










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versión impresa ISSN 1609-9117
Rev. investig. vet. Perú vol.29 no.1 Lima ene./mar. 2018
http://dx.doi.org/10.15381/rivep.v29i1.14253
ARTÍCULOS PRIMARIOS
Actividad antioxidante del isoespintanol y el timol en el semen equino criopreservado
Antioxidant activity of isoespintanol and thymol in cryopreserved stallion semen
Giovanni Restrepo B.1,3, Benjamín A. Rojano2
1 Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Medellín, Colombia
2 Facultad de Ciencias, Universidad Nacional de Colombia, Medellín, Colombia
3 E-mail: grestre0@unal.edu.co
RESUMEN
El isoespintanol proveniente de las hojas de Oxandra cf xylopioides y el timol presente en los aceites esenciales del tomillo (Thymus vulgaris L) y el orégano (Origanum vulgare L) son conocidos por su actividad antioxidante en diferentes medios. En este estudio se evaluó la actividad antioxidante del isoespintanol y el timol en el semen equino criopreservado. Se colectó el semen de cinco caballos criollos colombianos y se sometió a un protocolo de congelación, en un diluyente suplementado con isoespintanol o timol. Se evaluó la producción de especies reactivas de oxígeno y la peroxidación lipídica del semen descongelado. Como resultado, se observó una reducción pos-descongelación de la producción de especies reactivas de oxígeno del semen por efecto de ambos antioxidantes, mientras no se encontró diferencia entre los niveles de peroxidación lipídica del semen pos-descongelado. El isoespintanol y timol actúan como antioxidantes en el semen equino sometido a la criopreservación.
Palabras clave: antioxidante; congelación; especies reactivas de oxígeno; peroxidación lipídica
ABSTRACT
Isoespintanol from the leaves of Oxandra cf xylopioides and thymol present in the essential oils of thyme (Thymus vulgaris L) and oregano (Origanum vulgare L) is known for their activity as antioxidants in different media. In this study, the antioxidant activity of isoespintanol and thymol in cryopreserved stallion semen was evaluated. The semen of five Colombian Creole horses was collected and subjected to a freezing protocol, in an extender supplemented with isoespintanol or thymol. The production of reactive oxygen species and the lipid peroxidation of the thawed semen were evaluated. As a result, a post-thawing reduction of the production of reactive oxygen species of semen was observed due to the effect of both antioxidants, whereas no difference was found in the lipid peroxidation levels of post-thawed semen. Isoespintanol and thymol act as antioxidants in stallion semen subjected to cryopreservation.
Key words: antioxidant; freezing; reactive oxygen species; lipid peroxidation
INTRODUCCIÓN
La criopreservación de semen es uno de los procedimientos más importantes en el desarrollo de las biotecnologías para la reproducción asistida en equinos. Sin embargo, el semen equino es extremadamente sensible a las alteraciones celulares generadas por la congelación (Ball, 2008). Uno de los factores más relacionados con la baja fertilidad del semen equino criopreservado es el estrés oxidativo (Ball y Baumber, 2001). Se conoce que los procesos de congelación producen un incremento en la generación de especies reactivas de oxígeno (ERO), debido al des-acoplamiento del metabolismo oxidativo normal de los espermatozoides (Ortega-Ferrusola et al., 2009).
De igual forma, se sugiere que los procesos de congelación conducen a la pérdida de la actividad de defensa antioxidante, principalmente a causa de la alteración de las enzimas antioxidantes (Isachenko et al., 2004). La remoción del plasma seminal es otro proceso que favorece el estrés oxidativo del semen equino, toda vez que allí se encuentra la mayor parte de su capacidad antioxidante (Ball, 2008; Wnuk et al., 2010). Otros fenómenos que contribuyen al incremento de las ERO son: el aumento en el número de células apoptóticas (Ortega-Ferrusola et al., 2009), la presencia de leucocitos y espermatozoides morfológicamente anormales (Baumber et al., 2000) y el estrés osmótico, el cual ha sido asociado con el incremento del peróxido de hidrógeno (H2O2) (Burnaugh et al., 2007), conocido como la especie reactiva más involucrada en el daño de los espermatozoides equinos (Baumber et al., 2000). El estrés oxidativo desencadena la peroxidación lipídica, un proceso complejo que altera los ácidos grasos poliinsaturados presentes en alta proporción en la membrana de los espermatozoides equinos (Macias-Garcia et al., 2011) y que ocasiona una reducción de la movilidad, la vitalidad y la capacidad de dichas células para la fertilización (Baumber et al., 2002).
De acuerdo con lo anterior, los antioxidantes se han convertido en una valiosa alternativaconel findereducirel estrésoxidativo en el semen equino criopreservado (Barros et al., 2012; Cocchia et al., 2011; Lisboa et al., 2012). Sin embargo, una variedad de moléculas antioxidantes suplementadas en el diluyente para la criopreservación de semen equino no ha mostrado una mejora sustancial de la calidad seminal (Hussain et al., 2011; Martins et al., 2012; Vasconcelos et al., 2013; Martins et al., 2015). No obstante, en estudios recientes se han identificado algunas sustancias con efectos benéficos sobre los espermatozoides criopreservados (Coyan et al., 2011; Gibb et al., 2013; Stephens et al., 2013), reportándose en algunos casos, efectos deletéreos como el bloqueo meiótico de los ovocitos (Blanes-Zamora et al., 2008).
