










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versión impresa ISSN 1609-9117
Rev. investig. vet. Perú vol.29 no.1 Lima ene./mar. 2018
http://dx.doi.org/10.15381/rivep.v29i1.14194
ARTÍCULOS PRIMARIOS
Detección de una proteína asociada a la enfermedad de la necrosis hepatopancreática aguda (AHPND) en Litopenaeus vannamei bajo cultivo semi-intensivo en Ecuador
Detection of a protein associated with acute hepatopancreatic necrosis disease (AHPND) in Litopenaeus vannamei under semi-intensive farming in Ecuador
Katherine Yuliana Saavedra-Olivos1, Tessy Peralta-Ortiz1, Alberto Ordinola-Zapata1,2, John Estuardo Sandoval-Ramayoni1, Enedia Graciela Vieyra-Peña1, Marco Antonio Zapata-Cruz1, Auberto Hidalgo-Mogollón1, Braulio Morán-Ávila1, Oscar Augusto Mendoza-Neyra1, Magno Ego Mendoza-Dioses1, Sigrid Yamile Campoverde-Peña1
1 Facultad de Ingeniería Pesquera y Ciencias del Mar, Universidad Nacional de Tumbes, Puerto Pizarro, Tumbes, Perú
2 E-mail: aordinolaz@untumbes.edu.pe
RESUMEN
La presente investigación tuvo como objetivo detectar una proteína asociada a la necrosis hepatopancreática aguda (AHPND), en cultivos semi-intensivos en Ecuador. Se recolectaron camarones enfermos de tres camaroneras en la zona de Bellavista, provincia El Oro. Los hepatopáncreas fueron macerados y cultivados en medio TCBS y subcultivos en TSA y caldo LB. De las cepas bacterianas obtenidas, se extrajo las proteínas usando un kit comercial y se separaron mediante migración en gel SDS-PAGE. Estas fueron analizadas con un espectrómetro de masas MALDI TOF/TOF. La confirmación de las cepas se realizó mediante PCR utilizando cebadores TUMSAT-Vp3, que son específicos para detectar AHPND. Una de las cepas tuvo secuencias peptídicas similaresa lade la proteína PirvpB, causantedeAHPND, yfueidentificadacomo perteneciente a Vibrio parahaemolyticus y portadora del gen que codifica PirvpB. Los resultados mostraron que es posible usar la espectrometría de masas MALDI TOF/TOF en la detección de proteínas asociadas a AHPND en el cultivo de camarón.
Palabras clave: camarón; AHPND; Litopenaeus vannamei; síndrome de mortalidad precoz; Vibrio parahaemolyticus; PirvpB
ABSTRACT
The present study aimed to detect a protein associated with acute hepatopancreatic necrosis (AHPND) by mass spectrometry, reared under semi-intensive farming in Ecuador. Sick shrimps from three farms were collected in the Bellavista area in the El Oro province. The hepatopancreas were macerated and cultured in TCBS medium and subcultured in TSA and LB broth. In the bacterial strains obtained, the proteins were extracted using a commercial kit and separated by SDS-PAGE gel migration. These were analyzed with a MALDI TOF/TOF mass spectrometer. The confirmation of the strains was performed by PCR using TUMSAT-Vp3 primers, which are specific for detecting AHPND. One of the strains had peptide sequences similar to that of the PirvpB protein causing AHPND, and was identified as belonging to Vibrio parahaemolyticus and carrying the gene coding for PirvpB. The results showed that it is possible to use MALDI TOF/TOF mass spectrometry in the detection of AHPND-associated proteins in shrimp culture.
Keywords: shrimp; AHPND; Litopenaus vannamei; early mortality syndrome; Vibrio parahaemolitycus; PirvpB
INTRODUCCIÓN
La necrosis hepatopancreática aguda (AHPND), llamada también síndrome de mortalidad precoz (EMS), es una enfermedad que ocasiona grandes pérdidas económicas en el cultivo de camarones; tanto del camarón blanco (Litopenaeus vannamei) como del camarón tigre (Penaeus monodon) (De la Peña et al., 2015). Esta enfermedad es atribuida a una cepa de Vibrio parahaemolyticus que alberga el plásmido pVPA3-1 o pVA1 de 70 kpb (Lee et al., 2015), que contiene genes codificantes de toxinas binarias tipo Photorhabdus insect related (Pir): PirvpA y PirvpB, que inducen la muerte celular (Otta et al., 2014; Soto-Rodríguez et al., 2015).
