











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Las comunidades altoandinas asentadas por encima de los 3800 msnm tienen a la ga- nadería como la actividad más importante, y dentro de esta la crianza de los camélidos sudamericanos domésticos, no solo por su capacidad de adaptación a las difíciles condi- ciones medioambientales, sino por su empleo como una fuente alimenticia de proteína de origen animal y como un recurso para la pro- ducción de fibra de buena calidad (Fernández-Baca, 1991), en donde este recurso animal se suele distribuir generalmente en rebaños de 10 a 100 animales (Bravo y Huanca, 2012).
La mejora de la calidad genética de un rebaño justifica la aplicación de biotecnologías reproductivas como la superovulación y la transferencia de embriones, con el propósito de producir múltiples crías de hembras genéticamente superiores (Stewart, 2013). Sin embargo, a pesar de que el uso de protocolos de superovulación y que la tecnología de transferencia de embriones ha sido reporta- da en camélidos sudamericanos, aún existen deficientes resultados en la aplicación de la técnica de transferencia de embriones en estas especies, lo que ha sido relacionado con una alta variabilidad en la respuesta a los tra- tamientos con gonadotropinas durante la superestimulación y una baja tasa de recupe- ración de embriones (Ratto et al., 2013).
Se cree que esta variación es depen- diente del número de folículos que emergen durante el inicio de cada onda folicular, como sucede en rumiantes (Ireland et al., 2007), en donde la técnica de superovulación ha demostrado ser más efectiva en vacas con mayor número de folículos antrales al inicio de la superovulación (Singh et al., 2004). Los pequeños folículos antrales son el objetivo directo de los diversos protocolos con gonadotropinas y es precisamente la gran variabilidad individual en su número, lo que constituye el factor más limitante en el éxito de las biotecnologías de producción de embriones por superovulación en rumiantes (Ireland et al., 2007).
Múltiples estudios sobre el desarrollo de folículos ováricos en bovinos y dromedarios han demostrado la posibilidad de poder reali- zar una clasificación de los animales a partir del número de sus folículos emergentes, pre- vio al inicio de los procedimientos de superovulación; esto gracias a que se ha des- crito por ultrasonografía, que el número de folículos antrales durante la emergencia de las ondas foliculares representa una caracte- rística muy variable entre individuos distin- tos, pero altamente repetible dentro de un mismo animal (Burns et al., 2005; Manjunatha et al., 2012).
A partir de lo expuesto, el presente es- tudio tuvo por objetivo caracterizar y deter- minar el número de folículos emergentes en cada onda folicular en alpacas a fin de reali- zar una clasificación de los animales basada en el número de sus folículos emergentes.
MATERIALES Y MÉTODOS
El presente trabajo fue realizado duran- te enero y marzo del 2016 en el Centro de Investigación y Producción (CIP) Chuqui- bambilla, perteneciente a la Facultad de Me- dicina Veterinaria de la Universidad Nacio- nal del Altiplano. Se encuentra ubicado a 3950 msnm, en la región de Puno, Perú. Se em- plearon 34 alpacas Suri hembras adultas, va- cías, sin antecedentes de anormalidades reproductivas y con presencia de un folículo dominante >7 mm. Todas las alpacas en es- tudio fueron alimentadas con pastos natura- les, recibieron las mismas condiciones de manejo y se mantuvieron aisladas de los machos.
Las alpacas fueron distribuidas en dos grupos de similar número. Se procedió a divi- dir a los animales en dos grupos para reducir el estrés por manejo y manipulación. A todas las alpacas se les sincronizó la emergencia de una nueva onda folicular mediante la apli- cación intramuscular de 1 ml (4.2 µg) de acetato de buserelina (Conceptal, Intervet, Alemania) (Día 0). Los ovarios fueron escaneados mediante un ecógrafo transrectal (Aloka SSD-500, Japón) acoplado a un transductor lineal de 5 MHz. Todas las ecografías fueron realizadas cada dos días por un único operador por 60 días.