En otros sistemas biológicos se conoce la considerable capacidad antioxidante de diferentes moléculas de origen natural. Entre dichas moléculas está el timol (2-isopropil-5metilfenol), presente en la naturaleza en aceites esenciales de Thymus vulgaris L. (Tomillo) (fam. Lamiaceae) y Origanum vulgare L. (orégano) (fam. Lamiaceae). Esta molécula es conocida por una gran capacidad para inhibir la peroxidación lipídica en diferentes sistemas (Rojano et al., 2008c), al igual que por su alta actividad anti-radical (Lagouri y Boskou, 1995). El isoespintanol (2-isopropil3,6-dimetoxi-5-metilfenol) es un análogo biosintético del timol, extraído de las hojas de Oxandra cf xylopioides Diels (fam. Annonaceae). Es considerado como mejor captador de radicales y mejor reductor que el timol en diferentes medios, siendo catalogado como dos veces mejor antioxidante (Rojano et al., 2008a). El isoespintanol inhibe en alimentos la peroxidación lipídica (Rojano et al., 2008b) y en modelos animales se ha observado su capacidad para disminuir la inflamación (Rojano et al., 2007). Dado lo anterior, el timol, el isoespintanol e incluso otras moléculas isopropil-fenólicas, podrían tener un impacto positivo en la reducción del estrés oxidativo de células reproductivas. El objetivo de esta investigación fue evaluar la actividad antioxidante del isoespintanol y el timol en el semen equino criopreservado.
MATERIALES Y MÉTODOS
Se trabajó con 21 muestras de semen (eyaculados) provenientes de cinco caballos criollos colombianos, las cuales fueron distribuidas en dos etapas. La primera etapa fue la definición de la concentración de isoespintanol y timol a utilizar en el proceso de criopreservación, para lo cual se utilizaron seis muestras seminales. La segunda etapa fue la criopreservación del semen con las concentraciones seleccionadas de los antioxidantes y la evaluación pos-descongelación, para lo cual se destinaron 15 muestras seminales.
Obtención de Muestras Seminales
Los animales del estudio estuvieron ubicados en el norte del Valle de Aburra, departamento de Antioquia (Colombia), latitud 6°23’41.50"N y longitud 75°25’27.95"O, con una altitud promedio de 1475 msnm, una temperatura promedio de 22 °C y una pluviosidad cercana a los 2200 mm/año.
Los animales tenían entre 2 y 8 años y se encontraban en un régimen de mínimo una colecta semanal, con fertilidad comprobada mediante crías nacidas vivas y con una condición corporal de 6 a 7 (escala entre 1 y 9). Fueron sometidos a condiciones similares de manejo en estabulación, con alimentación a base de pasto estrella henificado (Cynodon nlemfuensis) suministrado a voluntad. El semen se colectó mediante el método de la vagina artificial, con una vagina modelo Missouri (Minitube, Tiefenbach, Alemania), lubricada con gel no espermicida y sobre una yegua. La fracción en gel del eyaculado fue removida por filtración.
Se avaluó la calidad del semen a través de la medición del volumen, la concentración de espermatozoides por espectrofotometría (Spermacue®, Minitube, Tiefenbach, Alemania), la movilidad espermática por microscopía (Eclipse E200®, Nikon Inc., Mellville, EEUU) y la morfología y vitalidad de los espermatozoides por la técnica de eosinanigrosina modificada (Gamboa y Ramalho-Santos, 2005; Brito et al., 2011). Como criterios de selección solo se procesaron los eyaculados con parámetros mínimos de 70% en la movilidad espermática, concentración de 100x106 espermatozoides/ml y 70% de morfología normal y vitalidad (Pérez-Osorio et al., 2008). El semen se diluyó en proporción 1:1 en un diluyente a base de leche semidescremada, caseinatos de sodio y azúcares, y luego se transportó al laboratorio en condiciones de refrigeración (5 ºC).
Definición de la Concentración Antioxidante
Las muestras de semen fresco fueron suplementadas de forma separada con isoespintanol o timol a las concentraciones de 20, 40, 50, 60, 80 y 100 µM, tomando como referencia estudios de Rojano et al.(2008b,c). La evaluación de la producción de ERO se hizo mediante la metodología de la 2,7-diclorodihidrofluoresceina diacetato (H2DCFDA) (Aitken et al., 2013). Cada muestra se preparó con 30 µl de semen, 240 µl de solución buffer (pH 7.4) y 30 µl de una solución 40 mM de H2DCFDA (Intervet International BV, Boxmeer, Países Bajos). Se emplearon condiciones controladas (37 °C, pH 7.4) y como referencia el antioxidante Trolox® (Merck, Darmstadt, Alemania). Las lecturas para cada eyaculado suplementado con las concentraciones de isoespintanol o timol en el semen fresco se realizaron por cuadriplicado, cada 60 s durante 80 min mediante un espectrofluorímetro LS 55 (Perkin Elmer, Massachusetts, EEUU).
Se calculó el porcentaje de inhibición de la producción de ERO de acuerdo con la expresión: Porcentaje de inhibición = (ms-m c/ mc)x100, donde ms es la pendiente de cada concentración y mc es la pendiente de un control sin antioxidante (Rojano et al., 2008a). Para el proceso de criopreservación seminal se seleccionó la concentración de cada antioxidante con la máxima capacidad de inhibición de la producción de ERO.