La AHPND se caracteriza por una severa atrofia del hepatopáncreas, que se muestra de un color pálido o blanco con manchas o rayas negras, desprendimiento masivo de células epiteliales de los túbulos; así como letargia, exoesqueleto blando, crecimiento lento, estómago e intestino semi vacío (Flegel, 2012; Tran et al., 2013; Otta et al., 2014; Hong et al., 2016).
La enfermedad afecta principalmente en los estanques de cultivo a las pos-larvas de camarones en los primeros 30 días después de la siembra (Flegel, 2012; Otta et al., 2014). Los primeros episodios de mortalidad causados por AHPND fueron reportados en China en 2009, pasando a Vietnam (2010), Malasia (2011), Tailandia y Filipinas (2012), llegando a América en 2013, afectando cultivos en México (Nunan et al., 2014; De la Peña et al., 2015; Arunrut et al., 2016; Thitamadee et al., 2016). No existen reportes de AHPND en Ecuador ni en Sudamérica, pero se sospecha que puede estar presente en varios países de Asia, Latinoamérica y El Caribe en los que aún no ha sido reportado (Bondad-Reantaso, 2016).
Existen diversas técnicas para la detección de la AHPND, tales como el análisis histopatológico, la hibridación in situ y la reacción de la cadena de la polimerasa (PCR), dirigida esta última a los genes que codifican a las toxinas PirvpA y PirvpB de Vibrio parahaemolyticus (Tinwongger et al., 2014; Han et al., 2015; Lee et al., 2015; Sirikharin et al., 2015).
Una herramienta que podría ser utilizada para la detección de esta enfermedad es la espectrometría de masas que permite el análisis de proteínas características del patógeno. Esta técnica tiene la ventaja de poder detectar cantidades muy pequeñas de péptidos, incluso al nivel de attomoles (Hillenkamp et al., 1991; Vorm et al., 1994; Singhal et al., 2015). Entre las técnicas de espectrometría de masas (MS), la ionización/ desorción láser asistida por matriz en tiempo de vuelo (MALDI TOF) ha mostrado mayores ventajas dada su alta sensibilidad, tolerancia a buffers y rápida adquisición de datos, además de una instrumentación sencilla (Vorm et al., 1994); por lo que podría ser utilizada en la identificación rápida del patógeno causante de la AHPND. Para lograr esto, sería necesario determinar las proteínas características del patógeno que podrían ser detectadas con esta técnica.
Debido a que han habido últimamente eventos de gran mortalidad en ciertos cultivos de camarón en Ecuador, donde algunos camarones mostraron signos similares al AHPND, se realizó la presente investigación con el objetivo de evaluar si la técnica de espectrometría de masas MALDI TOF-TOF podría detectar proteínas asociadas a un posible caso de AHPND en los cultivos de camarón afectados, para de esta manera tener una nueva técnica diagnóstica rápida y precisa para la detección de la enfermedad.
MATERIALES Y MÉTODOS
Obtención de Muestras
La recolección de camarones se realizó entre abril y octubre de 2016, en tres empresas camaroneras que reportaron mortalidades en sus cultivos y que se hallan ubicadas en la zona de Bellavista del cantón Santa Rosa en la provincia de El Oro, Ecuador (Figura 1). Se recolectaron 150 camarones de 1 a 9 g, que fueron transportados vivos en bolsas plásticas al Laboratorio de Biología Molecular de la Facultad de Ingeniería Pesquera y Ciencias del Mar de la Universidad Nacional de Tumbes, ubicado en la región Tumbes (Perú).
Cultivos Bacterianos
Se extrajo el hepatopáncreas de los camarones, en forma aséptica, y se colocaron en tubos de 1.5 ml (Axygen) conteniendo 500 µl de solución salina al 2%, donde fueron macerados. Se tomó 20 µl del macerado y se sembró en placas petri (Normax) conteniendo medio tiosulfato citrato bilis sucrosa (TCBS) (Merck) ajustado al 2% de NaCl (Merck). Todas las placas fueron incubadas a 37 °C durante 24 h. Las colonias bacterianas no fermentadoras de sucrosa (de color verde) que crecieron en el medio fueron subcultivadas en agar tripticasa de soya (TSA) (Oxoid) hasta obtener aislados puros. Posteriormente, las cepas bacterianas puras se suspendieron de manera independiente en 1 ml de caldo Luria Bertani (LB) (Invitrogen) ajustado al 2% de NaCl (Merck) y se incubaron a 37 °C durante 24 h.
