










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista de Investigaciones Veterinarias del Perú
versão impressa ISSN 1609-9117
Rev. investig. vet. Perú vol.30 no.4 Lima out./dez. 2019
http://dx.doi.org/10.15381/rivep.v30i4.17188
ARTÍCULOS PRIMARIOS
Expresión de citoquinas Th1 (IL-2, IL-12, IFN-γ, TNF-α), Th2 (IL-4, IL-10, TGF-β) y Th17 (IL-17) en linfocitos circulantes de cuyes inoculados con una cepa de campo de Salmonella Typhimurium
Expression of cytokines Th1 (IL-2, IL-12, IFN-γ, TNF-α), Th2 (IL-4, IL-10, TGF-β) and Th17 (IL-17) in circulating lymphocytes of guinea pigs inoculated with a field strain of Salmonella Typhimurium
David Mejía1, Guillermo Salvatierra1, Jorge Maximiliano1, Rocío Rímac1, Dennis Carhuaricra1, Marcos Almeyda1, Luis Luna1, Raúl Rosadio1, Lenin Maturrano1,2,3
1 Laboratorio de Microbiología y Parasitología Veterinaria, Facultad de Medicina Veterinaria, Universidad Nacional Mayor de San Marcos, Lima, Perú
2 Laboratorio de Zootecnia y Producción Agropecuaria, Facultad de Medicina Veterinaria. Universidad Nacional Mayor de San Marcos, Lima, Perú
3 E-mail: amaturranoh@unmsm.edu.pe
RESUMEN
El objetivo del estudio fue evaluar la expresión relativa de las citoquinas involucradas en la respuesta inmune Th1 (IL-2, IL-12, IFN-γ, TNF-α), Th2 (IL-4, IL-10 y TGF-β) y Th17 (IL-17) en 21 cuyes (Cavia porcellus) inoculados experimentalmente con una cepa aislada de campo de Salmonella Typhimurium a una dosis de 102 UFC/ml, vía intraperitoneal, y comparados con un grupo control de siete cuyes (inoculados con una cepa tratada térmicamente). Se tomaron muestras de sangre los días 1, 3, 5, 7, 9, 15 y 30 pos-inoculación (p.i.). Se extrajo el ARN total de las células linfocitarias y se desarrolló la RT-PCR en tiempo real usando cebadores específicos para las citoquinas. La expresión relativa fue determinada por el método comparativo 2-ΔΔCt a fin de evaluar la expresión de los ARNm de las citoquinas respecto al calibrador (cuy sano), y usando como gen constitutivo de referencia al GAPDH como normalizador. Las expresiones de los genes de las citoquinas en el grupo Tratamiento mostraron un aumento con respecto al grupo Control y una cinética ascendente con respecto a los días p.i. Los primeros días hubo predominio de las expresiones Th1 sobre Th2, posteriormente ambos aumentaron, con predominio de IL-4 e IL-17 a partir del día 15 p.i. La inoculación de S. Typhimurium estimuló una expresión mayoritaria de IL-12 respecto a las otras citocinas, lo cual induciría en los cuyes una respuesta de tipo celular o Th1.
Palabras clave: cuy; Salmonella Typhimurium; respuesta inmune; citocinas
ABSTRACT
The aim of this study was to evaluate the relative expression of the cytokines involved in the immune response of Th1 (IL-2, IL-12, IFN-γ, TNF-α), Th2 (IL-4, IL-10 and TGF-β) and Th17 (IL-17) in 21 guinea pigs (Cavia porcellus) experimentally inoculated with a field isolate of Salmonella Typhimurium at a dose of 102 CFU/ml, intraperitoneally, and compared with a control group (inoculated with a thermally treated strain). Blood samples were taken on days 1, 3, 5, 7, 9, 15 and 30 post-inoculation (p.i.). Total RNA was extracted from lymphocyte cells and real-time RT-PCR was conducted using specific primers for each of the cytokines. The relative mRNA expressions were determined by the comparative method 2-ΔΔCt with respect to the calibrator (healthy guinea pig) and using GAPDH as a constitutive gene of reference as a normalizer. The expressions of the cytokine genes in the Treatment group showed an increase with respect to the Control group and an ascending kinetics throughout the study. In the first days there was a predominance of Th1 expressions over Th2, later both increased with a predominance of IL-4 and IL-17 from day 15 p.i. Inoculation of S. Typhimurium stimulated a major expression of IL-12 with respect to the other cytokines, which would induce a cellular or Th1 response in guinea pigs.
Key words: guinea pig; Salmonella Typhimurium; immune response; cytokines
INTRODUCCIÓN
El cuy (Cavia porcellus) es un mamífero roedor, usualmente criado en los hogares de las zonas rurales altoandinas (Bustamante, 1993). El cuy es un animal rústico, de ciclo de vida corto, por lo que puede ser criado a bajo costo (Ordoñez, 2003). Su crianza en granja se ha desarrollado en las últimas décadas, y se considera que la población en el Perú asciende a más de 12 millones (INEI, 2012).