Criopreservación del Semen
La criopreservación se realizó mediante un protocolo de congelación modificado (Bustamante et al., 2009). El semen se centrifugó por 15 min a 1200 g, el precipitado se resuspendió en un diluyente de congelación a base de leche semidescremada, caseinatos de sodio y azúcares, suplementado con 4% de yema de huevo y5% de N,N-dimetilformamida (Sigma-Aldrich, St. Louis, EEUU), en cantidad suficiente para una concentración final de 100x106 espermatozoides/ml.
El semen diluido se dividió en tres alícuotas, asignadas aleatoriamente a uno de los tres tratamientos: timol, isoespintanol o control (sin antioxidante). Posteriormente, el semen se mantuvo a 5 °C por 15 min y luego se empacó en pajillas para 0.5 ml. Las pajillas se sometieron a vapores de nitrógeno líquido por 15 min, para finalmente ser almacenadas en un termo con nitrógeno líquido. Después de un periodo de almacenamiento mínimo de dos semanas, las pajillas de semen se descongelaron en agua a 37 °C por 1 min.
Evaluación de la Actividad Antioxidante
La medición de la producción de ERO del semen descongelado se hizo por cuadriplicado, cada 60 s durante 80 min para cada eyaculado y tratamiento, mediante la metodología previamente descrita de la H2DCFDA. Se halló la velocidad de producción de ERO del semen, mediante el cálculo de la pendiente (m = Δy/Δx), a partir de la evaluación de la cinética de producción de ERO (producción de ERO vs. tiempo).
También se realizó la evaluación pos-descongelación de la peroxidación lipídica del semen mediante la metodología modificada de las sustancias reactivas al ácido tiobarbitúrico TBARS (Morte et al., 2008). Muestras de 50 µl de semen se incubaron por 1 h a 37 °C y luego fueron enfriadas en agua con hielo durante 10 min. Posteriormente, se adicionaron 40 µl de ácido tricloroacético (TCA, Sigma-Aldrich, St. Louis, EEUU) al 15% y 80 µl de ácido tiobarbitúrico al 0.67% (TBA, Sigma-Aldrich, St. Louis, EEUU). La mezcla fue agitada e incubada por 20 min a 90 °C y luego enfriada en agua con hielo durante 10 min. Se adicionaron 300 µl de butanol (Sigma-Aldrich, St. Louis, EEUU), para luego agitar cada muestra por inversión durante 1 min. Finalmente se tomaron 300 µl de la capa superior de la muestra, los cuales se sirvieron en una placa multipozo. El complejo fluorescente formado fue leído por cuadriplicado a partir del semen descongelado proveniente de cada eyaculado y tratamiento, mediante en un espectrofluorímetro LS 55 (Perkin Elmer, Massachusetts, EEUU).
Análisis Estadístico
Para evaluar el efecto de los antioxidantes sobre la producción de ERO y la peroxidación lipídica del semen se ajustó un modelo mixto de medidas repetidas en el tiempo y un modelo mixto completamente aleatorizado, respectivamente. En ambos modelos se incluyó el efecto fijo del tratamiento y el efecto aleatorio anidado del eyaculado dentro del equino. Las covariables incluidas en cada modelo, se definieron mediante un análisis de correlación de Pearson. Para evaluar la influencia de los factores tiempo y edad del animal sobre la producción de ERO del semen se realizó un análisis de regresión lineal múltiple.
Dado el uso de pruebas paramétricas se comprobó la normalidad de los datos por la prueba de Shapiro-Wilk. La comparación de las medias entre los tratamientos se realizó mediante la prueba de Tukey. El nivel de significancia considerado para todas las evaluaciones fue p<0.05. Los resultados se expresaron como media ± error estándar. Todos los análisis se realizaron mediante el programa SAS 9.2. (SAS Inst. Inc., Cary, NC).