Extracción de Proteínas Bacterianas
Se tomó 1 ml de caldo LB de cada uno de los cultivos puros y las proteínas se extrajeron con el kit comercial Qproteome Bacterial Protein Prep Kit (Qiagen) siguiendo las especificaciones del fabricante. El sobrenadante conteniendo las fracciones de proteínas solubles fue teñido con buffer de carga (0.125 M de Tris (Promega) pH 6.8; 4% de dodecil sulfato de sodio SDS (Merck), 20% de glicerol (Merck), 0.02% de azul de bromofenol (Sigma-Aldrich), 0.2 M de DTT (OmniPlus) y agua grado HPLC (Merck) e incubado a 95 °C durante 5 min.
Se preparó gel de poliacrilamida, conformado por un gel separador al 15%, el cual se preparó con 3.4 ml de agua bidestilada, 7.5 ml de mix de acrilamida al 30% (29.2% de acrilamida [AppliChem], 0.8% de bisacrilamida [AppliChem] y agua bi-destilada), 3.8 ml de Tris (Promega) 1.5 M pH 8.8, 0.15 ml de SDS (Merck) al 10%, 0.075 ml de persulfato de amonio (APS) (AppliChem) al 10% y 0.0075 ml de TEMED (AppliChem); además de un gel de alineamiento (concentrador), preparado con 3.4 ml de agua bi-destilada, 0.83 ml de mix de acrilamida al 30%, 0.63 ml de Tris 1 M pH 6.8, 0.05 ml de SDS al 10%, 0.05 ml de APS al 10% y 0.005 ml de TEMED. Como tampón de migración se utilizó TANK 1X el cual se diluyó a partir de TANK 5X (0.124 M de Tris [Promega], 0.96M de glicina [Sigma-Aldrich], 0.5% de SDS [Merck] y agua bidestilada [Merck]). La separación de proteínas se realizó por migración electroforética a 90 V durante 3 h en el gel SDS-PAGE al 15%.
Las bandas presentes en el gel SDSPAGE se sometieron por 30 min a una solución de fijación conteniendo 50% de metanol (Merck), 10% de ácido acético (Merck) y 40% de agua grado HPLC. Luego, el gel fue colocado toda la noche en solución de tinción conteniendo 50% de metanol, 10% de ácido acético, 0.1% de azul de coomassie R-250 y agua grado HPLC. Finalmente fueron decoloradas con solución de lavado conteniendo 45% de metanol (Merck), 10% de ácido acético (Spectrum) y 45% de agua grado HPLC (Merck).
Recuperación y Digestión de Proteínas
La digestión de proteínas se realizó siguiendo el procedimiento descrito por Shevchenko et al. (2006) modificado, donde las proteínas de interés que migraron en el gel SDS-PAGE fueron recuperadas cortando con un bisturí estéril las secciones del gel que las contenían. Estas secciones fueron decoloradas en 50 µl de bicarbonato de amonio (NH4HCO3) (Sigma-Aldrich) 100 mM y 50 µl de acetonitrilo (Merck) e incubadas a 37 °C durante 30 min, agitando ocasionalmente. Luego fueron deshidratadas con 100 µl de acetonitrilo absoluto e incubadas a temperatura ambiente durante 30 min; para posteriormente ser tratadas con 30 µl de buffer de digestión (13 ng/µl de tripsina [Promega], 50 mM de NH4HCO3, pH 8.0) e incubadas a 4 °C durante 2 h. Finalmente, se volvió a incubar a 37 °C durante 18 h.
Identificación de Proteínas
Las muestras digeridas fueron mezcladas con 1 µl de matriz ácido α-ciano-4-hidroxicinámico (CHCA) a concentración de 10 mg/ml en 0.1% de ácido trifluroacético y 50% de acetonitrilo (ACN) y depositadas sobre una placa MALDI AB SCIEX. Los espectros de masa fueron adquiridos usando un espectrómetro de masas MALDI-TOF/TOF ABI SCIEX TOF/TOF TM 5800 SYSTEM (AB Sciex, EEUU) en modo reflectrón (1 kV) con un promedio de 750 disparos del láser, analizándose un rango de masas de 400 a 4000 Da. La calibración previa se realizó usando el kit de calibración del instrumento AB SCIEX TOF/TOF, siguiendo las indicaciones del fabricante.