La salmonelosis representa la principal amenaza a la producción de cuyes. Es una enfermedad con altos índices de mortalidad y morbilidad, principalmente en crías recién destetadas, ocasionando severas pérdidas económicas a los productores (Chauca, 1997; Matsuura et al., 2010). El principal agente etiológico de esta enfermedad es la bacteria Salmonella enterica, subespecie enterica serovar Typhimurium, la cual ocasiona una infección sistémica similar a la observada en humanos infectados con S. Typhi (Figueroa et al., 2005). La infección por Salmonella induce la generación de células T CD4+ y CD8+ específicas, y ambas poblaciones de células T son importantes para la protección durante las respuestas primarias y secundarias, aunque los mecanismos subyacentes a la protección mediada por las células T aún no se conocen por completo (Figueroa et al., 2005). La respuesta a S. Typhimurium implica inmunidad mediada por células T y B, y los mecanismos mediados por ambas poblaciones de linfocitos son importantes para el control de la infección primaria y la protección contra la infección secundaria (Mittrücker et al., 2002).
Haraga et al. (2008) explican el estudio de la biología e interacción con la célula huésped en la infección por Salmonella. La respuesta inmune incluye a los linfocitos T colaboradores: Th1 que son estimulados por IL12 y secretan IL-2 e interferón-γ. Los Th2 son estimulados por IL-1 y secretan IL-4, IL10 e IL-13, y los linfocitos Th17 son estimulados por IL-6, TGF-β e IL-23. Estos linfocitos secretan IL-17 y promuevn la inflamación mediada por neutrófilos (Tizard, 2009). La respuesta inmune generada por la infección de S. Typhimurium en cuyes es importante porque el conocimiento obtenido acerca de la estimulación inmunológica de determinadas poblaciones celulares ayudaría a generar vacunas efectivas. Es así que, el objetivo del presente estudio fue evaluar la respuesta inmune in vivo a través de la expresión de citoquinas de la respuesta inmune Th1, Th2 y Th17 en linfocitos sanguíneos, en cuyes desafiados con una cepa virulenta de Salmonella Typhimurium.
MATERIALES Y MÉTODOS
Lugar de Estudio
La crianza de los cuyes se realizó en las instalaciones del bioterio de la Facultad de Medicina Veterinaria (FMV) de la Universidad Nacional Mayor de San Marcos (UNMSM). La inoculación de Salmonella Typhimurium, las necropsias de los animales, el procesamiento de muestras, el aislamiento e identificación de cepas y los procedimientos de biología molecular se realizaron en el Laboratorio de Biología y Genética Molecular de la FMV-UNMSM durante la segunda mitad de 2016 y en 2017.
Material Experimental
Se emplearon como sujetos de estudio a 31 cuyes destetados entre 15 y 30 días de edad. Los cuyes fueron adquiridos del Programa de Investigación y Proyección Social en Animales Menores de la Universidad Nacional Agraria La Molina, Lima. Previo al desarrollo del experimento, los animales tuvieron un periodo de acondicionamiento en el bioterio de la FMV-UNMSM de 10 días. Los animales recibieron las mismas condiciones de crianza de su lugar de procedencia. Fueron distribuidos en dos grupos: Tratamiento (21 animales inoculados con la cepa virulenta) y Control (7 animales inoculados con la misma cepa, pero tratada térmicamente). El estudio fue aprobado por el Comité de Ética y Bienestar Animal de la FMVUNMSM mediante Constancia de Autorización N.° 2016-005.
Cepa Salmonella Typhimurium
Para la inoculación de los animales, se utilizó una cepa de S. Typhimurium procedente del cepario del laboratorio de FMVUNMSM (Unidad de Biología y Genética molecular), el cual se encontraba preservado en glicerol al 20% y almacenado a -80 °C. La cepa fue aislada de un cuy que murió con signos compatibles con salmonelosis clínica, procedente de una granja de crianza comercial en Pachacamac, Lima, en enero de 2016. El aislamiento e identificación de Salmonella fue realizado según los estándares de la norma ISO 6579 (2002). Adicionalmente, esa cepa fue confirmada mediante técnicas moleculares e identificación de los genes invA y fliC, según el protocolo de Jamshidi et al. (2010) estandarizado en el laboratorio. Para la reactivación de la cepa se procedió a colocar una alícuota en caldo APT (Agua Peptonada Tamponada) e incubado a 37 °C por 24 horas y confirmación mediante identificación de colonias sospechosas en agar XLD (Xilosa Lisina Deoxicolato). La cepa reactivada fue colocada en caldo TSA (Tripticasa Soya Agar) e incubada por 37 °C por 24 horas. Para la inoculación, el caldo conteniendo la cepa fue centrifugado a 5000 g por 10 minutos a 4 °C y diluido en PBS estéril hasta obtener una concentración de 102 UFC/ml (unidades formadoras de colonias), el cual fue determinado mediante espectrofotometría a 600 nm de longitud de onda.
Inoculación Experimental y Muestras
En el Día 0 (día de inicio del experimento) se sacrificaron tres animales escogidos al azar. Para esto, lo animales fueron previamente anestesiados usando una solución de ketamina, xilacina y atropina (Ket-A-Xyl®, Agrovet Market) a una dosis de 0.05-0.1 ml/cuy vía i.m. Una vez anestesiados, se extrajo 3 ml de sangre del corazón (por punción cardiaca), colocando las muestras en tubos con EDTA. Luego se procedió a la eutanasia mediante aplicación intracardiaca de 2 ml de pentobarbital sódico a 6.5 g (Halatal®, Montana). Este mismo procedimiento se hizo para el sacrificio de los demás cuyes durante los días de muestreo (días 1, 3, 5, 7, 9, 15 y 30).