RESULTADOS
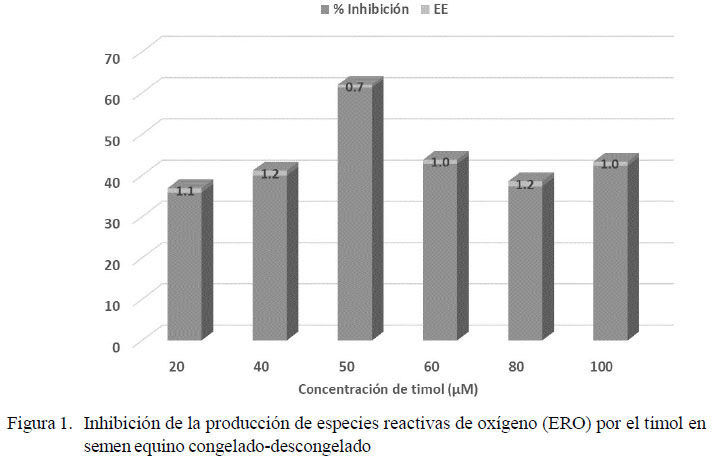
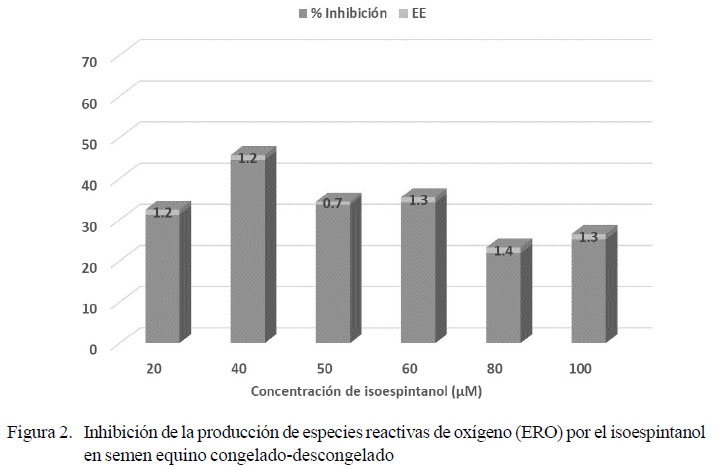
Los resultados para los parámetros de calidad del semen fresco fueron (media ± error estándar): volumen 36.7 ± 5.3 ml; concentración 223.42 ± 26.9 x106 espermatozoides/ml; movilidad espermática 77.5 ± 3.3%; vitalidad 79.9 ± 2.7% y morfología normal 83.6 ± 2.1%. A partir de 2594 registros de producción de ERO en semen fresco suplementado con isoespintanol o timol en seis concentraciones (20, 40, 50, 60, 80, 100 µM), se halló el valor promedio de 24.29 ± 0.34 Unidades Relativas de Fluorescencia (URF). Las concentraciones con mayor inhibición de ERO para cada antioxidante fueron: timol 50 µM con 61.3% ± 0.7 (Figura 1) e isoespintanol 40 µM con 44.5% ± 1.2 (Figura 2). Dichas concentraciones fueron utilizadas para la criopreservación seminal.
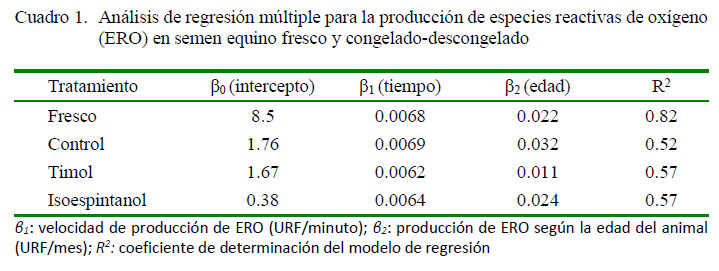
A partir de 2653 registros de producción de ERO de semen fresco y criopreservado se halló un promedio de 15.96 ± 0.23 URF, con un coeficiente de variación de 28.7%. El coeficiente de determinación (R2) del modelo para ERO fue de 0.86. En el modelo mixto ajustado fueron significativos los efectos fijos del tiempo y el tratamiento (p<0.05). Los coeficientes de regresión para la producción de ERO se presentan en el Cuadro 1.
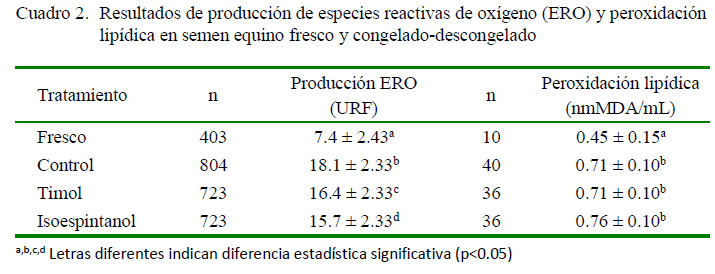
A partir de 122 registros de peroxidación lipídica del semen se encontró un promedio de 0.70 ± 0.02 nmMDA/ml, con un coeficiente de variación de 23.5%. Se halló un coeficiente de correlación entre la peroxidación lipídica y la velocidad de producción de ERO de γ = 0.48. El promedio para la velocidad de producción de ERO fue de 0.0065 ± 0.0002 URF/min. El coeficiente de determinación (R2) del modelo para peroxidación lipídica fue de 0.82. Fue significativo el efecto fijo del tratamiento (p<0.05). Los análisis de comparación de las medias de producción de ERO y peroxidación lipídica bajo los diferentes tratamientos se presentan en el Cuadro 2.
DISCUSIÓN
En la literatura se reportan indicadores de calidad del semen equino que pueden ser equiparables a los hallados en esta investigación (Pérez-Osorio et al., 2008; Cocchia et al., 2011; Restrepo et al., 2012). La calidad del semen equino podría ser determinante en su estado de óxido-reducción, toda vez que se conoce que existe una generación diferencial de ERO entre los espermatozoides no viables o de pobre calidad, respecto a aquellos con metabolismo rápido y altos niveles de fosforilación oxidativa (Gibb et al., 2013). De igual forma, la producción de ERO está ligada a la actividad metabólica de los espermatozoides y depende en gran medida de la disponibilidad de sustancias energéticas (Vasconcelos et al., 2010). Los resultados encontrados evidencian una constante producción de ERO por parte del semen equino fresco, la cual fue inhibida en diferente proporción, según los niveles de suplementación con timol e isoespintanol (figuras 1 y 2). En ambos casos se observó un nivel máximo de inhibición de ERO, a partir del cual se produjo un marcado descenso de la inhibición, en coincidencia con el incremento en la concentración de antioxidante.