Se seleccionaron como iones precursores para la fragmentación aquellos con razón masa/carga (m/z) de 2287.0730 y 2286.9246. Las secuencias obtenidas en la fragmentación fueron procesadas a través del software Protein Pilot (AB Sciex, EEUU), utilizando para ello bases de datos de proteínas relacionadas a AHPND disponibles en el sitio web de Uniprot (http://www.uniprot.org). Dichas secuencias también fueron analizadas con la base de datos de proteínas del National Center for Biotechnology Information (NCBI) (http:/ /blast.ncbi.nlm.nih.gov/Blast.cgi? PAGE=Proteins), mediante la herramienta Basic LocalAlignment Search Tool (BLAST) (Altschul et al., 1997).
Extracción de ADN de Cepas Bacterianas
Se centrifugó 1 ml de la suspensión bacteriana a 15 493 g durante 10 min, se eliminó el sobrenadante y se realizaron lavados con solución salina al 2%. Para la lisis celular se adicionó el buffer TENS (50 mM de Tris-HCl [Promega], 50 mM de EDTA [AppliChem], 100 mM de NaCl [Merck], 1% de SDS [Merck]) y 5 µl de proteinasa K (Merck) (20 mg/ml). Luego se incubó a 55 °C durante 2 h. Posteriormente, se adicionaron 300 µl de fenol (Applichem), 288 µl de cloroformo (Merck) y 12 de alcohol isoamil (Merck) (25:24:1), se homogenizó y se centrifugó a 15 493 g durante 10 min. Se recuperó 300 µl del sobrenadante en un nuevo tubo de 1.5 ml y se adicionó solución de cloroformo y alcohol isoamil (24:1) en igual proporción que el sobrenadante recuperado. Seguidamente se centrifugó a 1493 g durante 10 min. El sobrenadante fue transferido en un tubo nuevo y se le adicionó el doble de su volumen de etanol (Spectrum) al 95% y se centrifugó a 15 493 g durante 10 min. Se eliminó el sobrenadante y el pellet fue lavado con 1 ml de etanol al 75% centrifugándose a 9167 g durante 10 min. El sobrenadante fue eliminado y el ADN sedimentado se dejó secar a temperatura ambiente. Finalmente, se resuspendió en 50 µl de TE (10 mM Tris [Promega] y 1 mM EDTA [Applichen]).
Identificación de Cepas Bacterianas mediante ARNr 16S
El ADN de las cepas puras fue utilizado para realizar la PCR con los cebadores ARNr 16S para bacterias: w001 (5´AGAGTTTGATYMTGGCTCAG-3´) y w002 (5´-GNTACCTTGTTACGACTT-3´) que amplifican un producto de 1500 pares de bases (pb) (Godon et al., 1997). El volumen de reacción fue de 25 µl: 12.5 µl de PCR Master Mix 2X, 10.5 µl de agua (Thermo Scientific),0.5 µlde cada cebador (0.4 pmol/µl) y 1 µl de ADN. Se consideró como control negativo agua estéril y como control positivo el ADN de una cepa bacteriana aislada del camarón previamente identificada.
Las condiciones de amplificación fueron: desnaturalización inicial a 94 °C durante 3 min, seguido de 35 ciclos de desnaturalización a 95 °C durante 30 s, alineamiento a 55 °C durante 45 s, extensión a 72 °C durante 5 min, seguidos de un ciclo final de extensión final a 72 °C durante 5 min. Las reacciones de PCR fueron realizadas en un termociclador de 24 pozos Piko™ series (Thermo Scientific, EEUU). Los productos amplificados fueron visualizados mediante electroforesis en un gel de agarosa al 1.5% teñido previamente con 3 µl de bromuro de etidio a una concentración de 10 mg/ml.
La secuenciación la realizó la Empresa Macrogen (Maryland, EEUU), empleándose 15 µl del producto de la PCR y los cebadores universales para bacterias F518 (5´CCAGCAGCCGCGGTAATACG-3´) y R800 (5´-TACCAGGGTATCTAATCC -3´) (Lane, 1991). Las secuencias obtenidas fueron editadas y alineadas en el programa informático MEGA v. 5. Para determinar la afiliación filogenética se realizaron búsquedas de similitud utilizandoel programaBLAST (Altschul et al., 1997).