Las muestras de sangre del Día 0 fueron utilizados como calibradores/ normalizadores para la PCR en tiempo real. Muestras de tejidos de los animales sacrificados para análisis histopatológico y de aislamiento de Salmonella sp, consistentes en hígado, vesícula biliar, nódulo linfático, pulmón, intestino delgado y cerebro fueron analizados con el fin de corroborar el suceso en la inoculación experimental. Las muestras de tejido para aislamiento de Salmonella sp fueron colocadas en caldo Rappaport-Vassiliadis por 18 horas a 37 °C y posteriormente sembradas en placas de agar XLD e incubadas a 37 °C por 24 horas. Las muestras de tejidos para histopatología fueron fijadas en formaldehido al 10% y teñidos con hematoxilina-eosina.
Se realizó la inoculación experimental a los 21 cuyes del grupo Tratamiento con 102 UFC/ml de S. Typhimurium suspendido en 1 ml de PBS por vía intraperitoneal (IP), según Panda et al. (2014) y a los 7 cuyes del grupo Control con la misma cepa tratada térmicamente (ebullición a 100 °C por 5 minutos). Se tomaron muestras de sangre y tejidos los días 1, 3, 5, 7, 9, 15 y 30 pos-inoculación (p.i.) experimental, usando los protocolos de anestesia y sacrificio previamente descritos.
ARN de Células Mononucleares Sanguíneas
Las células mononucleares de sangre periférica (sangre de los cuyes tomada del corazón) fueron obtenidas mediante separación por gradiente de Ficoll usando para ello Ficoll-PaqueTM (Sigma, USA), siguiendo las instrucciones del fabricante, para luego ser lavadas y diluidas en PBS 1X pH: 7.8. Las células mononucleares obtenidas fueron lisadas a través de choque térmico usando nitrógeno líquido y baño seco a 56 °C. El ARN de las células fue extraído usando el protocolo Trizol® Reagent (Thermo Fisher Scientific, USA). El ARN obtenido fue cuantificado utilizando el kit Qubit® RNA HS Assay Kits (Thermo Scientific). Para la lectura se utilizó el equipo de cuantificación Qubit® 3.0 fluorometer (Thermo Fisher Scientific).
PCR Tiempo Real
Para la síntesis de ADNc se utilizó el Revertaid RT Reverse Transcription Kit (Thermo Fisher Scientific), siguiendo las instrucciones del fabricante. La reacción en cadena de la polimerasa con transcriptasa inversa (RT PCR) en tiempo real fue realizada en un termociclador PikoReal 96 (Thermo Scientific) usando como fluoróforo Máxima SYBR Green. Se utilizaron los componentes y condiciones descritos en los cuadros 2 y 3. Se determinó el perfil citocínico Th1, Th2 y Th17 en las células mononucleares de los cuyes inoculados experimentalmente con S. Typhimurium utilizando los cebadores (primers) señalados en el Cuadro 1.
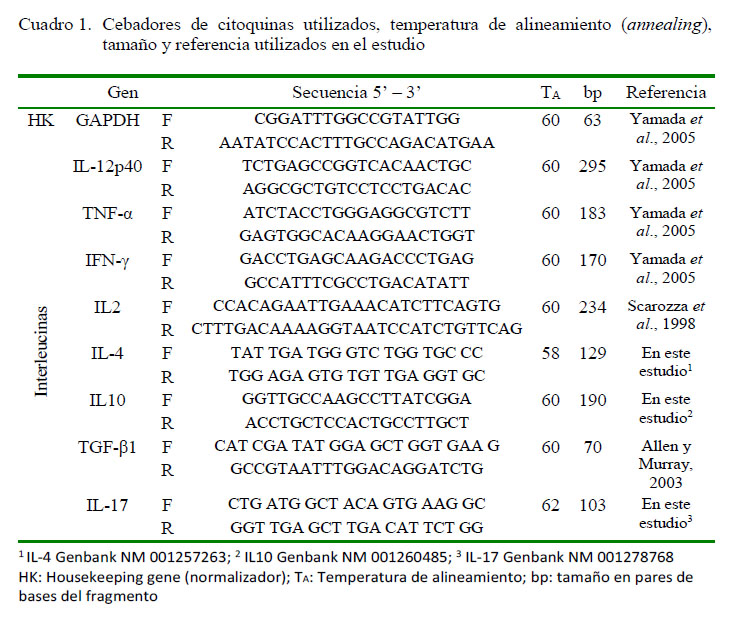
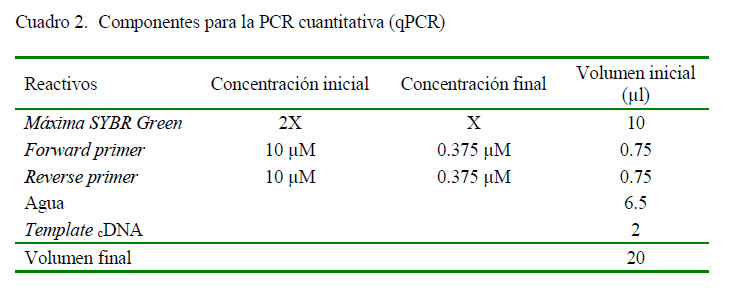
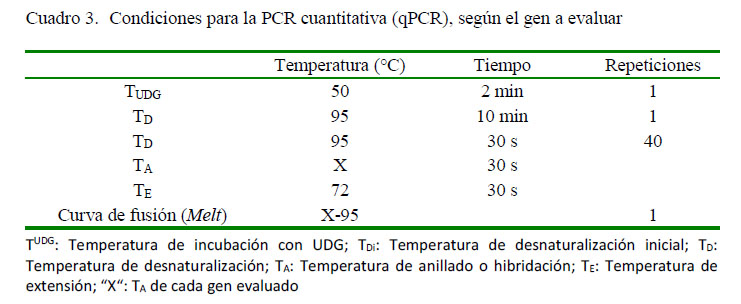
Cuantificación Relativa
Para el análisis de los niveles de expresión de las citoquinas se utilizó el método del Ct (threshold cycle) comparativo (Livak y Schmittgen, 2001), que permite comparar las curvas de amplificación de las citoquinas producidas. Esta aproximación sigue la siguiente fórmula: N= 2-ΔΔCt, donde N = Cantidad relativa de ARNm con respecto al calibrador y ÄÄCt = Diferencia entre el control endógeno y el ARNm a analizar con respecto al calibrador.
