas (García, 1995), se ha demostrado que afecta la calidad de la lana en ovejas (Kiley, 1976; Solís, 1991); asimismo, la terapia con testosterona en ovinos ocasiona un engrosamiento de las fibras aunque sin afectar su longitud (Southcott y Royal, 1971). No se conoce con exactitud el rol de la testosterona sobre el crecimiento de la fibra en alpacas, aunque hay indicios que demuestran que el incremento de los niveles séricos de testosterona afecta el diámetro de fibra y la longitud de mecha (Carpio et al., 1999; Pinazo, 2000; Gardiner, 2005). Ante esto, el presente estudio buscó estudiar el rol biológico de la testosterona sobre la produc-ción y calidad de fibra, a fin de adoptar o corregir medidas que lleven a una producción eficiente de la fibra.
MATERIALES Y MÉTODOS
Lugar de Estudio y Animales
La fase de campo del estudio se realizó en el Centro de Investigación CICAS -La Raya, de la Universidad Nacional de San Antonio Abad del Cusco (UNSAAC), ubicado a 4200 msnm. Las muestras de fibra se analizaron en la Estación Experimental IVITA-Maranganí de la Universidad Nacional Mayor de San Marcos.
Se seleccionaron, al azar, 30 alpacas machos de 18 meses de edad, de la raza Huacaya y de color blanco. Todos presentaban los testículos descendidos y una completa adhesión pene prepucial y ausencia de libido. Los animales eran de un mismo rebaño y estuvieron sometidos a un mismo manejo y alimentación en pastura natural.
Procedimiento Experimental
Se utilizó un diseño completamente randomizado con tres grupos experimentales de 10 animales cada uno: T2E = machos enteros más testosterona, T0 = machos enteros, y C0 = castrados. Este diseño tuvo un doble control, en donde el T0 evaluó el efecto de la testosterona exógena del T2E, y el C0 el efecto de la testosterona endógena del T0. Los machos del C0 se castraron un mes antes de iniciarse el estudio.
El estudio tuvo una fase pre-experimental de cuatro semanas, donde se esquiló un cuadrante de 10 x 10 cm al inicio y al final de cada periodo para evaluar la producción de fibra. Además, se evaluó las adherencias pene-prepuciales, tamaño testicular, líbido sexual y peso corporal.
La fase experimental constó de tres periodos de cuatro semanas cada uno. Se admi nistró enantato de testosterona (0.5 mg/kg, vía i.m., diluida en 0.5 ml de complejo vitamínico liposolubleAD3E) al grupo T2E cada dos semanas; así mismo, se evaluaron las adherencias pene-prepuciales, tamaño testicular, líbido sexual y peso corporal cada dos semanas. La esquila del cuadrante se practicó al final de cada periodo. Los animales de los grupos T0 y C0 solo recibieron el complejo vitamínicoAD3E, como placebo, en cantidad y forma similar al T2E.
Métodos de Evaluación
Determinación del grado de adherencia pene-prepucial
Las alpacas se sujetaron en posición de cúbito lateral. El pene se expuso mediante la extensión de la flexura sigmoidea y retracción prepucial. Los grados de adherencia (Sumar, 1991) fueron: 0) Completa adherencia pene-prepucial; 1) El proceso uretral se encuentra libre; 2) Liberación parcial del pene; 3) Completa liberación peniana.
Comportamiento sexual
El comportamiento sexual se evaluó en un corral a campo abierto de 64 m2. Se enfrentó tres machos con una hembra receptiva por espacio de 5 minutos. Se usó una escala del 1 al 4 (San Miguel et al., 2002): 1) No muestra libido; 2) Libido tenue, intento de monta; 3) Libido con monta, pero sin cópula; y 4) Libido con monta y cópula. En este último caso, en los casos donde pudo haber más de un macho con libido manifiesta, el primer macho fue retirado inmediatamente después de confirmarse la cópula efectiva, permitiendo la evaluación del siguiente macho, y así sucesivamente.
Tamaño testicular
Se midió con un calibrador verniercalliper de 0.5 mm de precisión. Se registró el largo (caudo-craneal), ancho (dorso-ventral), y grosor (latero-lateral) de cada testículo. Cada medida se hizo por duplicado, considerándose el promedio como el valor representativo (San Miguel et al., 2002).