Diagnóstico de AHPND mediante PCR
Las mismas cepas bacterianas fueron sometidas a PCR con cebadores específicos para la detección de la AHPND: TUMSATVp3F(5´-GTGTTGCATAATTTTGTGCA-3´) y TUMSAT-Vp3 R (5´-TTGTACAGAAACCACGACT-3´) que amplifican un producto de 360 pb (Tinwongger et al., 2014). Las condiciones de amplificación fueron: desnaturalización inicial a 95 °C durante 2 min, 30 ciclos de desnaturalización 95 °C durante 30 s, alineamiento a 56 °C durante 30 s y elongación a 72 °C durante 30 s, seguido de una extensión final a 72 °C durante 5 min. Los productos amplificados fueron visualizados en un gel de agarosa al 1.5 % teñido con 3 µl de bromuro de etidio a una concentración de 10 mg/ml. La secuenciación la realizó la empresa Macrogen (Maryland, EEUU), con los mismos cebadores de amplificación de los fragmentos para AHPND, y la búsqueda de similitud se hizo con el programa BLAST.
RESULTADOS
Del cultivo realizado en medio TCBS a partir de muestras de hepatopáncreas de L. vannamei con signos clínicos de AHPND se aislaron 25 cepas no fermentadoras de sacarosa, que mediante caracterización molecular por el ARNr 16S se identificaron como especies del género Vibrio, entre ellas V. parahaemolyticus y V. alginolyticus.
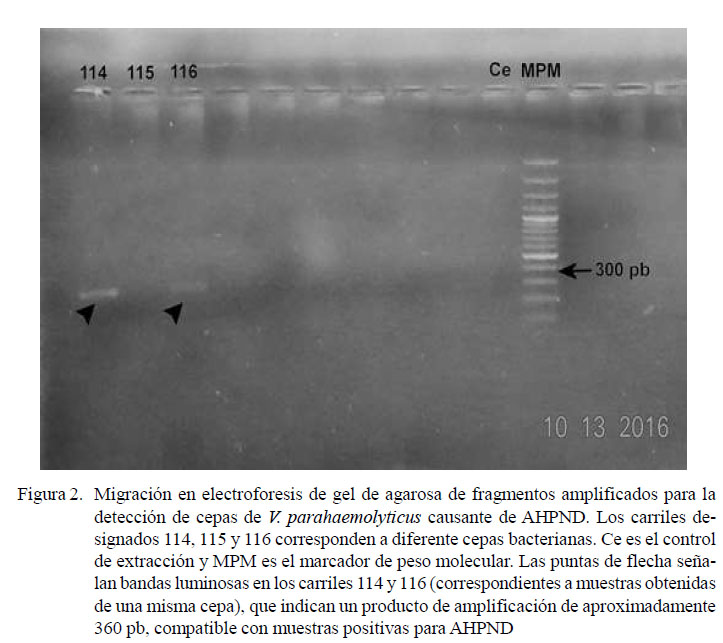
De las 25 cepas bacterianas caracterizadas mediante el análisis de un fragmento del gen ARNr 16S, dos de ellas resultaron positivas al ser amplificadas con los cebadores TUMSAT-Vp3 específicos para AHPND, originando un fragmento de ADN del tamaño esperado (360 pb) (Figura 2). Estas cepas fueron aisladas a partir de un camarón de cerca de 1 g de peso. El alineamiento y posterior análisis en Blast indicó gran similitud (99%) de la secuencia en estudio, con la secuencia de V. parahaemolyticus strain V.03 hypothetical protein, PirA-like toxin, and PirB-like toxin genes, complete cds.
