











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
La determinación de la productividad de la biomasa es fundamental para conocer la capacidad de almacenamiento de carbono de las plantas, ya que esta varía de acuerdo con el sitio ecológico (ShipleyyDion, 1992; Brown et al., 2004). En muchas especies de gramíneas nativas resulta complejo tomar medidas directas de la biomasa sin perturbar la mata, por lo que se estila el uso de modelos alométricos con las que se hacen inferencia de productividad (Yuen et al., 2016), y así poder inferir la cantidad de carbono que es retenido por el pastizal.
La cosecha de tallos, raíces y hojas es el estilo común para estimar la producción de biomasa; sin embargo, con la alometría solo se utiliza el área de las coronas y la altura de la mata para desarrollar modelos que tengan buena confianza de predicción en base al criterio de Akaike (Yang et al., 2017). La alometría es un método no destructivo que permite cuantificar y monitorear cambios ecológicos dentro de los ecotonos sucesionales dinámicos sensibles al cambio climático (Osland et al., 2014).
Los ecosistemas altoandinos tropicales sobre los 3500 msnm, donde las comunidades principales son los pajonales, carecen de descriptores de fisionomía y funcionamiento para un estudio ecológico eficiente. En este caso, los modelos alométricos pueden permitir obtener niveles de referencia de la captura de carbono para establecer políticas de manejo de pastizales, así como medir el impacto de cambios de uso de la tierra (Oliveras et al., 2014). La biomasa vegetativa está asociada a la biomasa reproductiva en muchas gramíneas altoandinas tropicales, por lo tanto, la medición alométrica permitirá, además dar un estimado de la capacidad reproductiva de muchas especies (Niu et al., 2009).
Las modelos alométricos tienen la necesidad del uso de gran cantidad de información para que tengan buena precisión, de allí que los modelos multiespecie son una buena alternativa en ecosistemas altoandinos tropicales que tienen escasa información (Nafus et al., 2009). Por lo tanto, el objetivo del presente estudio fue desarrollar un modelo alométrico multiespecie para estimar biomasa en Festuca dolichophylla (J. Presl), Festuca humilior (Nees & Meyen) y Calamagrostis vicugnarum (Weddell).
MATERIALES Y MÉTODOS
Área de Estudio
El estudio se realizó en el distrito de Canchayllo (Figura 1), área que corresponde a tundra húmeda, que presentó una temperatura ambiental promedio de 9 °C, velocidad de viento del N-E a 11 km/h, humedad ambiental 70% y precipitación total 680 mm. El suelo es del orden inceptisol, suborden ustepts, gran grupo calciustepts y subgrupo de lithic calciustepts; con pendientes entre l0 y 50%, textura franco arenoso y permeabilidad moderada. La capacidad de uso mayor del suelo comprendió a tierras aptas para pastos de baja calidad agrológica, asociado a tierras de protección limitada por suelo y erosión. La toma de muestras se realizó en una superficie de 5000 m2. El pastizal nativo tuvo un periodo de descanso (exclusión del pastoreo) de 12 meses. En el área de trabajo se instalaron seis transectos al paso de 50 metros lineales.
Material Vegetal
La Festuca dolichophylla (J. Presl) es de raíz adventicia fasciculada con rizomas no alargados, tallos de 50-90 cm de altura; lígula ciliolada de 1.8-2.7 mm, con extensiones laterales de 3 mm; láminas foliares involutas permanentemente enrolladas de 10-20 cm x 0.8-1.4 mm, panículas abiertas y laxas de 1220 x 3-7 cm, ramas inferiores 6-10 cm, espiguillas 12-13 mm, gluma inferior 4.5-5 mm, gluma superior 5.7-6.2 mm y lemas 6-7.2 mm. Es considerada como gramínea de porte alto (Tovar, 1993).
La Festuca humilior (Nees & Meyen) es de hojas basales laxas y pampilíneas, de 3-6 espiguillas, flores de glumas y lemas lanceoladas acuminadas desiguales, tallos erguidos y semierguidos de 20-35 cm, lígula de 0.5-0.7 mm, láminas foliares involutas o convolutas subuladas de 4-15 cm x 0.7-1.2 mm, panículas contraídas angostas con ramas adpersa o adperso-ascendentes de 4-7 cm. Es considerada como gramínea de porte mediano (Tovar, 1993).