Análisis Estadístico
Los resultados se presentan en promedios obtenidos para cada citocina. Así mismo, se incluyen representaciones gráficas de las curvas mostrando la dinámica citocínica durante los 30 días del estudio.
RESULTADOS Y DISCUSIÓN
En el análisis histopatológico se encontraron lesiones en los órganos evaluados en todos los muestreos; lesiones que coinciden con lo observado en otros estudios (Layme et al., 2011) y que comprueban la infección por salmonelosis. Estas lesiones no fueron observadas en los cuyes del grupo Control. Por otro lado, fue posible recuperar la cepa inoculada en los tejidos analizados (hígado, vesícula biliar, contenido intestinal) en los individuos del grupo Tratamiento.
Se evaluó la cinética de la expresión del ARNm de las principales citocinas Th1, Th2 y Th17 mediante una técnica de PCR en tiempo real en cuyes inoculados con una cepa de Salmonella Typhimurium. Luego se cuantificaron los niveles de GADPH, IL-2, IL-4, IL-10, IL-12, IL-17, TNF-α, IFN-γ y TGF-β, detectándose en todos los animales, pero con una alta variabilidad entre estos.
Los resultados para GAPDH evidenciaron un aumento a partir del día 3 p.i. en todos los muestreos, porque es un gen de mantenimiento y es utilizado como gen endógeno en el PCR realizado. Esto demuestra que no hubo efecto citotóxico sobre los leucocitos y que su integridad y viabilidad son útiles para el análisis de expresión de otros genes de las citoquinas del estudio (Schmittgen y Livak, 2008).
La IL-12 en el primer día presentó una expresión de 75.44 veces más que el calibrador y que fue aumentando hasta el día 30 p.i. con un valor de 317.79 veces más. Los valores en el grupo control se mantuvieron con expresiones mínimas y fueron constantes durante el estudio (Figura 1A). IL-12 es producida principalmente por células presentadoras de antígeno, como los fagocitos y las células dendríticas en respuesta a la estimulación microbiana (Trinchieri, 2003). La IL-12 es considerada el factor clave para la respuesta inmune celular o Th1 (Germann et al., 1993). Los estudios en ratones con genes inactivados y el fenotipo de pacientes raros con mutaciones en el receptor para la IL-12 apoyan la conclusión de que la IL-12 es importante para la producción de IFNpor los linfocitos NK y los linfocitos T, y para la resistencia del huésped a las bacterias intracelulares y algunos virus (Abbas et al., 2012).
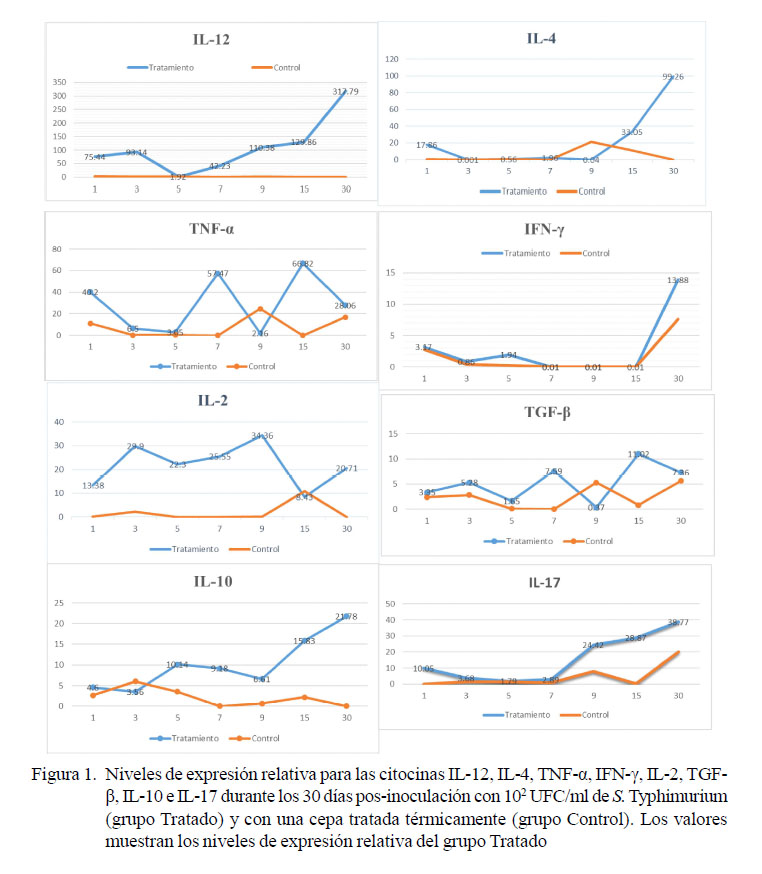
La IL-4 se expresó el primer día 17.86 veces más que el calibrador, para reducirse hasta el día 9 y posteriormente incrementar hasta 99.26 veces hacia el día 30 p.i. En el grupo control, los niveles de expresión se mantuvieron bajos, excepto el día 9 (21 veces más) (Figura 1B). El aumento de la expresión de IL-4 se asocia a una función de activación y multiplicación de linfocitos B productoras de inmunoglobulinas y de la diferenciación a células del perfil Th2 (células TCD4+) (Paul, 2010). Los ratones que albergan mutaciones en el gen IL-4(-/-) se infectaron con una variedad de S. Typhimurium HWSH (tipo salvaje), en comparación con los ratones IL-4(+/+). Los ratones IL-4(-/-) exhibieron un retraso en el tiempo de muerte después de la infección con S. Typhimurium HWSH (Everest et al., 1997). Grupos de ratones IL-4(+/+) infectados con S. Typhimurium HWSH purE (derivado menos virulento) mostraron muertes esporádicas y micro/macro abscesos alojados en sus tejidos, particularmente asociados con el hígado. Sin embargo, los ratones IL-4(-/-) infectados con dosis similares de la bacteria S. Typhimurium HWSH purE fueron resistentes a la muerte y no pudieron desarrollar abscesos detectables. La respuesta inmune tanto en ratones IL-4(-/-) como en IL-4(+/+) fue del tipo Th1 (Everest et al., 1997).