El estímulo hormonal (D = día cero) se aplicó para inducir la ovulación en los anima- les. Dos días después de su sincronización, se empezó un seguimiento ecográfico transrectal por individuo para determinar la posible ovulación, la cual fue establecida como la desaparición de un folículo dominan- te previamente registrado.
En ambos grupos de estudio, Todos los folículos >3 mm fueron medidos diametral- mente. El número, tamaño y posición de las estructuras ováricas observadas fueron graficadas en ovariogramas independientes. El día de la emergencia folicular fue conside- rado como el día en que el nuevo folículo do- minante presentó, retrospectivamente, un diá- metro >4 mm (Adams et al., 1990).
El número de folículos antrales (NFA) se definió como la cantidad total de folículos >3 mm de diámetro visibles por ultraso- nografía en ambos ovarios. Cada animal fue evaluado durante la emergencia de tres on- das foliculares consecutivas para determinar la repetibilidad del número de folículos antrales por alpaca. Así mismo, se evaluó el NFA >3 mm durante días aleatorios (días 12 y 24) del crecimiento folicular para compa- rarlos con el número de folículos antrales durante los días esperados de la emergencia folicular, según la metodología propuesta por Gobikrusthanth et al. (2017) en vacas. Los días fijos 12 y 24 fueron elegidos tomando como referencia el trabajo de Vaughan et al. (2004) en donde se reporta que el intervalo entre emergencias foliculares sucesivas (in- tervalo interonda) es de 12 días.
La normalidad del NFA y del intervalo interonda fueron analizados mediante la prue- ba de D'Agostino-Pearson. Se emplearon las pruebas de Mann-Whitney para identificar diferencias significativas en el NFA entre la emergencia folicular y un periodo aleatorio de crecimiento folicular. Se utilizó la prueba de Kruskal-Wallis para determinar diferen- cias significativas en el NFA >3 mm entre alpacas individuales, entre emergencias foliculares sucesivas y entre las categorías de clasificación por animal. La repetibilidad (rango 0-1; donde 1 es una repetibilidad per- fecta) fue calculada para el NFA >3 mm y el intervalo interonda. Se empleó el programa estadístico SPSS v. 24 (IBM, USA), y se consideraron que las diferencias observadas fueron estadísticamente significativas cuan- do p<0.05.
RESULTADOS
El diámetro promedio de los folículos preovulatorios >7 mm al inicio del estudio fue de 9.3 ± 1.79 mm (rango 7-14). La tasa de ovulación fue del 100% (34/34) luego de la aplicación de un análogo de la hormona liberadora de gonadotropinas (GnRH) para la sincronización de la onda folicular. El in- tervalo desde la administración del análogo de GnRH hasta el desarrollo de una emer- gencia folicular fue de 4.5 ± 1.4 días (rango 2-8). No se reportaron emergencias foliculares los días 9 y 10 pos-sincronización; sin embargo, se presentaron tres animales que tuvieron la emergencia de sus ondas foliculares hasta llegado el día 12 pos- sincronización.
En la Figura 1 se representa la distribu- ción del número de folículos antrales (NFA) >3 mm. El número promedio de folículos >3 mm no evidenció diferencias significativas entre las tres emergencias foliculares suce- sivas (Figura 2). Sin embargo, se determinó que el número promedio de folículos emer- gentes >3 mm difiere significativamente (p<0.05) entre alpacas, según la prueba no paramétrica de Kruskal-Wallis para muestras independientes.