La Calamagrostis vicugnarum (Weddell) Pilger es de láminas foliares involutas, casi filiformes de 2-4.5 cm x 0.5 mm, tallos de 5-25 cm, panículas 2-6 cm, glumas angostas hacia el ápice, lema cilidricas de 3.5-4 mm, raíces rizomatosas. Es considerada gramínea de porte bajo (Tovar, 1993).
Mediciones Alométricas
Las mediciones se realizaron al final del periodo lluvioso (abril-mayo), considerándose como especies claves del sitio de pastizal a las tres especies en estudio. Con una regla milimétrica se midió el diámetro basal estimando un promedio en dos medidas ortogonales (± 0.1 mm), tanto para las matas grandes como pequeñas. La altura (cm) de mata se midió desde el suelo hasta la canopia delimitada por la hoja bandera. Posterior a la medición se cosecharon las matas a 1 cm del suelo (incluida la mitad de la corona) y se llevaron al Laboratorio de Ecología y Utilización de Pastizales de la Universidad Nacional Agraria La Molina, Lima. Se les colocó en una estufa a 60 °C por 48 horas, previa eliminación del material senescente y luego se pesaron las muestras en una balanza digital (Ohaus®, Ranger 3000) con capacidad para 3000 g y precisión de 0.1 g, y finalmente se determinó la biomasa vegetal (kg MS).
Modelos Alométricos Generalizados
Se obtuvo una relación curvilínea (escala geométrica) de biomasa-altura utilizando transformaciones de logaritmo natural (ln). El modelo (lny)=a + b (lnx) se transformó en y = eaxb, una variación de la función de potencia (y=axb) para las relaciones alométricas (Andariese y Covington, 1986). La variable respuesta fue la biomasa de la mata: Y= biomasa del año 2016 al término de la época lluviosa [ln(g, mata-1)]. Las variables explicativas fueron el diámetro basal [XD, ln(cm)], diámetro canopia [XC, ln(cm)] y la altura de la planta [XH, ln(cm)].
Se utilizó una regresión simple por pasos (p<0.05) para ingresar el comando de ajuste del modelo en el software R v. 3.4, para predecir la biomasa en cada especie (especies individuales) y para todas las especies combinadas (multiespecies).
Se propusieron siete modelos de acuerdo con las variables explicativas, de los cuales se eligió uno mediante el principio de parsimonia y el criterio de Akaike (IAC). Se evaluó el efecto de una especie individual en los modelos multiespecie, al excluir sistemáticamente una especie del modelo y luego predecir la biomasa para las especies excluidas (w). En seguida, se comparó el R2 ajustado procedente del modelo multiespecies completo (tres especies). Las especies excluidas influyeron en el modelo multiespecies completo si R2 multiespecie not equal to R2 individual .
RESULTADOS
Variables Claves para los Modelos Alométricos
Los modelos propuestos analizados bajo el IAC indicaron que los modelos 5 y 7 fueron los más apropiados para proponer un modelo alométrico; sin embargo, bajo el principio de parsimonia el modelo apropiado es el siguiente: Y = 0.104 * XD 1.584 * XH 1.065 (Cuadro 1)
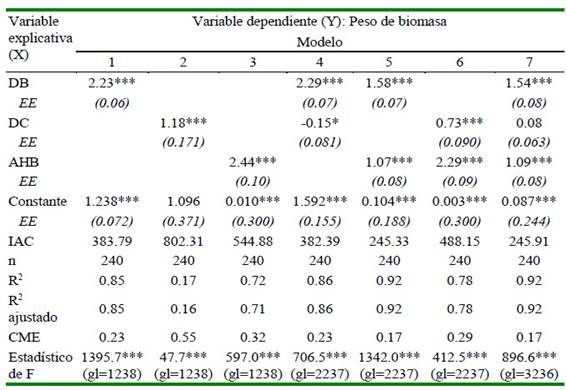
* p<0.1; ** p<0.05; *** p<0.01; EE: Error estándar; IAC: Criterio de información de Akaike; CME: Cuadrado medio del error
La variable de diámetro de canopia solamente explica el 16.3% del modelo, mientras que las variables diámetro basal y altura a la hoja bandera explican el 85.4 y 71.5%, respectivamente, de allí que pueden dar predictivos más precisos. El R2 ajustado del modelo, al adicionar la variable explicativa altura aumenta solo en 6.5%. Ante eso, es mejor quedarse con el modelo en función al diámetro basal, debido a su menor IAC y mayor R2. El mejor principio de parsimonia lo evidenció el modelo Y = 1.238 * XD 2.231.