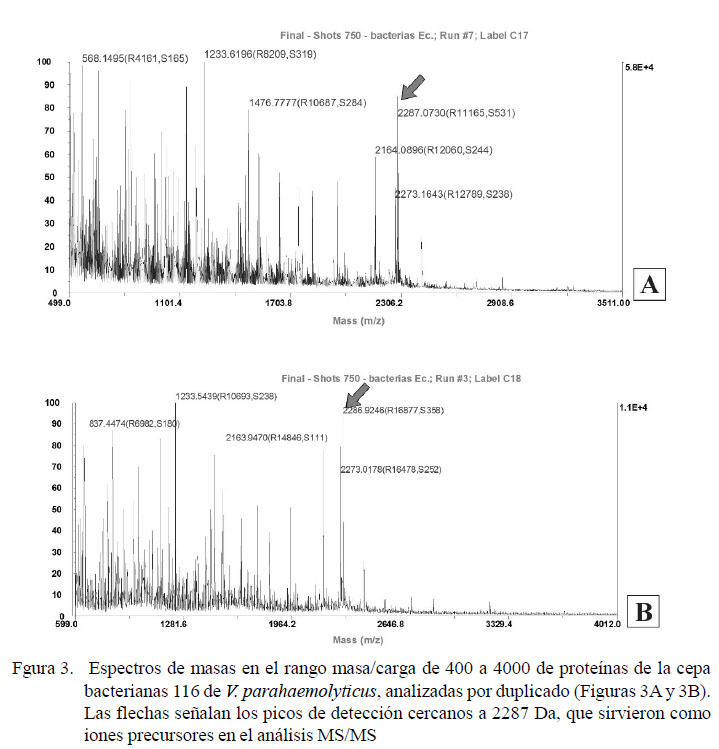
En la Figura 3 se muestra el perfil de espectro de masas de las proteínas en el rango aproximado de 400 a 4000 Da de una cepa bacteriana positiva a AHPND mediante PCR, el cual fue utilizado para seleccionar iones con razón masa/carga (m/z) de 2287.0730 y 2286.9246 como iones precursores y fragmentados. Las secuencias peptídicas obtenidas para los fragmentos fueron analizadas con el software Protein Pilot, detectándose varios fragmentos similares a la proteína PirvpB de V. parahaemolyticus, en particular la secuencia aminoacídica TNEYVVTMSSLTEFNPNNAR, obtenida al analizar los péptidos de la cepa investigada. Estas tuvieron un nivel de confianza del 85% al ser comparadas con la secuencia JHE-Like toxin PirB-like OS=V. parahaemolyticus de las bases de datos Toxin Shrimp y AHPND de Uniprot.
DISCUSIÓN
En esta investigación se detectó una cepa bacteriana asociada a mortalidades de camarón en cultivo en Ecuador, que fue identificada como correspondiente a V. parahaemolyticus. En ella, además, se pudo aislar un fragmento peptídico correspondiente a la proteína PirvpB. El análisis de PCR y la posterior secuenciación también demostraron que las cepas (incluyendo la anteriormente mencionada) presentaron los genes de las toxinas tipo PirvpA y PirvpB, características de cepas causante de AHPND (Otta et al., 2014; Han et al., 2015; Soto-Rodríguez et al., 2015).
Se empleó la espectrometría de masas como criterio diagnóstico para la AHPND tal como también lo hicieron Han et al. (2015), quienes lograron detectar proteínas Pirvp, mostrando la utilidad de dicha técnica para identificar las toxinas de la AHPND. Esta técnica viene siendo utilizada con mayor frecuencia para la detección de Vibrio en muestras humanas en hospitales (Dieckmann et al., 2010) por lo que debe ser considerada como una técnica muy prometedora para diagnóstico rápido, preciso y altamente sensible (Hillenkamp et al., 1991; Vorm et al., 1994; Shevchenko et al., 1996).
Asimismo, se confirmó por PCR que la cepa mencionada portaba el plásmido con los genes que codifican para las toxinas causantes de AHPND. La detección de una secuencia correspondiente al plásmido por PCR es confiable, pues se utilizaron los cebadores de TUMSAT-Vp3, en los cuales se ha reportado hasta 100% de precisión al evitar falsos positivos (Tinwongger et al., 2014).
A nivel de Latinoamérica AHPND ha sido reportada solo en México (Nunan et al., 2014; Soto-Rodríguez et al., 2015), realizándose la detección mediante técnicas histológicas y PCR; así mismo, Restrepo et al. (2016), valiéndose de análisis genómicos, reportan una cepa patogénica de V. parahaemolyticus en camarones de Sudamérica con síntomas de AHPND. En la presente investigación se pudo lograr la identificación de una proteína asociada aAHPND en Ecuador usando espectrometría de masas, la cual fue confirmada con la identificación de la cepa bacteriana mediante secuenciación de un fragmento del genARNr 16S y de un fragmento del gen codificador de la toxina PirVpB mediante PCR, lo que indica que las cepas encontradas pueden originar AHPND.