La TNF-α muestra una expresión en el primer día de 40.2 veces más en promedio que el calibrador. Se observó una alta variabilidad en la expresión de esta citocina entre los animales del grupo Tratamiento (Figura 1C). TNF-α es una citocina proinflamatoria que desempeña un papel importante en los mecanismos de defensa del huésped contra patógenos intracelulares. Se han descrito varios componentes bacterianos que pueden afectar los niveles de expresión de TNF-α (Vowels et al., 1995). Los principales componentes de Salmonella que pueden inducir la producción de TNF-α son el lipopolisacárido (LPS), flagelina y proteínas de membrana externa (porinas) (Galdiero et al., 1993, Ciacci-Woolwine et al., 1998). La expresión de esta citocina en casos de infección por Salmonella modula la expresión de ciertas proteínas y factores de virulencia en esta bacteria, resultando en una mayor capacidad de invasión y mayor activación de vías proinflamatorias, demostrando un posible papel de interacción de TNF-α del hospedero con Salmonella (Ma et al., 2010).
La expresión de IFN-γ se mantuvo bajo con relación a las otras citocinas, tanto en el grupo Tratamiento como en el grupo Control, alcanzando un pico de máxima expresión de 13.88 veces más que el calibrador en el grupo Tratamiento en el día 30 p.i. (Figura 1D). Esa citocina, producida principalmente por células T activadas y células NK (natural killer), es importante en la inmunidad contra patógenos intracelulares, como Salmonella enterica (Bao et al., 2000). En aves, S. Pullorum es capaz de inducir bajos niveles de IFN-λ en el bazo y tonsilas cecales de aves infectadas experimentalmente, sugiriendo un direccionamiento hacia una respuesta de tipo Th2, que promovería la infección persistente en estos animales (Tang et al., 2018). La persistencia de S. Typhimurium en cuyes (animales portadores) podría ser debido a este fenómeno.
La citocina IL-2 muestra una expresión relativa ascendente desde el día 1 p.i. hasta el día 3 (29.9 veces más que el calibrador), manteniéndose hasta el día 9 (34 veces más), disminuyendo hacia el día 15 para alcanzar el día 30 una expresión de 20.71 veces más, mientras que la expresión de esta citocina en el grupo Control se mantuvo en niveles muy bajos (Figura 1E). El incremento de esta citocina es suficiente para la maduración de linfocitos y su migración hacia el sitio de la inflamación, ya que se considera que es uno de los factores de crecimiento más importante de linfocitos T cuando son activados o estimulados por un antígeno, expandiendo su número significativamente (Malek et al., 2002). Ratones deficientes en IL-2 suelen tener una mortalidad del 50% entre la cuarta y sexta semana de edad a causa de anemia hemolítica autoinmune, mientras que los que sobreviven desarrollan enfermedad inflamatoria intestinal histológicamente similar a las lesiones observadas en colitis ulcerativa en humanos (Meijssen et al., 1998).