Los coeficientes de regresión (Cuadro 1) muestran una reducción pos-descongelación en la velocidad de producción de ERO (β1) por la suplementación del semen equino con timol e isoespintanol, respecto al control. Este efecto es importante dado que la acumulación de ERO en el semen podría asociarse al incremento en las alteraciones estructurales descritas en las células espermáticas (Baumber et al., 2003; Wnuk et al., 2010). Los coeficientes de regresión para la producción de ERO por efecto de la edad del reproductor (β2) fueron menores en el semen suplementado con timol e isoespintanol, respecto al control, indicando una reducción en la generación de ERO asociada a la edad por efecto de ambos antioxidantes. Se ha descrito que los espermatozoides provenientes de los animales de mayor edad producen más radicales libres que los provenientes de animales jóvenes, debido a la reducción de la actividad antioxidante enzimática (Weir y Robaire, 2007).
El sometimiento del semen a procesos biotecnológicos como la congelación produce un incremento en la generación de ERO (Ball, 2008). Esto se ha atribuido, entre otros factores, al aumento en la presencia de células apoptóticas (Brum et al., 2008) y a la centrifugación del semen durante su procesamiento (Vasconcelos et al., 2010). Fue evidente la disminución en la producción de ERO del semen por la suplementación con timol e isoespintanol durante la criopreservación (Cuadro 2), lo cual es acorde con los efectos antioxidantes de ambas moléculas, previamente observados en otros medios y sistemas biológicos (Rojano et al., 2008a; Al-Malki, 2010).
Hay dos mecanismos propuestos por los cuales los antioxidantes pueden desempeñar su función protectora. El mecanismo HAT (transferencia de átomo de hidrogeno) y el mecanismo SET (transferencia de un electrón) (Leopoldini et al., 2004). El efecto antioxidante del timol y el isoespintanol en presencia de agua y metanol siguen un mecanismo de HAT (Rojano et al., 2008a). De esta forma, sería de esperar la acción de un mecanismo HAT en el semen equino diluido por corresponder a un sistema acuoso. En este mecanismo, la ERO retira un átomo de hidrógeno del antioxidante, el cual se convierte en un radical. La estabilidad del radical formado confiere al antioxidante una mayor eficiencia, de modo que sea poco probable que reaccione nuevamente con el sustrato (Özkan y Erdoðan, 2011). La diferencia en la reducción de las ERO por efecto del isoespintanol respecto al timol, podría ser explicada por la mayor capacidad del isoespintanol como atrapador de ERO. La estabilidad del radical formado por la reacción antioxidante, podría a su vez explicarse por las diferentes formas de resonancia de la molécula (ordenamiento de los electrones). El isoespintanol basa su acción antioxidante en su alta estabilidad, reflejada en su potencia derivada de la formación de puentes de hidrógeno intra e intermoleculares, que dejan el centro reactivo del isoespintanol (enlace O-H) disponible para dar electrones a los radicales libres. Por lo tanto, su actividad antioxidante se incrementa respecto al timol, el cual crea puentes intermoleculares con los solventes, disminuyendo así su actividad (Rojano et al., 2008a).
El análisis de los mecanismos de neutralización de las ERO por acción de los isopropilfenoles podría enfocarse en la participación del peróxido de hidrogeno (H2O2), como la ERO predominante en el semen equino, así como en la capacidad de los espermatozoides de generar anión superóxido (O2-•) (Burnaugh et al., 2007). Dado que el isoespintanol tiene una baja capacidad para la captura del anión superóxido (Rojano et al., 2008a), el papel del isoespintanol y el timol como antioxidantes fenólicos podría estar en la interrupción de la segunda etapa de la cadena de propagación de oxidación de lípidos, neutralizando los radicales lipídicos (ROO•) mediante la donación rápida de átomos de hidrógeno (Rojano et al., 2008c), lo cual explicaría la reducción en la producción de ERO en el semen equino por acción de ambas moléculas. Otro mecanismo propuesto sería la inhibición directa del peróxido de hidrógeno, dado que un estudio reciente demostró la capacidad del timol para neutralizar esta ERO (Özkan y Erdoðan, 2011). Sería de esperarse un efecto similar por parte del isoespintanol, toda vez que se conoce su similitud estructural al timol, al igual que su mayor capacidad antioxidante (Rojano et al., 2008a).
La peroxidación de los lípidos constituyentes de las membranas celulares es otro factor asociado con la pérdida de la movilidad, la viabilidad, la integridad de la membrana plasmática y la capacidad para la fertilización de los espermatozoides (Aitken, 1995; Baumber et al., 2000; Neild et al., 2002). Con base a los resultados de esta investigación, la peroxidación lipídica presentó un incremento significativo en el semen equino criopreservado respecto al semen fresco. Este efecto, observado previamente en el semen equino, ha sido atribuido al estrés oxidativo del proceso, conjugado con el alto contenido de ácidos grasos poliinsaturados en las membranas de los espermatozoides de dicha especie (Morte et al., 2008). La peroxidación lipídica ocasiona la pérdida de ácidos grasos poliinsaturados de las membranas, con una producción asociada de hidroperóxidos lipídicos (ROOH), radicales alcoxi (RO-) y radicales peroxilo (ROO•), los cuales promueven una reacción en cadena de peroxidación lipídica y en última instancia conducen a la producción de aldehídos citotóxicos como malondialdehido y 4hidroxinonenal (Aitken, 1995). De otro lado, se ha descrito al radical hidroxilo (HO•) como una especie altamente reactiva con un alto poder de iniciación de la peroxidación lipídica en el semen (Baumber et al., 2003), por lo cual la reducción de la peroxidación lipídica en el semen equino debería estar dirigida a la neutralización de la ERO que la desencadenan. Pese a que el isoespintanol y el timol como antioxidantes fenólicos participarían en la neutralizaciónlos radicales lipídicos(Rojano et al., 2008c), no se evidenció su efecto en la reducción de la peroxidación del semen criopreservado.