Evaluación del Modelo Multiespecie
El Calamagrostis vicugnarum (Weddell) Pilger tiene un impacto al ser excluido del modelo multiespecie significativo evidenciado su menor R2 ajustado (6.7%) (Cuadro 2); sin embargo, no evidenció influencia cuando se le utilizó en los modelos predictivos con Festuca dolichophylla (J. Presl) y Festuca humilior (Nees & Meyen).
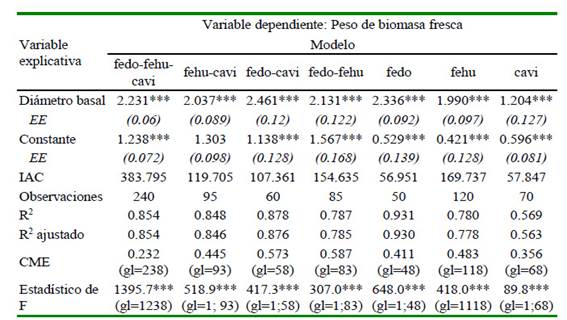
* p<0.1; ** p<0.05; *** p<0.01; EE: Error estándar; IAC: Criterio de información de Akaike; CME: Cuadrado medio del error
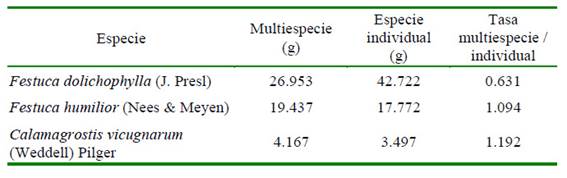
Cuadro 3 Relación de las predicciones de biomasa de los modelos de multiespecieespecies individuales
El coeficiente regresor disminuye cuando se excluye del modelo las mediciones de F. dolichophylla (J. Presl); sin embargo, el R2 ajustado es similar en ambos modelos. Por otro lado, el coeficiente regresor aumenta en el modelo donde se incluye F. dolichophylla (J. Presl) y C. vicugnarum (Weddell) Pilger. El coeficiente regresor es similar en el modelo multiespecie cuando se excluye C. vicugnarum (Weddell) Pilger (Cuadro 2).
Al excluir F. dolichophylla (J. Presl) y F. humilior (Nees & Meyen), la capacidad predictiva se comporta de manera similar que cuando están incluidos en el modelo; sin embargo, el comportamiento predictivo se sobreestima para C. vicugnarum (Weddell) Pilger.
El modelo individual de C. vicugnarum (Weddell) Pilger evidencia menor R2 ajustado (56.3%), mientras que F. dolichophylla (J. Presl) muestra un mayor R2 ajustado (93%), incluso mayor a los de multiespecie. El modelo individual de F. dolichophylla (J. Presl) tiene un regresor comparable con el de los modelos multiespecie, mientras que el modelo individual de C. vicugnarum (Weddell) Pilger muestra un regresor por debajo de los modelos multiespecie (Cuadro 2).
Predicción del Modelo Multiespecie en la Población Actual
La relación del promedio predicho en el modelo multiespecie con el modelo individual está por encima del 9-19% en F. humilior (Nees & Meyen) y C. vicugnarum (Weddell) Pilger; mientras en F. dolichophylla (J. Presl) está por debajo en 37%.