Técnica del cuadrante
Se determinó la producción y calidad de fibra por unidad de superficie y tiempo, a través de un área de 10x 10 cmenla parte media del costillar derecho y a 20 cm de la columna vertebral. Se usó un rasurador eléctrico provisto de un peine Nº 50 y de tamaño «000». El cuadrante fue rasurado al ras de la piel (Braga y Leyva, 1996).
Peso de fibra sucia y limpia (g/cm2)
La fibra se pesó en una balanza electrónica de 1 mg de precisión, previo acondicionamiento en campanas conteniendo sílica gel por 24 horas. El lavado de la fibra sucia se hizo en el extractor Soxhlet con éter de petróleo, colocando las muestras en cartuchos «Thimbel» por 30 minutos. Las muestras se enjuagaron con agua corriente por 5 minutos y desecadas en una estufa a 80 ºC por 3 horas.
Rendimiento de fibra limpia (%)
Se determinó con la fórmula usada por Braga y Leyva (1996): R= (Peso de fibra limpia/Peso de fibra sucia) x 100.
Determinación del diámetro de fibra (µm)
Se usó la fibra limpia y la técnica de proyección microscópica, de acuerdo a la norma descrita por la ASTM D2130-90 (2001). La técnica consiste en cortar las fibras con un micrótomo en segmentos con una altura de 0.6 mm; luego, las fibras se montan en un portaobjetos con bálsamo de Canadá y cubreobjetos, y el diámetro se mide con un microscopio de proyección con un objetivo de 20x (500 aumentos), llegándose a realizar 100 observaciones por muestra (Braga y Leyva, 1996).
Peso corporal
Se usó una balanza tipo reloj (150 kg de capacidad y 0.5 kg de sensibilidad), con la ayuda de un trípode.
Análisis Estadístico
La ocurrencia de liberación de las adherencias peneprepuciales y de libido sexual fueron expresadas en forma porcentual, y la diferencia entre grupos fue determinando con la prueba Chi cuadrado. Los datos promedios de tamaño testicular (promedio de los dos testículos), peso de fibra sucia y limpia, diámetro de fibra, rendimiento de fibra limpia y peso corporal fueron evaluados con el análisis por covarianzas, después que los datos fueron corregidos por la misma prueba, usando los datos de la fase pre-experimental como covariables de corrección. El análisis de diferencia entre grupos se hizo con la prueba de cuadrados mínimos cuando la interacción fue significativa, y en caso contrario se uso la prueba de Tukey protegida. Los datos se analizaron utilizando el paquete estadístico SAS (SAS Institute, 1990).
Los modelos lineales utilizados para el análisis de covarianza fueron Y1 = Tto+Sem +Tto*Sem y Y2 = Tto+Per+Tto*Per, donde Y1= tamaño testicular (largo, ancho y grosor), peso corporal, Y2= peso de fibra sucia y limpia, diámetro de fibra y rendimiento de fibra limpia, Tto = tratamiento, Sem = semana, y Per = periodo.
RESULTADOS
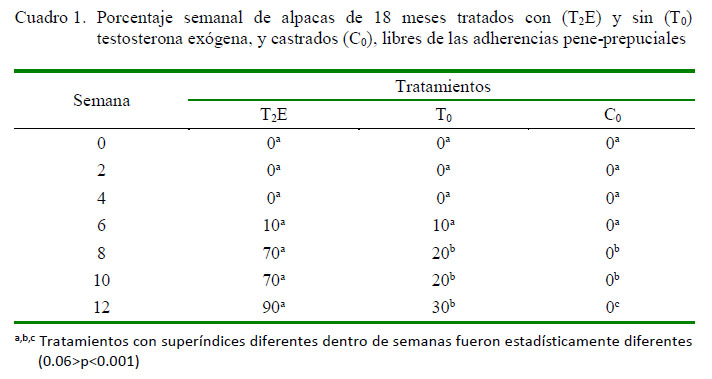
La liberación completa de las adherencias pene-prepuciales se observó desde la semana 6 en algunos animales tratados con (T2E) y sin (T0) testosterona exógena. El número de animales sin adherencias prepuciales se incrementó significativamente en los T2E de la semana 8 (p<0.001) a la semana 12 (p<0.006), en relación a los T0. En forma similar, se observó diferencias significativas entre los T0 y los C0 (p<0.06) en la semana 12 (Cuadro 1 ).