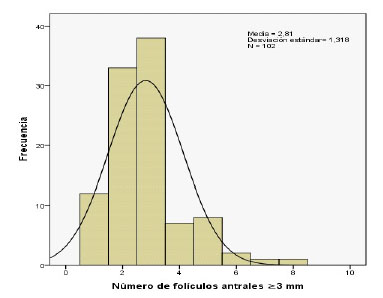
Figura 1 Distribución del número de folículos antrales >3 mm determinados durante la emergencia folicular en 34 alpacas
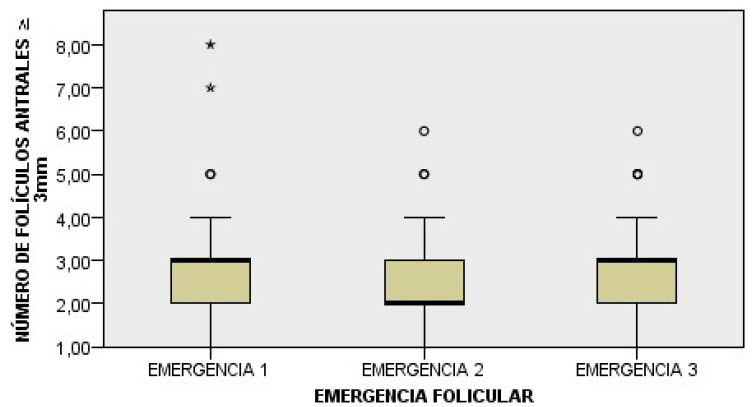
Figura 2 Diagrama de cajas del número promedio de folículos antrales >3 mm entre emergencias foliculares sucesivas en 34 alpacas
El NFA >3 mm determinados durante tres emergencias foliculares consecutivas y dos periodos aleatorios de crecimiento folicular (días fijos 12 y 24) en las 34 alpacas se pre- senta en el Cuadro 1. La repetibilidad del NFA >3 mm determinada durante las tres emer- gencias foliculares consecutivas fue de 0.72 (p<0.01) y durante dos periodos aleatorios de crecimiento folicular (días fijos 12 y 24) fue de 0.20 entre las mismas alpacas en es- tudio.
Cuadro 1 Folículos >3 mm durante tres emergencias foliculares consecutivas y dos periodos aleatorios (días 12 y 24) de crecimiento folicular en 34 alpacas
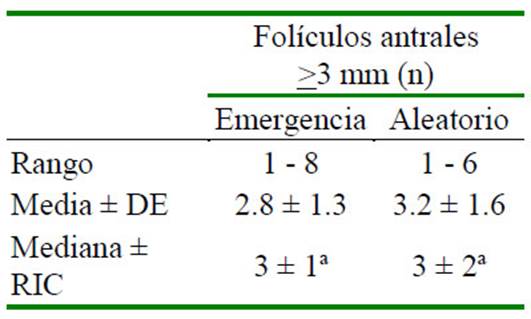
Letras distintas en una misma fila indican diferencia significativa (p<0.05)
DE: desviación estándar; RIC: rango intercuartil
En el Cuadro 2 se presenta la clasifica- ción de las 34 alpacas según el valor prome- dio del número de folículos antrales (NFA) >3 mm, donde las categorías propuestas fue- ron: alto (NFA >3), intermedio (3 >NFA > 2) y bajo (NFA <2).
Cuadro 2 Clasificación de las alpacas según el número promedio de folículos antrales (NFA) >3 mm

a,b,c Letras distintas en una misma columna indican diferencias significativas (p<0.05)
El intervalo interonda promedio (± des- viación estándar) fue del 16.9 ± 3.9 días, en donde los mayores porcentajes de intervalos fueron aquellos con 14 (16%), 16 (18%) y 18 (19%) días; inclusive registrándose interva- los de 20 a 24 días durante la evaluación de 68 ondas foliculares en 34 alpacas. Asimis- mo, se determinó que el intervalo interonda en alpacas presentó una repetibilidad de 0.23.
DISCUSIÓN
Este estudio pretendió caracterizar la emergencia folicular y determinar la repeti- bilidad de la onda folicular en alpacas en base al conteo del número de folículos emergen- tes. Para ello, se procedió a la sincronización de la emergencia de una nueva onda folicular mediante la aplicación parenteral de un aná- logo de GnRH; la cual permitiría, según Miragaya et al. (2006), estimular el surgimien- to de una nueva emergencia folicular sincronizada y el desarrollo de una fase lútea natural.