Es importante considerar que la presencia de sustancias con acción antioxidante propias del semen, así como de otras disponibles en el diluyente, con un efecto de compensación de la actividad antioxidante (Bustamente et al., 2009), podrían reducir la peroxidación de los lípidos, siendo probable el enmascaramiento de la acción reductora de la peroxidación por parte de los isopropilfenoles, tal como ha sido descrita en otros medios (Rojano et al., 2008b,c). Es posible que el desconocimiento de muchos de los posibles mecanismos enzimáticos y mediados por sustancias de bajo peso molecular, involucrados en la reducción de las ERO en el semen equino, no permita dilucidar los eventos específicos relacionados con el control de la peroxidación. Sin embargo, es importante considerar como factores determinantes, la evidente participación del peróxido de hidrogeno como la principal ERO causante de citotoxicidad en el semen equino (Burnaugh et al., 2007), la inhibición de sistemas enzimáticos a causa de las ERO y de algunos productos de la peroxidación como el malondialdehido y el 4-hidroxinonenal (Baumber et al., 2000), la reducción de alteraciones citológicas por acción de la catalasa y el glutatión reducido (Baumber et al., 2003) y la disminución del estrés oxidativo con el mejoramiento de la calidad del semen refrigerado por efecto de la enzima superóxido dismutasa (SOD) (Cocchia et al., 2011).
Es probable que el marcado incremento en la generación de ERO en el semen criopreservado pudiese limitar el efecto del timol y el isoespintanol sobre la peroxidación lipídica a las concentraciones utilizadas en el estudio. Otro aspecto por considerar sería la reducción de la actividad enzimática en el semen criopreservado por efecto de la remoción del plasma seminal (Pérez-Osorio et al., 2008; Lozano et al., 2011). De otro lado, se ha descrito que una de las propiedades del timol, está en incrementar la actividad de algunas enzimas antioxidantes hepáticas (Al-Malki, 2010), mientras un estudio reciente demostró la capacidad del isoespintanol en la disminución de la actividad de la SOD y GPx en el semen equino diluido con fines de congelación (Restrepo y Rojano, 2017).
CONCLUSIONES
-
La congelación produce un evidente incremento en la producción de especies reactivas de oxígeno y en la peroxidación lipídica del semen equino.
-
El isoespintanol y el timol en diferentes concentraciones tienen la capacidad de reducir la generación de especies reactivas de oxígeno del semen equino.
-
El isoespintanol y el timol tienen un efecto antioxidante en el semen equino sometido a congelación, toda vez que producen una disminución pos-descongelación de la producción de especies reactivas de oxígeno.
-
No se evidenció algún efecto del timol y el isoespintanol en la peroxidación lipídica del semen equino criopreservado.
LITERATURA CITADA
1. Aitken R. 1995. Free radicals, lipid peroxidation and sperm function. Reprod Fertil Dev 7: 659-668. doi: 10.1071/ RD9950659 [ Links ]
2. Aitken R, Smith T, Lord T, Kuczera L, Koppers A, Naumovski N, Connaughton H, et al. 2013. On methods for the detection of reactive oxygen species generation by human spermatozoa: analysis of the cellular responses to catechol oestrogen, lipid aldehyde, menadione and arachidonic acid. Andrology 1: 192-205. doi: 10.1111/ j.2047-2927.2012.00056.x [ Links ]
3. Al-Malki A. 2010. Antioxidant properties of thymol and butylated hydroxytoluene in carbon tetrachloride – induced mice liver injury. JKAU Sci 22: 239-248. doi: 10.4197 / Sci. 22-1.16 [ Links ]
4. Ball B. 2008. Oxidative stress, osmotic stress and apoptosis: impacts on sperm function and preservation in the horse. Anim Reprod Sci 107: 257-267. doi: 10.1016/j.anireprosci.2008.04.014 [ Links ]
5. Ball B, Vo A, Baumber J. 2001. Generation of reactive oxygen species by equine spermatozoa. Am J Vet Res 62: 508-515. [ Links ]
6. Barros L, Silva S, Almeida F, Silva E, Carneiro G, Guerra M. 2012. Effect of addition of acetyl-cysteine and glutathione peroxidase in freezing extender of stallion semen. J Equine Vet Sci 32: 475-518. doi: 10.1016/j.jevs.2012.- 06.026 [ Links ]
7. Baumber J, Ball B, Gravance C, Medina V, Davies-Morel M. 2000. The effect of reactive oxygen species on equine sperm motility, viability, acrosomal integrity, mitochondrial membrane potential and membrane lipid peroxidation. J Androl 21: 895-902. doi: 10.1002/ j.1939-4640.2000.tb03420.x [ Links ]