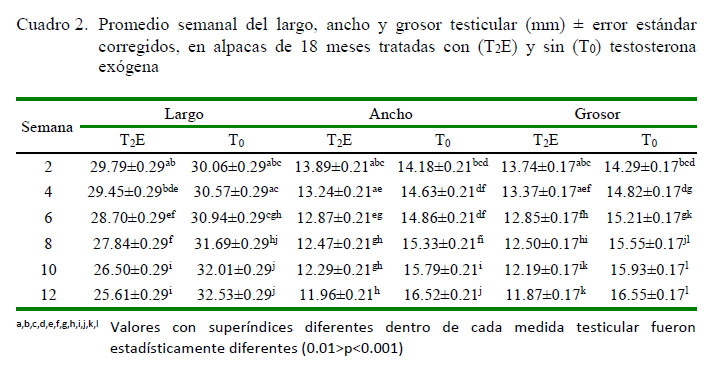
El efecto de las variables tratamiento y semana fueron dependientes, a través de la interacción significativa (p<0.0001) tratamiento*semana en el largo, ancho y grosor de los testículos, donde el incremento en el tamaño, en sus tres dimensiones, ocurrió a partir de la cuarta semana en adelante en las alpacas sin tratamiento hormonal (T0), a diferencia de los que recibieron testosterona (T2E) (0.01>p<0.0001) (Cuadro 2 ).
En el peso de fibra sucia y fibra limpia, el efecto interactivo tratamiento*periodo y el de la variable tratamiento estuvo ausente; sin embargo, hubo efecto simple de la variable periodo (p<0.0001). El peso de fibra sucia se incrementó constante y significativamente (p<0.05) cada 4 semanas, siendo 1.51 ± 0.05, 1.71 ± 0.05 y 1.94 ± 0.05 g/cm2en los periodos I, II y III, respectivamente. En forma similar ocurrió con el peso de fibra limpia (p<0.05), siendo de 1.27 ± 0.04, 1.46 ± 0.04 y 1.68 ± 0.04 g/cm2 en los periodos I, II y III, respectivamente. Los pesos de fibra sucia y limpiadelosanimalesdelos gruposT2E,T0y C0 fueron similares.
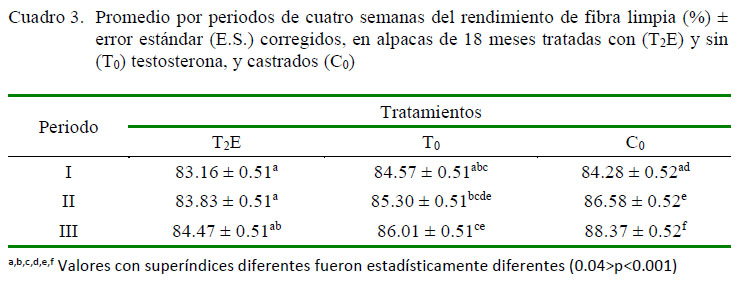
El rendimiento de fibra limpia fue afectado por la interacción tratamiento*periodo (p<0.05), donde las alpacas tratadas con testosterona (T2E) tuvieron menor rendimiento que las alpacas sin tratamiento (T0) y castradas (C0) en los periodos II y III (0.04>p<0.0001); así mismo, el rendimiento de fibra limpia fue mayor en los T0 que en los C0 en el periodo III (p<0.001) (Cuadro 3 ).
El efecto interactivo tratamiento* periodo estuvo ausente en el diámetro de fibra; sin embargo, hubo efecto simple (p<0.0001) de las variables periodo y tratamiento. Independiente del tratamiento, el diámetro de fibra se incrementó constantemente, cada 4 semanas, entre los periodos I (18.23 ± 0.15 µm) y II (18.92 ± 0.15 µm), y marcadamente en el periodo III (19.90 ± 0.15 µm, p<0.05). Por otro lado, independiente del periodo, los animales tratados (T2E) y no tratados (T0) con testosterona tuvieron diámetros de fibra si-milares,19.56 ± 0.15y19.59± 0.15µm,respectivamente, pero ambos fueron mayores (p<0.05) que los animales castrados (C0) (17.90 ± 0.15 µm).
El efecto interactivo tratamiento* periodo estuvo ausente en el peso corporal; sin embargo, hubo efecto simple (p<0.0001) de las variables periodo y tratamiento. Independiente del tratamiento, el incremento del peso corporal fue significativo (p<0.05) a partir de la sexta semana; e independiente del periodo, los animales tratados con testosterona (T2E) fueron más pesados (39.80 ± 0.15 kg, p<0.05) que los animales sin tratamiento (T0) y castrados (C0), que tuvieron pesos similares (38.81 ± 0.15 y 37.94 ± 0.15 kg, respectivamente).