La tasa deovulaciónluegode laestimulación hormonal fue de 100% (34/34); similar a la descri- ta por Andrade (2007) y Huanca (2008) en alpacas. La detección del cuerpo lúteo a los 5.8 ± 1.4 días y el diámetro máximo prome- dio de 12.6 ± 1.5 mm fueron similares a los reportados en alpacas por Fernández-Baca et al. (1970) y Bravo et al. (1991).
Luego de la ovulación, el desarrollo de un cuerpo lúteo no interrumpió el surgimiento de una nueva emergencia folicular, confor- me a lo señalado por Ratto et al. (2003). Tampoco logró ejercer un efecto significati- vo sobre el número de folículos antrales >3 mm durante la primera emergencia folicular en comparación con las dos siguientes emer- gencias de las ondas foliculares; es decir, que a pesar del desarrollo luteal, el número de folículos emergentes >3 mm fue similar en las tres emergencias foliculares consecutivas.
El intervalo entre la sincronización has- ta la emergencia folicular fue de 4.5 ± 1.4 días, similar a intervalo de 4.3 ± 1.4 días re- portado por Andrade (2007) en alpacas. El retraso en la emergencia folicular hasta el día 12 después de la sincronización de tres animales pudo deberse a la presencia de folículos preovulatorios en fase de regresión previos a la sincronización que estarían luteinizándose, fenómeno descrito en llamas por Bravo et al. (1990).
Para el desarrollo del análisis se asumió que el número de folículos emergentes >3 mm determinados por ultrasonografía transrectal reflejan la reserva de folículos primordiales disponible en una alpaca adulta, tomando de referencia el estudio histológico sobre la re- serva ovárica realizado por Ireland et al. (2008) en vacas. En ese sentido, se pudo des- cribir que el número de folículos antrales >3 mm durante la emergencia folicular en alpacas se caracteriza por presentar una dis- tribución asimétrica en su histograma (Figura 1), donde la mayor parte de los animales presentaron un bajo número de folículos antrales (1 a 3 folículos). Según la prueba de D'Agostino-Pearson, se pudo verificar que el número de folículos emergentes >3 mm no sigue una distribución normal. Esta asimetría en la distribución de los folículos emergentes constituye un rasgo común en otras especies como los bovinos (Burns et al., 2005; Ireland et al., 2007; Shahzad et al., 2014; Gobikrushanth et al., 2017) y dromedarios (Manjunatha et al., 2012); y que, como tal, podría explicar en parte la respuesta indivi- dual diversa de estas especies a los distintos protocolos de superovulación ovárica descri- tos hasta la actualidad en alpacas.
Dado que el número de folículos antrales no presentó una distribución normal, la medi- da de tendencia central empleada debería ser la mediana en lugar de la media; sin embar- go, por fines comparativos con relación a otros estudios, ambos valores fueron calculados y presentados en el presente trabajo.
El promedio (± desviación estándar) del número de folículos emergentes con un diá- metro >3 mm fue de 2.8 ± 1.3. Sin embargo, hubo animales que presentaron solo un folí- culo durante las tres emergencias foliculares y otros que presentaron de cinco a más folículos emergentes. El rango de 1-8 folículos ilustra la gran variación para esta caracterís- tica en alpacas, hecho que fue descrito pre- viamente por diversos autores en bovinos (Burns et al., 2005; Ireland et al., 2007; Shahzad et al., 2014; Gobikrushanth et al., 2017) y dromedarios (Manjunatha et al., 2012).
En base al registro individual de datos, se identificó una notable similitud en el nú- mero promedio de folículos >3 mm durante las emergencias de las ondas foliculares sucesivas (p<0.05) (Figura 2); sin embargo, hubo una diferencia significativa en el núme- ro de folículos entre alpacas (p<0.05). Así mismo, el NFA >3 mm durante tres emer- gencias foliculares consecutivas y durante dos periodos aleatorios de crecimiento folicular (días fijos 12 y 24) no difirieron signifi- cativamente (Cuadro 1). Esto podría ser ex- plicado porque probablemente durante una etapa aleatoria de crecimiento folicular esté presente una población mixta de folículos en crecimiento y regresión (pequeños, medianos y grandes) lo cual contribuiría con un incre- mento en el NFA >3 mm y permitiría regis- trar de este modo un número de folículos si- milar al observado durante la emergencia de las ondas foliculares, en donde se espera que haya una población en crecimiento más funcionalmente homogénea (folículos peque- ños y medianos). Esta similitud en el NFA promedio durante estadios distintos del desa- rrollo folicular ha sido descrita por Gobikrushanth et al. (2017) en vacas.