Sería recomendable que se mantenga una vigilancia sanitaria adecuada en los cultivos de camarón en Ecuador, para determinar si existen posibles casos de presencia de AHPND en los mismos o si lo encontrado en esta investigación se trata de un caso aislado.
CONCLUSIONES
-
Se logró detectar, mediante MS MALDI TOF/TOF, la secuencia peptídica TNEYVVTMSSLTEFNPNNAR asociada a la toxina PirvpB causante de AHPND.
-
Mediante MS MALDI TOF/TOF y PCR se ha detectado por primera vez la proteína PirvpB en una cepa bacteriana aislada de camarones cultivados en Ecuador.
Agradecimientos
Se agradece a los empresarios camaroneros de la zona de Bellavista (Ecuador) que proporcionaron las muestras para esta investigación, al Círculo de Estudiantes de la FIPCM (Universidad Nacional de Tumbes, Perú) por el apoyo en el procesamiento de las muestras y al Laboratorio Costero Tumbes del Instituto del Mar del Perú (IMARPE).
LITERATURA CITADA
1. Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. 1997. Gapped BLAST and PSIBLAST: a new generation of protein database search programs. Nucl Acids Res 25: 3389-3402. doi: 10.1093/nar/ 25.17.3389 [ Links ]
2. Arunrut N, Kampeera J, Sirithammajak S, Sanguanrut P, Proespraiwong P, Suebsing R, Kiatpathomchai W. 2016. Sensitive visual detection of AHPND bacteria using loop-mediated isothermal amplification combined with DNA-functionalized gold nanoparticles as probes. PLoS One 11(3): e0151769. doi: 10.1371/journal.- pone.0151769 [ Links ]
3. Bondad-Reantaso MG. 2016. Acute hepatopancreatic necrosis disease (AHPND) of penaeid shrimps: global perspective. In: Pakingking Jr RV, de Jesus-Ayson EGT, Acosta BO (eds). Addressing acute hepatopancreatic necrosis disease (AHPND) and other transboundary diseases for improved aquatic animal health in Southeast Asia. [Internet]. Available in https:// repository.seafdec.org.ph/bitstream/ h a n d l e / 1 0 8 6 2 / 3 0 8 4 / B o n d a d - ReantasoMG2016.pdf?sequence=3 [ Links ]
4. De la Peña LD, Cabillon NAR, Catedral DD, Amar EC, Usero RC, Monotilla WD, Calpe AT, et al. 2015. Acute hepatopancreatic necrosis disease (AHPND) outbreaks in Penaeus vannamei and P. monodon cultured in the Philippines. Dis Aquat Organ 116: 251-254. doi: 10.3354/dao02919 [ Links ]
5. Dieckmann R, Strauch E, Alter T. 2010. Rapid identification and characterization of Vibrio species using wholecell MALDI-TOF mass spectrometry. J Appl Microbiol 109: 199-211. doi: 10.1111/ j.1365-2672.2009.04647.x [ Links ]
6. Flegel TW. 2012. Historic emergence, impact and current status of shrimp pathogens in Asia. J Invertebr Pathol 110: 166-173. doi: 10.1016/j.jip.2012.03.004 [ Links ]
7. Godon JJ, Zumstein E, Dabert P, Habouzit F, Moletta, R. 1997. Molecular microbial diversity of an anaerobic digestor as determined by small-subunit rDNA sequence analysis. Appl Environ Microbiol 63: 2802- 2813. [ Links ]
8. Han JE, Tang KFJ, Tran LH, Lightner DV. 2015. Photorhabdus insect-related (Pir) toxin-like genes in a plasmid of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease (AHPND) of shrimp. Dis Aquat Organ 113: 33-40. doi: 10.3354/dao02830 [ Links ]
9. Hillenkamp F, Karas M, Beavis RC, Chait BT. 1991. Matrix-assisted laser desorption/ionization mass spectrometry of biopolymers. Anal Chem 63: 1193A- 1202A. doi: 10.1021/ac00024a716 [ Links ]
10. Hong X, Lu L, Xu D. 2016. Progress in research on acute hepatopancreatic necrosis disease (AHPND). Aquacult Int 24: 577-593. doi: 10.1007/s10499-015- 9948-x [ Links ]