DISCUSIÓN
La administración de testosterona durante 12 semanas a alpacas de 18 meses de edad, con completa adhesión pene-prepucial, ocasionó la liberación del pene en el 90% de los animales, resultado superior al obtenido por San Miguel et al. (2002) empleando 109 mg quincenales y por 10 semanas. Los resultados demostraron que la separación pene-prepucial es testosterona dependiente, lo cual había sido descrito en alpacas (Chuna et al., 1999), bovinos (Lunstra et al., 1978) y ovinos (Pattie et al., 1964). Asimis-mo, el efecto de la testosterona fue reforzado por el efecto de la castración, que arrestó la liberación pene-prepucial en todos los machos, resultados que fueron similares a los encontrados en otras especies (Korenbrot et al., 1977; Mather, 1980; Bloch y Mills, 1995).
La administración de testosterona exógena redujo el incremento del tamaño testicular, similar al efecto encontrado en alpacas de un año de edad (San Miguel et al., 2002), y en bovinos (Godfrey et al., 1992), y hámsters (Pak et al., 2002) prepúberes, como resultado de la supresión de la espermatogénesis (Freitag y Docke, 1987; Pak et al., 2002). Debe haber ocurrido un efecto inhibitorio de la testosterona sobre la secreción de la GnRH y gonadotropinas (Godfrey et al., 1992; Dewan et al., 2000), ya que el crecimiento testicular depende de la secreción de las gonadotropinas en ovinos (Brown et al., 1994; Ulker et al., 2005) y bovinos (Huxsoll et al., 1998). Se asume, además, que el sistema aromataza de las regiones hipotalámicas que controlan la secreción negativa de GnRH en las alpacas se encuentra funcional entre los 18 y 21 meses de edad, puesto que la testosterona ejerce su acción negativa sobre la GnRH solamente tras su aromatización a estrógenos en ovinos y bovinos (Schanbacher, 1982).
El inicio de la libido sexual es uno de los indicadores del logro de la pubertad en el macho, la cual depende de la actividad testicular para secretar niveles apropiados de testosterona (Hafez, 1989), que estimulen los centros sexuales cerebrales, a través de una activación previa de los receptores androgénicos y del sistema enzimático aromatasa (Vagell y McGinnis, 1997). Es posible que en el presente estudio no se activaron los receptores androgénicos o del sistema aromatasa para la metabolización de la testosterona. Por otro lado, la castración ocasionó la esperada ausencia del desarrollo del comportamiento sexual, aunque con una terapia con testosterona a niveles fisiológicos del animal adulto, se ha demostrado en ovinos y ratas que se favorece el inicio y mantenimiento de la libido sexual (Clegg et al., 1969; Bloch y Mills, 1995).
La similitud en el diámetro de fibra entre alpacas tratadas con y sin testosterona, sugiere ausencia del efecto aditivo de esta hormona, probablemente debido a que los animales no han alcanzado una edad suficiente para obtener una respuesta del folículo piloso o porque el tratamiento hormonal fue insuficiente. En la rata se logró obtener un incremento del diámetro de fibra por efecto de la testosterona (Paus y Cotsarelis, 1999) y en ovinos se pudo observar una tendencia al incremento del diámetro de fibra con dosis crecientes (Osborne y Widdows, 1961; Southcott y Royal, 1971), aunque no se encontró efecto al realizar una con hemicastrados o con criptorquídeos inducidos (Foster et al., 1997). Sin embargo, no se descarta el efecto de la testosterona, toda vez que el diámetro de la fibra fue significativamente menor en los animales castrados. Esto se corrobora por el estudio de Gardiner (2005), aunque hay otros estudios donde no hubo diferencias (Rodríguez et al., 1986).
La fibra sucia, una vez eliminada las impurezas del vellón (materiales extraños y secreciones de las glándulas sebáceas y sudoríparas) se convierte en fibra limpia, de allí que el mayor rendimiento de fibra limpia dependerá del menor grado de impurezas en la fibra sucia (Ruiz de Castilla, 1994). En el presente estudio, el rendimiento de fibra limpia en los animales tratados con testosterona fue inferior debido al estímulo que ejerce sobre la actividad secretoria de las glándulas sebáceas (Ebling et al., 1969), cuyas secreciones facilitan la mayor captación de impurezas (Ryder, 1973).Asimismo, el mejor rendimiento de fibra limpia en los castrados se pudo haber debido a la disminución de la actividad secretoria de las glándulas sebáceas (Kiley, 1976; Iwata et al., 2000).