8. Baumber J, Ball B, Linfor J, Meyers S. 2003. Reactive oxygen species and cryopreservation promote DNA fragmentation in equine spermatozoa. J Androl 24: 621-628. doi: 10.1002/j.1939- 4640.2003.tb02714.x [ Links ]
9. Baumber J, Vo A, Sabeur K, Ball B. 2002. Generation of reactive oxygen species by equine neutrophils and their effect on motility of equine spermatozoa. Theriogenology 57: 1025-1033. doi: 10.1016/S0093-691X(01)00710-5 [ Links ]
10. Blanes-Zamora R, Vaca-Sánchez R, González-Pérez J, Báez-Quintana D, Rodríguez-Díaz R, Bethencourt J. 2008. El uso de pentoxifilina mejora la movilidad espermática en los casos de microinyección espermática con biopsia testicular. Rev Int Androl 6: 242-248. doi: 10.1016/S1698-031X(08)76154-5 [ Links ]
11. Brito L, Greene L, Kelleman A, Knobbe M, Turner R. 2011. Effect of method and clinician on stallion sperm morphology evaluation. Theriogenology 76: 745-750. doi: 10.1016/j.theriogenology. 2011.04.007 [ Links ]
12. Brum A, Sabeur K, Ball B. 2008. Apoptotic-like changes in equine spermatozoa separated by densitygradient centrifugation or after cryopreservation. Theriogenology 69: 1041-1055. doi: 10.1016/j.theriogenology. 2008.01.014 [ Links ]
13. Burnaugh L, Sabeur K, Ball B. 2007. Generation of superoxide anion by equine spermatozoa as detected by dihydroethidium. Theriogenology 67: 580-589. doi: 10.1016/j.theriogenology.2006.07.021 [ Links ]
14. Bustamante I, Pederzolli C, Sgaravatti A, Gregory R, Dutra C, Jobim M, Mattos R. 2009. Skim milk-egg yolk based semen extender compensates for non-enzymatic antioxidant activity loss during equine semen cryopreservation. Anim Reprod 6: 392-399. [ Links ]
15. Cocchia N, Pasolinia M, Mancini R, Petrazzuolo O, Cristofaro I, Rospane I, Sica A, et al. 2011. Effect of sod (superoxide dismutase) protein supplementation in semen extenders on motility, viability, acrosome status and ERK (extracellular signal-regulated kinase) protein phosphorylation of chilled stallion spermatozoa. Theriogenology 75: 1201-1210. doi: 10.1016/j.theriogenology. 2010.11.031 [ Links ]
16. Coyan K, Baspinar N, Numan M, Peker P. 2011. Effects of cysteine and ergothioneine on post-thawed Merino ram sperm and biochemical parameters. Cryobiology 63: 1-6. doi: 10.1016/ j.cryobiol.2011.04.001 [ Links ]
17. Gamboa S, Ramalho-Santos J. 2005. SNARE proteins and caveolin-1 in stallion spermatozoa: possible implications for fertility. Theriogenology 64: 275-291. doi: 10.1016/j.theriogenology.2004.11.021 [ Links ]
18. Gibb Z, Butler T, Morris L, Maxwell W, Grupen C. 2013. Quercetin improves the postthaw characteristics of cryopreserved sex-sorted and nonsorted stallion sperm. Theriogenology 79: 1001- 1009. doi: 10.1016/j.theriogenology. 2012.06.032 [ Links ]
19. Hussain J, Salam A, Gohar A. 2011. A study on the cryopreservation of stallion semen with alpha lipoic acid. Intl R J Pharmaceut 1: 21-26. [ Links ]
20. Isachenko V, Isachenko E, Katkov I, Montag M, Dessole S, Nawroth F, Van Der Ven H. 2004. Cryoprotectant-free cryopreservation of human spermatozoa by vitrification and freezing in vapor: effect on motility, DNA integrity, and fertilization ability. Biol Reprod 71: 1167- 1173. doi: 10.1095/biolreprod.104.028811 [ Links ]
21. Lagouri V, Boskou D. 1995. Screening for antioxidant activity of essential oils obtained from spices. Dev Food Sci 37: 869-879. doi: 10.1016/S0167-4501(06)- 80203-7 [ Links ]
22. Leopoldini M, Marino T, Russo N, Toscano M. 2004. Antioxidant properties of phenolic compounds. Hatom versus electron transfer mechanism. J Phys Chem 108: 4916-4922. doi: 10.1021/jp037247d [ Links ]
23. Lisboa F, Hartwig F, Maziero R, Monteiro G, Papa F, Dell’aqua J. 2012. Use of L-carnitine and acetyl-Lcarnitine in cooled-stored stallion semen. J Equine Vet Sci 32: 493-494. doi: http:// dx.doi.org/10.1016/j.jevs.2012.06.057
24. Lozano D, Gil L, Álvarez C. 2011. Efecto de la adición de plasma seminal en el semen equino descongelado. Sanid Mil 67: 284-290. doi: 10.4321/S1887- 85712011000400005 [ Links ]
25. Macias-Garcia B, Gonzalez- Fernandez L, Ortega-Ferrusola C, Morillo-Rodriguez A, Gallardo- Bolanos J, Rodriguez-Martinez H, Tapia J, et al. 2011. Fatty acids and plasmalogens of the phospholipids of the sperm membranes and their relation with the post-thaw quality of stallion spermatozoa. Theriogenology 75: 811- 818. doi: 10.1016/j.theriogenology. 2010.10.021 [ Links ]