11. Lane DJ. 1991. 16S/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M (eds). Nucleic acid techniques in bacterial systematics. Chichester: John Wiley & Sons. p 115-175. [ Links ]
12. Lee CT, Chen IT, Yang YT, Ko TP, Huang YT, Huang JY, Huang MF, et al. 2015. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc Natl Acad Sci USA 112: 10798-107803. doi: 10.1073/pnas.1503129112 [ Links ]
13. Nunan L, Lightner D, Pantoja C, Gomez-Jimenez S. 2014. Detection of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Dis Aquat Organ 111: 81-86. doi: 10.3354/dao02776 [ Links ]
14. Otta S, Praveena E, Bhuvaneswari T, Patil P. 2014. Early mortality syndrome (EMS)/acute hepatopancreatic necrosis disease (AHPND) – an update. In: Otta S, Poornima M, Patil P (eds). Training manual on health management practices for finfish and shellfish of brackish water environment. Chennai: Central Institute of Brakishwater Aquaculture, Indian Council of Agricultural Research. p 98-100. [ Links ]
15. Restrepo L, Bayot B, Betancourt I, Pinzón A. 2016. Draft genome sequence of pathogenic bacteria Vibrio parahaemolyticus strain Ba94C2, associated with acute hepatopancreatic necrosis disease isolate from South America. Genom Data 9: 143-144. doi: 10.1016/j.gdata.2016.08.008 [ Links ]
16. Shevchenko A, Wilm M, Vorm O, Mann M. 1996. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal Chem 68: 850-858. doi: 10.1021/ac950914h [ Links ]
17. Shevchenko A, Tomas H, Havli J, Olsen JV, Mann M. 2006. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat Protoc 1: 2856-2860. doi: 10.1038/nprot.2006.468 [ Links ]
18. Singhal N, Kumar M, Kanaujia PK, Virdi JS. 2015. MALDI-TOF mass spectrometry: an emerging technology for microbial identification and diagnosis. Front Microbiol 6: 791. doi: 10.3389/ fmicb.2015.00791 [ Links ]
19. Sirikharin R, Taengchaiyaphum S, Sanguanrut P, Chi TD, Mavichak R, Proespraiwong P, Nuangsaeng B, et al. 2015. Characterization and PCR detection of binary, pir-like toxins from Vibrio parahaemolyticus isolates that cause acute hepatopancreatic necrosis disease (AHPND) in shrimp. PLoS One 10: e0126987. doi: 10.1371/ journal.pone.0126987 [ Links ]
20. Soto-Rodríguez SA, Gomez-Gil B, Lozano- Olvera R, Betancourt-Lozano M, Morales-Covarrubias MS. 2015. Field and experimental evidence of Vibrio parahaemolyticus as the causative agent of acute hepatopancreatic necrosis disease of cultured shrimp (Litopenaeus vannamei) in Northwestern Mexico. Appl Environ Microbiol 81: 1689-1699. doi: 10.1128/AEM.03610-14 [ Links ]
21. Thitamadee S, Prachumwat A, Srisala J, Jaroenlak P, Salachan PV, Sritunyalucksana K, Flegel, TW, et al. 2016. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 452: 69-87. doi: 10.1016/ j.aquaculture.2015.10.028 [ Links ]
22. Tinwongger S, Proespraiwong P, Thawonsuwan J, Sriwanayos P, Kongkumnerd J, Chaweepack T, Mavichaket R, et al. 2014. Development of PCR diagnosis for shrimp acute hepatopancreatic necrosis disease (AHPND) strain of Vibrio parahaemolyticus. Fish Pathol 49: 159- 164. doi: 10.3147/jsfp.49.159 [ Links ]
23. Tran L, Nunan L, Redman RM, Mohney LL, Pantoja CR, Fitzsimmons K, Lightner DV. 2013. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis Aquat Organ 105: 45-55. doi:10.3354/dao02621 [ Links ]
24. Vorm O, Roepstorff P, Mann M. 1994. Improved resolution and very high sensitivity in MALDI TOF of matrix surfaces made by fast evaporation. Anal Chem 66: 3281-3287. doi: 10.1021/ ac00091a044 [ Links ]
Recibido: 15 de junio de 2017
Aceptado para publicación: 3 de noviembre de 2017