La citocina TGF-β mostró una baja expresión en comparación a las demás citocinas, alcanzando su máxima expresión el día 15 (11.02 veces más respecto al calibrador), pero con una gran variación en su expresión entre los individuos analizados. La expresión de esta citocina se mantuvo en niveles bajos en el grupo control (Figura 1F). La TGF-β es producida por varios tipos de células, incluyendo linfocitos B y T y macrófagos activados, siendo una citoquina multifuncional capaz de una variedad de efectos inmunológicos (Kehrl et al., 1986). Es importante en la progresión de las infecciones bacterianas y parasitarias, dado que se asocia con la inmunorregulación y el control de las actividades macrófagas. Diversos estudios in vitro sugieren que TGFβ tiene potentes efectos sobre la integridad de la mucosa gastrointestinal y la reparación de tejidos (Beck et al., 2003). Galdiero et al. (1999) encontraron que la administración in vivo de TGF-β en ratones infectados experimentalmente con S. Typhimurium indujo un aumento en la liberación de IFN-γ, demostrando que la expresión de TGF-β resulta crucial para la defensa en una infección por Salmonella en ratones.
La IL-10 mostró una expresion ascendente a lo largo del estudio, siendo alcanzado su máximo pico de expresión el día 30 (21.78 veces). El grupo Control, como en los otros casos, mantuvo una expresión baja (Figura 1G). Salazar et al. (2017) encontraron en ratones que no expresan esta interleucina (-IL-10) resistencia a la infección por S. Typhimurium en comparacion con individuos wild type (+IL-10), demostrando que se requiere la expresión de IL-10 para que S. Typhimurium produzca una infeccion sistémica. Concentraciones bajas de esta citocina como las que se observaron en este estudio favorecerían una respuesta de tipo celular, el cual es apropiado para combatir una infeccion por bacterias intracelulares, como S. Typhimurium y el direccionamiento a una respuesta de tipo humoral favorecería una infeccion sistémica por organismos intracelulares (Paradise et al., 2006).
La expresión de la IL-17 fue baja desde el principio en los animales del grupo Tratamiento, llegando a un pico de expresión de 38.77 veces más el día 30 p.i. mientras que en el grupo Control se expresó débilmente los días 9 y 30 (Figura 1H). Se ha demostrado que el IFN-γ y la IL-17 aumentan durante el periodo inicial de la enfermedad ocasionada por S. Typhimurium (Godinez et al., 2008). La respuesta inmune Th1 es conocida por ser esencial para el control de la infección por Salmonella en ratones y humanos. Rhee et al. (2005) demostraron que los ratones con deficiencia de IFN-γ son susceptibles a la infección por S. Typhimurium. Los macrófagos y las células dendríticas infectadas con S. Typhimurium son una fuente de citocinas, incluida IL-12, que estimula las células T en la mucosa intestinal para producir IFN-γ, IL17A e IL-22 (Godinez et al., 2009).
La interleucina 17 es una citocina proinflamatoria que se produce por las células T CD4+. Induce la diferenciación y la quimiotaxis de neutrófilos y contribuye a la eliminación de patógenos o induce enfermedades inflamatorias autoinmunes (InfanteDuarte et al., 2000). IL-17 induce no solo la migración de neutrófilos sino también la expresión de diversas moléculas antimicrobianas (Mayuzumi et al., 2010), incluyendo βdefensinas (BD), proteína S100A8/9, un quelante que elimina iones esenciales para la actividad bacteriana, y lipocalina-2, un inhibidor de la adquisición de hierro bacteriano (Liang et al., 2006; Godinez et al., 2008). En este estudio, se demuestra que IL-17 es producido pos-inoculación en cuyes. IL-17 ejerce múltiples funciones, tanto en la inmunidad adquirida como en la innata, produciendo citoquinas proinflamatorias que inducen la diferenciación y la migración de neutrófilos (Matsuzaki y Umemura, 2007; Ouyang et al., 2008).
En resumen, los cuyes inoculados con la cepa virulenta de S. Typhimurium mostraron una expresión mayoritaria de la interleucina 12 (IL-12), lo que estimularía una respuesta inmune de tipo Th1, siendo este tipo de respuesta la adecuada en casos de infecciones por organismos intracelulares como S. Typhimurium, quedando claro que la derivación de otro tipo de respuesta inmune como la humoral no es adecuada y puede conllevar a la exacerbación de la infección.
CONCLUSIONES
-
En ensayos in vivo, la administración de 102 UFC/ml de Salmonella Typhimurium, vía intraperitoneal, en cuyes incrementó la expresión de IL-2, IL-4, IL-10, IL-12, IL-17, TNF-α, IFN-γ y TGF-β en distintos grados.
-
La citocina IL-12 presentó la mayor expresión en comparación con las demás citocinas.
Agradecimientos
Al Programa de Innovación para la Competitividad y Productividad INNÓVATE Perú (FINCyT), fuente financiadora del proyecto «Desarrollo de una vacuna para el control y prevención de la Salmonelosis en la producción de Cuyes» Contrato N.° 362 PNICPPIAP 2014.