La repetibilidad de 0.72 de NFA >3 mm puede considerarse como alta (Harper, 1994). Esta repetibilidad es similar a la reportada en otras especies como los bovinos (Burns et al., 2005; Ireland et al., 2007; Shahzad et al., 2014; Gobikrushanth et al., 2017) y drome- darios (Manjunatha et al., 2012). Además, se pudo identificar que esta repetibilidad va- lorada durante tres emergencias foliculares consecutivas es mayor que aquella determi- nada durante dos periodos aleatorios del cre- cimiento folicular (días fijos 12 y 24) (r=0.20); siendo esta menor repetibilidad comparable a la reportada en vacas Holstein (r=0.37) (Gobikrushanth et al., 2017). Estos resulta- dos sugieren que la evaluación ecográfica del número de folículos antrales >3 mm durante la emergencia de una onda folicular podría representar un criterio de selección de alpacas, como un paso previo al inicio de los protocolos de superovulación.
Mossa et al. (2012) encontraron una alta repetibilidad (0.95) del número de folículos antrales en vacas Holstein, siendo similar a la reportada por Manjunatha et al. (2012) en dromedarios (r=0.95) y por Shahzad et al. (2014) en búfalos (0.85). En un estudio para- lelo en vacas lecheras se identificó una mo- derada repetibilidad (0.58) en el conteo total de folículos emergentes luego de una abla- ción folicular (Boni et al., 1997), posiblemen- te debido a la falta de un seguimiento ecográfico de la onda folicular, ya que la eva- luación del número de folículos se limitó a un periodo fijo de 10 días luego de la ablación.
La repuesta a los protocolos de superovulación en alpacas son variables; así, Huanca (2008) obtuvo hasta 27 folículos preovulatorios, mientras que en el presente estudio fue de hasta 8 folículos. Esto podría indicar que en alpacas, el diámetro al cual los folículos emergentes se convertirían en de- pendientes de gonadotropinas (especialmen- te FSH) podria estar alrededor de los 2 mm, como sucede el ovino (McNeilly et al., 1991). En función a lo descrito por Shazad et al. (2014) en búfalos, las alpacas fueron clasifi- cadas con base al número de folículos emer- gentes en tres categorías, pudiéndose esti- mar que un 25% de las alpacas presentarían un número alto de folículos antrales.
El intervalo interonda de 16.9 ± 3.96 días; donde la mayoría de las ondas foliculares fue entre 14 y 18 días, fue similar al descrito por Vaughan et al. (2004) en donde se observaron un mayor número de intervalos interonda entre 12 y 16 días. Estas diferencias entre estudios podrían deberse, en parte, al mayor tamaño que exhiben los folículos dominantes en el presente estudio (9-13 mm); pues según Vaughan et al. (2004), el diámetro del folículo dominante se halla directamente correlacionado con el lar- go del intervalo interonda.
La repetibilidad de 0.23 del intervalo interonda puede indicar que esta caracterís- tica no repetible dentro de un mismo animal (Harper, 1994). Por otro lado, este valor fue menor que el r=0.88 reportado por Manjunatha et al. (2012) en dromedarios; observa- ciones que permitirían definir al intervalo interonda como una característica no previ- sible en alpacas.
CONCLUSIONES
El número de folículos antrales >3 mm durante la emergencia folicular es dis- tinto entre alpacas adultas (p<0.05).
El número de folículos antrales >3 mm durante la emergencia folicular es una característica repetible dentro de un mis- mo individuo (r = 0.72) (p<0.01).
El intervalo entre ondas foliculares representa una característica variable y no repetible (r= 0.23) en alpacas