Los modelos de especie individual evidenciaron una ligera sobreestimación; sin embargo, el modelo multiespecie en F. humilior (Nees & Meyen) y C. vicugnarum (Weddell) Pilger evidencia sobreestimación. El regresor del modelo individual de C. vicugnarum (Weddell) Pilger muestra una pendiente distinta al modelo multiespecie; no obstante, la relación entre los estimados de los modelos es aceptable.
DISCUSIÓN
La fortaleza del modelo multiespecie propuesto para predecir la biomasa en gramíneas cespitosas comunes en pastizales altoandinos fue consistente con trabajos reportados para otras especies de plantas (Aguilar et al., 2012; Kebede y Soromessa, 2018). Los resultados son consistentes en cuanto al uso de información multiespecie e individuales de gramíneas nativas, donde también se evidenció que la altura mejora la fortaleza predictiva de los modelos (Andariese y Covington, 1986; Nafus et al., 2009). El modelo con la variable explicativa de diámetro basal presentó una precisión de 78.587.6%, que está dentro del rango 80-91% reportado (Nafus et al., 2009; Oliveras et al., 2014; Yang et al., 2017).
El peso de biomasa se relaciona a través de ecuaciones alométricas de potencia con medidas como diámetro basal, diámetro de canopia y altura a la hoja bandera en gramíneas, que a su vez estén asociados con el metabolismo ecológico en ecosistemas (Brown et al., 2004). El crecimiento alométrico de las diferentes características funcionales medidas en las gramíneas nativas está asociado a perturbaciones como el pastoreo y la estacionalidad climática, las cuales han sido tomadas en cuenta en el desarrollo de este modelo (Acosta-Gallo et al., 2011; Xiliang et al., 2016).
La transformación logarítmica de la biomasa en plantas ajusta más los modelos predictivos alométricos, ya que se evidencia una distribución log-normal de la variable dependiente. Las variables independientes pueden ser asignadas por componentes principales o un análisis stepwise (Chappell, 1989; Aguilar et al., 2012). Los modelos de este estudio coincidieron con una distribución lognormal y las variables explicativas se determinaron mediante stepwise.
Muchas de las especies de gramíneas nativas asocian su crecimiento de biomasa y producción de semillas a variables climáticas como la precipitación, siendo posible estudiar patrones alométricos para estimar la producción de semilla (Chen y Giladi, 2018). Las gramíneas altoandinas están fuertemente asociadas a la precipitación, coincidiendo su máximo crecimiento con la finalización de la época húmeda y la producción de semillas.
La captura de carbono y nitrógeno están asociadas con el desarrollo foliar en los árboles, siendo considerados bancos de almacenamiento de estos minerales en beneficio del cambio climático (Spinnler et al., 2003; McClaran et al., 2013; Paul et al., 2019). Los pajonales altoandinos son considerados bancos de almacenamiento de carbono y nitrógeno, por lo que el uso de las ecuaciones para la predicción de su volumen de almacenamiento es clave.
La plasticidad de muchas plantas es un inconveniente al predecir la biomasa mediante ecuaciones alométricas, ya que en especies poco rígidas es difícil desarrollar una ecuación predictiva. La plasticidad de las matas es una característica evolutiva que prioriza el crecimiento individual en desmedro del mantenimiento del sistema (Weiner, 2004). Las gramíneas nativas erectas son rígidas por lo que la estimación con ecuaciones alométricas es muy confiable, además que son especies adaptadas y mantienen el ecosistema altoandino.
CONCLUSIONES
El modelo alométrico multiespecie en el estudio de Festuca dolichophylla (J. Presl), Festuca humilior (Nees & Meyen) y Calamagrostis vicugnarum (Weddell) Pilger proponen al diámetro basal y altura a la hoja bandera como variables explicativas con fortaleza predictiva, siendo la altura a la hoja bandera la que evidenció un mejor comportamiento.
Los modelos alométricos multiespecie en función a diámetro basal permitió aumentar la fortaleza predictiva en las especies estudiadas; sin embargo, hay que considerar el comportamiento de la pendiente en Calamagrostis vicugnarum (Weddell) Pilger y la relación modelo multiespecie e individual en Festuca dolichophylla (J. Presl) cuando se desee estimar la biomasa a partir de dicha ecuación.