Las alpacas de 1.5 años de edad, tratadas con testosterona, alcanzaron mayores pesos vivos que los no tratadas, en forma similar a otrosestudios(San Miguel etal.,2002) en alpacas de un año de edad; sin embargo, las discrepancias con otros resultados concuerdan con lo indicado por Hansel y McEntee (1981), que el efecto anabólico depende de la dosis, especie y estado nutritivo del animal, o del nivel de testosterona endógena (San Miguel et al., 2002). Por otro lado, los pesos vivos similares entre machos enteros sin tratamiento y machos castrados indican que, entre los 18 y 21 meses de edad, la influencia de la testosterona endógena sobre el crecimiento corporal es inexistente (Pereira et al., 2002; De Azambuja et al., 2003).
CONCLUSIONES
-
La liberación de las adherencias peneprepuciales en la alpaca se estimula por la testosterona exógena y se suprime por la castración.
-
El incremento del tamaño testicular se reduce por la testosterona exógena.
-
El diámetro de fibra de los machos tratados y no tratados con testosterona fue similar, pero mayor que en los castrados.
-
Los pesos de fibra sucia y limpia no se alteraron por la administración de testosterona ni por la castración.
-
El rendimiento de fibra limpia se redujo por la administración de testosterona; sin embargo, hubo un mayor rendimiento en los castrados.
-
El peso corporal de los machos tratados con testosterona fue mayor que en machos no tratados y castrados.
Agradecimiento
Los autores agradecen a INCAGRO por el financiamiento del presente estudio.
LITERATURA CITADA
1. Adams NR, Cronjé PB. 2003. A review of the biology linking fibre diameter with fleece weight, liveweight, and reproduction in merino sheep. Aus J Agric Res 54: 1-10. doi: 10.1071/ AR02059 [ Links ]
2. ASTM D2130-90. 2001. Standard method for studies of diameter in wool and others animal fibers by microprojection. West Conshohocken. PA, USA: American Society for Testing and Materials. 11 p. [ Links ]
3. Bloch GJ, Mills R. 1995. Prepubertal testosterone treatment of neonatally gonadectomized male rats: defeminization and masculinization of behavioral and endocrine function in adulthood. Neurosci Biobehav Rev 19: 187-200. doi: 10.1016/0149-7634(94)-00064-8 [ Links ]
4. Braga W, Leyva V. 1996. Efecto de la altitud sobre la producción de fibra de alpaca. En: XIX Reunión APPA. Cusco: Asociación Peruana de Producción Animal. [ Links ]
5. Brown BW, Mattner PE, Carroll PA, Holland EJ, Paull DR, Hoskinson RM, Rigby RD. 1994. Immunization of sheep against GnRH early in life: effects on reproductive function and hormones in rams. J Reprod Fertil 101: 15-21. [ Links ]
6. Cardellino R, Rovira J. 1995. Mejoramiento genético animal. Uruguay: Hemisferio Sur. 253 p. [ Links ]
7. Carpio M, Ordóñez C, Alarcón V, Bravo PW. 1999. Presencia de espermatozoides, niveles de testosterona, y tamaño testicular en alpacas. En: II Congreso CMCS. Cusco: Congreso Mundial sobre Camélidos Sudamericanos. [ Links ]
8. Chuna P, Leyva V, Franco J. 1999. Adherencias pene-prepuciales y niveles de testosterona circulantes en alpacas. Rev Inv Vet Perú 10(2):11-16. [ Links ]
9. Clegg MT, Beamera W, Bermant G. 1969. Copulatory behaviour of the ram, Ovis aries. III: Effects of pre- and postpubertal castration and androgen replacement therapy. Anim Behav 17: 712-717. doi: 10.1016/S0003-3472(69)- 80017-5 [ Links ]
10. De Azambuja EL, Das Dores Ferreira L, Da Rocha MA, Yurika I. 2003. Desempenho de cordeiros inteiros ou submetidos a diferentes métodos de castração abatidos aos 30 kg de peso vivo. Rev Bras Zootec 32: 745-752. doi: 10.1590/S1516-35982003000300027 [ Links ]
11. Dewan ZF, Morris ID, Lendon RG. 2000. Administration of exogenous testosterone in the adult rat and its effects on reproductive organs, sex hormones and body-weight. Bangladesh Med Res Counc Bull 26(2): 48-55. [ Links ]
12. Ebling FJ, Ebling E, Skinner J. 1969. Influence of pituitary hormones on the response of the sebaceous glands of the male rat to testosterone. J Endocrinol 45: 245-56. doi: 10.1677/joe.0.0450245 [ Links ]