LITERATURA CITADA
Abbas A. Lichtman A. Pillai S. 2012. Inmunología celular y molecular. 7° ed. Barcelona, España: Elsevier. 546 p. [ Links ]
Allen SS, McMurray DN. 2003. Coordinate cytokine gene expression in vivo following induction of tuberculosis pleurisy in guinea pigs. Infect Immun 71: 4271-4277. doi: 10.1128/IAI.71.8.4271-4277.2003 [ Links ]
Bao S, Beagley KW, France MP, Shen J, Husband AJ. 2000. Interferongamma plays a critical role in intestinal immunity against Salmonella Typhimurium infection. Immunology 99: 464-472. doi: 10.1046/j.1365-2567.2000.-00955.x [ Links ]
Beck P, Rosenberg I, Xavier R, Koh T, Wong J, Podolsky D. 2003. Transforming growth factor-â mediates intestinal healing and susceptibility to injury in vitro and in vivo through epithelial cells. Am J Pathol 162: 597-608. doi: 10.1016/s0002-9440(10)63853-9 [ Links ]
Bustamante J. 1993. Producción de cuyes. Lima: Univ. Nacional Mayor de San Marcos. 259 p. [ Links ]
Chauca L. 1997. Producción de cuyes (Cavia porcellus). Roma: Organización de las Naciones Unidas para la Agricultura y la Alimentación – FAO. 120 p. [ Links ]
Ciacci-Woolwine F, Blomfield IC, Richardson SH, Mizel SB. 1998. Salmonella flagellin induces tumor necrosis factor alpha in a human promonocytic cell line. Infect Immun 66: 1127-1134. [ Links ]
Everest P1, Allen J, Papakonstantinopoulou A, Mastroeni P, Roberts M, Dougan G. 1997. Salmonella typhimurium infections in mice deficient in interleukin-4 production: role of IL-4 in infection-associated pathology. J Immunol 159: 1820-1827. [ Links ]
Figueroa OI, Verdugo RA. 2005. Mecanismos moleculares de patogenicidad de Salmonella sp. Rev Latinoam Microbiol 47: 25-42. [ Links ]
Galdiero F, de L’Ero CG, Benedetto N, Galdiero M, Tufano MA. 1993. Release of cytokines induced by Salmonella typhimurium porins. Infect Immun 61:155-161.
Galdiero M, Marcatili A, Cipollaro de L’Ero G, Nuzzo I, Bentivoglio C, Galdiero M, Romano Carratelli C. 1999. Effect of transforming growth factor â on experimental Salmonella Typhimurium infection in mice. Infect Immun 67: 1432-1438.
Germann T, Gately MK, Schoenhaut DS, Lohoff M, Mattner F, Fischer S, Jin SC, et al. 1993. Interleukin-12/T cell stimulating factor, a cytokine with multiple effects on T helper type 1 (Th1) but not on Th2 cells. Eur J Immunol 23: 1762-1770. doi: 10.1002/eji.1830230805 [ Links ]
Godinez I, Haneda T, Raffatellu M, George M, Paixao T, Rolán H, Santos R, et al. 2008. T cells help to amplify inflammatory responses induced by Salmonella enterica serotype Typhimurium in the intestinal mucosa. Infect Immun 76: 2008-2017. doi: 10.1128/IAI.01691-07 [ Links ]
Godinez I, Raffatellu M, Chu H. 2009. Interleukin-23 orchestrates mucosal responses to Salmonella enterica serotype Typhimurium in the intestine. Infect Immun 77: 387-398. doi: 10.1128/IAI.00933-08 [ Links ]
Haraga A, Ohlson MB, Miller SI. 2008. Salmonellae interplay with host cells. Nat Rev Microbiol 6: 53-66. doi: 10.1038/nrmicro1788 [ Links ]
Infante-Duarte C, Horton HF, Byrne MC, Kamradt T. 2000. Microbial lipopeptides induce the production of IL17 in Th cells. J Immunol 165: 6107-6115. doi: 10.4049/jimmunol.165.11.6107 [ Links ]
[INEI] Instituto Nacional de Estadística e Informática. 2012. IV Censo Nacional agropecuario 2012 Sistema de consulta de cuadros estadísticos, Perú. [Internet]. Disponible en: http://siea.minagri.gob.pe/siea/?q=iv-censonacional-agropecuario-2012. [ Links ]
ISO 6579. 2002. Microbiology General guidance on methods for the detection of Salmonella. Geneve, Switzerland: International Organization for Standardization. [Internet]. Available in: https://www.iso.org/standard/12985.html. [ Links ]
Jamshidi A, Kalidari G, Hedayati M. 2010. Isolation and identification of Salmonella Enteritidis and Salmonella Typhimurium from the eggs of retail stores in Mashhad, Iran using conventional culture method and multiplex PCR assay. J Food Safety 30: 558-568. doi: 10.1111/j.1745-4565.2010.00225.x [ Links ]
Kehrl JH, Roberts AB, Wakefield LM, Jakowlew S, Sporn MB, Fauci AS. 1986. Transforming growth factor-β is an important immunomodulatory protein for human B lymphocytes. J Immunol 137: 3855-3860. [ Links ]
Layme A, Perales R, Chavera C, Gavidia C, Calle S. 2011. Lesiones anatomopatológicas en cuyes (Cavia porcellus) con diagnóstico bacteriológico de Salmonella sp. Rev Inv Vet Perú 22: 369-376. [ Links ]
Liang SC, Tan XY, Luxenberg DP, Karim R, Dunussi-Joannopoulos K, Collins M, Fouser LA. 2006. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J Exp Med 203: 2271-2279. doi: 10.1084/jem.20061308 [ Links ]