26. Martins H, Brito M, Sampaio I, Stahlberg R, Souza M, Penna C, Lagares M. 2015. 67 effects of antioxidants lactoferrin and catalase on stallion frozen semen. Reprod Fertil Dev 27: 126-127. doi: https://doi.org/10.1071/ RDv27n1Ab67 [ Links ]
27. Martins H, Souza M, Penna C, Da Silva G, Côrtes S, Stahlberg R, Brito M, et al. 2012. Effects of lactoferrin supplementation to milk- and caseinatebased extenders on sperm motility, membrane integrity and nitric oxide levels of cooled stallion semen. J Equine Vet Sci 32: 475-518. doi: 10.1016/ j.jevs.2012.06.060 [ Links ]
28. Morte M, Rodrigues A, Soares D, Rodrigues A, Gamboa S, Ramalho- Santos J. 2008. The quantification of lipid and protein oxidation in stallion spermatozoa and seminal plasma: seasonal distinctions and correlations with DNA strand breaks, classical seminal parameters and stallion fertility. Anim Reprod Sci 106: 36-47. doi: 10.1016/ j.anireprosci.2007.03.020 [ Links ]
29. Neild D, Gadella B, Colendrander B, Agüero A, Brouwers J. 2002. Lipid peroxidation in stallion spermatozoa. Theriogenology 58: 295-298. doi: 10.1016/S0093-691X(02)00799-9 [ Links ]
30. Ortega-Ferrusola C, García B, Gallardo- Bolaños J, Gonzáles- Fernández L, Rodriguez-Martinez H, Tapia J, Peña F. 2009. Apoptotic markers can be used to forecast the freezeability of stallion spermatozoa. Anim Reprod Sci 114: 393-403. doi: 10.1016/j.anireprosci.2008.10.005 [ Links ]
31. Özkan A, Erdoðan A. 2011. A comparative evaluation of antioxidant and anticancer activity of essential oil from Origanum onites (Lamiaceae) and its two major phenolic components. Turk J Biol 35: 735-742. doi: 10.3906/biy-1011-170 [ Links ]
32. Pérez-Osorio J, Mello F, Juliani G, Lagares M, Lago L, Henry M. 2008. Effect on post-thaw viability of equine sperm using stepwise addition of dimethyl formamide and varying cooling and freezing procedures. Anim Reprod 5: 103-109. [ Links ]
33. Restrepo G, Duque J, Montoya J. 2012. Effect of two protocols of cryopreservation on fertilizing capacity of stallion (Equus caballus) semen. Rev Fac Nal Agr Medellín 65: 6711-6718. [ Links ]
34. Restrepo G, Rojano B. 2017. Efecto del isoespintanol y el timol en la actividad antioxidante de semen equino diluido con fines de congelación. Rev Med Vet 35: 149-158. doi: 10.19052/mv.4397 [ Links ]
35. Rojano B, Gaviria C, Gil M, Saez J, Schinella G, Tournier H. 2008a. Actividad antioxidante del isoespintanol en diferentes medios. Vitae 15: 173-181. [ Links ]
36. Rojano B, Gaviria C, Saez J. 2008b. Determinación de la actividad antioxidante en un modelo de peroxidación lipídica de mantequilla inhibida por el isoespintanol. Vitae 15: 212-218. [ Links ]
37. Rojano B, Peréz E, Figadère B, Martin M, Recio M, Giner R, Rios J, et al. 2007. Constituents of Oxandra cf. xylopioides with anti-inflammatory activity. J Nat Prod 70: 835-838. doi: 10.1021/np060333v [ Links ]
38. Rojano B, Saez J, Schinella G, Quijano J, Vélez E, Gil A, Notario R. 2008c. Experimental and theoretical determination of the antioxidant properties of isoespintanol (2-Isopropyl- 3,6-dimethoxy- 5-me thylphenol). J Mol Struct 877: 1-6. doi: https://doi.org/ 10.1016/j.molstruc.2007.07.010 [ Links ]
39. Stephens T, Brooks R, Carrington J, Cheng L, Carrington A, Porr C, Splan R. 2013. Effects of pentoxifylline, caffeine, and taurine on post-thaw motility and longevity of equine frozen semen. J Equine Vet Sci 33: 615-621. doi: 10.1016/j.jevs.2012.10.004 [ Links ]
40. Vasconcelos A, Santana M, Santos A, Santero M, Lagares M. 2010. Metabolic evaluation of cooled equine spermatozoa. Andrología 42: 106-111. doi: 10.1111/j.1439-0272.2009.00963.x [ Links ]
41. Vasconcelos J, Chaveiro A, Góis A, Moreira da Silva F. 2013. Effects of á-tocopherol and ascorbic acid on equine semen quality after cryopreservation. J Equine Vet Sci 33: 787-793. doi: https:// doi.org/10.1016/j.jevs.2012.12.012 [ Links ]
42. Weir C, Robaire B. 2007. Spermatozoa have decreased antioxidant enzymatic capacity and increased reactive oxygen species production during aging in the Brown Norway rat. J Androl 28: 229- 240. doi: 10.2164/jandrol.106.001362 [ Links ]
43. Wnuk M, Lewinska B, Oklejewicz G, Bartosz M, Tischner M, Bugno- Poniewierska M. 2010. Redox status of equine seminal plasma reflects the pattern and magnitude of DNA damage in sperm cells. Theriogenology 74: 1677- 1684. doi: 10.1016/j.theriogenology. 2010.07.007 [ Links ]
Recibido: 29 de agosto de 2017
Aceptado para publicación: 2 de enero de 2018