13. Fernández-Baca S. 1971. La alpaca: Reproducción y cCrianza. Bol Div IVITA-Univ. Nac. Mayor de San Marcos 7: 14-29.
14. Foster FM, Jackson RB, Hopkins DL, Corkrey R. 1997. Production and management considerations of running wethers, hemi-castrates and induced cryptorchids for wool production. Aus J Exp Agric 37: 303-310. [ Links ]
15. Freitag J, Docke F. 1987. Differential effects of chronic testosterone treatment on the onset of puberty in male rats. Exp Clin Endocrinol 90: 361-364. doi: 10.1055/ s-0029-1210712 [ Links ]
16. García A. 1995. Fisiología veterinaria. España: McGraw Hill Interamericana. 1074 p. [ Links ]
17. Gardiner C. 2005. «What is the value of a gelding alpaca». Alpaca Library. [Internet]. Disponible en: http:// www.alpacas.com/alpacalibrary [ Links ]
18. Godfrey RW, Lunstra DD, Schanbacher BD. 1992. Effect of implanting bull calves with testosterone propionate, dihydrotestosterone propionate or oestradiol-17 beta prepubertally on the pituitary-testicular axis and on postpubertal social and sexual behaviour. J Reprod Fertil 94: 57-69. [ Links ]
19. Hafez ESE. 1989. Reproducción e inseminación artificial en animales. 5ta ed. México: Interamericana McGraw-Hill. 694 p. [ Links ]
20. Hansel W, McEntee K. 1981. Procesos reproductores masculinos. En: Dukes HH, Swenson MJ (eds). Fisiología de los animales domésticos. Tomo II. México: Aguilar. p 1649-1701. [ Links ]
21. Huanca T. 2000. Evaluación de la pubertad en alpacas machos. Tesis de Magíster. Lima: Facultad de Medicina Veterinaria, Univ Nacional Mayor de San Marcos. 66 p. [ Links ]
22. Huxsoll CC, Price EO, Adams TE. 1998. Testis function, carcass traits, and aggressive behavior of beef bulls actively immunized against gonadotropinreleasing hormone. J Anim Sci 76: 1760- 1766. [ Links ]
23. Iwata E, Wakabayashi Y, Kakuma Y, Kikusui T, Takeuchi Y, Mori Y. 2000. Testosterone-dependent primer pheromone production in the sebaceous gland of male goat. Biol Reprod 62: 806-810. [ Links ]
24. Kiley M. 1976. A review of the advantages and disadvantages of castrating farm livestock with particular reference to behavioural effects. Br Vet J 132: 323-331. [ Links ]
25. Korenbrot CC, Huhtaniemi IT, Weiner RI. 1977. Preputial separation as an external sign of pubertal: development in the male rat. Biol Reprod 17: 298-303. doi: 10.1095/ biolreprod17.2.298 [ Links ]
26. Lunstra DD, Ford JJ, Echternkamp SE. 1978. Puberty in beef bulls: hormone concentrations, growth, testicular development, sperm production and sexual aggressiveness in bulls if different breeds. J Anim Sci 46: 1054-1062. [ Links ]
27. Mather EC. 1980. Puberty in the bull. In: Morrow D (ed). Current therapy in theriogenology. Diagnosis, treatment, and prevention of reproductive diseases in animals. Philadelphia: WB Saunders. p 339-342. [ Links ]
28. Mc Donald LE. 1991. Endocrinología veterinaria y reproducción. 4ta ed. México: Interamericana McGraw-Hill. 551 p. [ Links ]
29. Nuñez R, Olarte U, Olivera L. 1996. Relación de la espermatogénesis y desprendimiento de la adherencia peneprepucial de la alpaca (Lama pacos). ALLPAK’A Rev Inv Camélidos Sudamericanos 5(1): 74-87.