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25: 402-408. doi: 10.1006/meth.2001.1262 [ Links ]
Ma J, Zhang Y, Xia Y, Sun J. 2010. The inflammatory cytokine tumor necrosis factor modulates the expression of Salmonella typhimurium effector proteins. J Inflamm 7: 42. doi: 10.1186/1476-9255-7-42. [ Links ]
Malek TR, Yu A, Vincek V, Scibelli P, Kong L. 2002. CD4 regulatory T cells prevent lethal autoimmunity in IL-2R deficient mice: implications for the nonredundant function of IL-2. Immunity 17: 167-178. doi: 10.1016/s10747613(02)00367-9 [ Links ]
Matsuura A, Morales S, Calle S, Ara M. 2010. Susceptibilidad a antibacterianos in vitro de Salmonella enterica aislada de cobayos de crianza familiarcomercial en la provincia de Carhuaz, Ancash. Rev Inv Vet Perú 21: 93-99. doi: 10.15381/rivep.v21i1.355 [ Links ]
Matsuzaki G, Umemura M. 2007. Interleukin-17 as an effector molecule of innate and acquired immunity against infections. Microbiol Immunol 51: 11391147. doi: 10.1111/j.1348-0421.2007.tb04008.x [ Links ]
Mayuzumi H, Inagaki-Ohara K, Uyttenhove C, Okamoto Y, Matsuzaki G. 2010. Interleukin-17A is required to suppress invasion of Salmonella enterica serovar Typhimurium to enteric mucosa. Immunology 131: 377-385. doi: 10.1111/j.1365-2567.2010.03310.x [ Links ]
Meijssen MAC, Brandwein SL, Reinecker HC, Bhan AK, Podolsky DK. 1998. Alteration of gene expression by intestinal epithelial cells precedes colitis in interleukin-2-deficient mice. Am J Physiol 274: 472-479. doi: 10.1152/ajpgi.1998.274.3.G472 [ Links ]
Mittrucker H, Kohler A, Kaufmann S. 2002. Characterization of the murine Tlymphocyte response to Salmonella enterica serovar Typhimurium infection. Infect Immun 70: 199-203. doi: 10.1128/iai.70.1.199-203.2002 [ Links ]
Ordoñez R. 2003. Plan de Introducción de la carne de cuy en Lima Metropolitana: Estudio de mercado y propuesta empresarial. Tesis de Maestría. Lima, Perú: Pontificia Universidad Católica del Perú. 213 p. [ Links ]
Ouyang W, Kolls JK, Zheng Y. 2008. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 28: 454-467. doi: 10.1016/j.immuni.2008.03.004 [ Links ]
Paul WE. 2010. What determines Th2 differentiation, in vitro and in vivo? Immunol Cell Biol 88: 236-239. doi: 10.1038/icb.2010.2 [ Links ]
Panda A, Tatarov I, Masek BJ, Hardick J, Crusan A, Wakefield T, Carroll K, et al. 2014. A rabbit model of non-typhoidal Salmonella bacteremia. Comp Immunol Microbiol Infect Dis 37: 211-220. doi: 10.1016/j.cimid.2014.05.004 [ Links ]
Paradise LJ, Friedman H, Bendinelli M. 2006. Opportunistic intracellular bacteria and immunity. Italia: Springer Science & Business Media. 322 p. [ Links ]
Rhee SJ, Walker WA, Cherayil BJ. 2005. Developmentally regulated intestinal expression of IFN-γ and its target genes and the age-specific response to enteric Salmonella infection. J Immunol 175: 1127-1136. doi: 10.4049/jimmunol.175.2.1127 [ Links ]
Salazar GA, Peñaloza HF, Pardo-Roa C, Schultz BM, Muñoz-Durango N, Gómez RS, Salazar FJ, et al. 2017. Interleukin-10 production by T and B cells is a key factor to promote systemic Salmonella enterica serovar Typhimurium infection in mice. Front Immunol 8: 889. doi: 10.3389/fimmu.2017.00889 [ Links ]
Scarozza AM, Ramsingh AI, Wicher V, Wicher K. 1998. Spontaneous cytokine gene expression in normal guinea pig blood and tissues. Cytokine 10: 851-859. doi: 10.1016/S1043-4666(98)90002-3 [ Links ]
Schmittgen T, Livak K. 2008. Analyzing real-time PCR data by comparative Ct method. Nature Protocols 3: 1101-1108. [ Links ]
Tang Y, Neil Foster, Michael A. Jones, Paul A. Barrow. 2018. Model of persistent Salmonella infection: Salmonella enterica serovar Pullorum modulates the immune response of the chicken from a Th17-type response towards a Th2-type response. Infect Immun 86(8): pii: e00307-18. doi: 10.1128/IAI.00307-18
Tizard IR. 2009. Inmunología veterinaria. 8° ed. Barcelona, España: Elsevier. 592 p. [ Links ]
Trinchieri G. 2003. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat Rev Immunol 3: 133-146. doi: 10.1038/nri1001 [ Links ]
Vowels BR, Yang S, Leyden JJ. 1995. Induction of proinflammatory cytokines by a soluble factor of Propionibacterium acnes: implications for chronic inflammatory acne. Infect Immun 63: 3158-3165. [ Links ]
Yamada H, Udagawa T, Mizuno S, Sugawara I. 2005. Newly designed primer sets available for evaluating various cytokines and iNOS mRNA expression in guinea pig lung tissues by RT-PCR. Exp Anim 54: 163-172. doi: 10.1538/expanim.54.163 [ Links ]
Recibido: 4 de marzo de 2019
Aceptado para publicación: 23 de octubre de 2019