30. Osborne WB, Widdows FA. 1961. Sheath rot and testosterone - Its effects and side effects on health and production. Wool Tech Sheep Breed 8: 99. [ Links ]
31. Pak TR, Lynch GR, Tsai PS. 2002. Differential alteration of the reproductive axis by testosterone and estrogen in peripubertal and adult male Siberian hamsters (Phodopus sungorus). Biol Reprod 67: 706-711. doi: 10.1095/ biolreprod.102.003434 [ Links ]
32. Pattie WA, Godlee AC, Bouton PE. 1964. The effects of castration and of the poll gene on prime lamb production. Aus J Exp Agric Anim Husb 4(15): 386- 391. [ Links ]
33. Paus R, Cotsarelis G. 1999. The biology of hair follicles. New Engl J Med 341: 491-497. doi: 10.1056/NEJM19990- 8123410706 [ Links ]
34. Pereira PHS, Osório JCS, Osório MTM, Oliveira NM, Faria HV, Pimentel MA. 2002. Componentes do peso vivo em cordeiros castrados e não castrados. Rev Bras Agrociência 8(1): 57-60. doi: 10.18539/cast.v8i1.429 [ Links ]
35. Pinazo RA. 2000. Algunas características físicas de la fibra de alpaca huacaya y suri en el CIP La Raya-UNA-Puno. Tesis de Médico Veterinario y Zootecnista. Puno: Univ Nacional del Antiplano. 78 p. [ Links ]
36. Rodríguez H, Quintana F, Gamarra M, Manrique G. 1986. Efectos del sexo, color y variedad sobre parámetros productivos en alpacas tuis. En: IX Reunión APPA. Tingo María: Asociación Peruana de Producción Animal. [ Links ]
37. Ruiz de Castilla M. 1994. Camelicultura: alpacas y llamas del sur del Perú. Cusco: Mercantil. 180 p. [ Links ]
38. Ryder ML. 1973. Hair. In: Edward A (ed). Studies in Biology N°41. London: Camelot Press. 58 p. [ Links ]
39. Salas WH. 2003. Variabilidad en la permanencia de la adherencia peneprepucial en la alpaca huacaya. Tesis de Magíster. Lima: Univ Nacional Agraria la Molina. 50 p. [ Links ]
40. San Miguel C, Leyva V, García W. 2002. Administración de testosterona en alpacas con adherencias pene-prepuciales. Rev Inv Vet Perú 13(2): 28-36. doi: 10.15381/rivep.v13i2.7327 [ Links ]
41. SAS Institute. 1990. User’s guide, version 6, 4th ed. Vol 1. Cary, North Caroline: SAS Institute Inc. 943 p.
42. Schanbacher BD. 1982. Hormonal interrelationships between hypothalamus, pituitary and testis of rams and bulls. J Anim Sci 55(Suppl 2): 56-67. [ Links ]
43. Solís R. 1991. Tecnologías de lana y fibras de animales especiales. Revisión. Cerro de Pasco: Univ Nacional Daniel Alcides Carrión. 596 p. [ Links ]
44. Southcott WH, Royal WM. 1971. Effect of implanted testosterone propionate on the growth and wool production of merino wethers. Aust J Agric Res 22: 271-282. [ Links ]
45. Sumar J. 1983. Studies in reproductive pathology in alpacas. MSc Thesis. Uppsala: Swedish Univ Agric Sci 90 p. [ Links ]
46. Sumar J. 1991. Fisiología de la reproducción del macho y manejo reproductivo. En: Fernández-Baca S (ed). Avances y perspectivas del conocimiento de los camélidos sudamericanos. Chile: Oficina Regional de la FAO para América Latina y el Caribe. p 111- 125. [ Links ]
47. Ulker H, Kanter M, Gokdal O, Aygun T, Karakus F, Sakarya ME, DeAvila DM, Reeves JJ. 2005. Testicular development, ultrasonographic and histological appearance of the testis in ram lambs immunized against recombinant LHRH fusion proteins. Anim Reprod Sci 86: 205-219. doi: 10.1016/j.anireprosci.2004.07.011 [ Links ]
48. Vagell ME, McGinnis MY. 1997. The role of aromatization in the restoration of male rat reproductive behavior. J Neuroendocrinol 9: 415-421. [ Links ]
Recibido: 15 de agosto de 2016
Aceptado para publicación: 7 de diciembre de 2016
