











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Cebus albifrons es una especie de primate neotropical perteneciente a la Familia Cebidae, Subfamilia Cebinae. Esta especie es conocida bajo distintos nombres comunes tales como mico, macaco, mono blanco, maicero cariblanco y machín blanco entre otros en español y como White-fronted capuchin en inglés (Defler, 2010). Esta especie se caracteriza por ser de tamaño mediano, con una longitud corporal de 35.8 a 46.0 cm, y longitud de la cola de 40.1 a 47.5 cm. Los machos pesan de 3.2 a 3.5 kg, y las hembras de 2.3 a 2.9 kg. Usualmente de color marrón claro o leonado en el dorso y blanco cremoso en el vientre y alrededor del rostro (Defler, 2010; Fragaszy, 2004).
Los C. albifrons viven en grupos heterosexuales de 8 a 35 individuos (Defler, 1979, 2010). Si bien no se tiene reportes específicos para C. albifrons, las hembras del género Cebus alcanzan la madurez sexual a los 5 años, con gestación promedio 152 días, pariendo una cría cada 1-2 años (Fragaszy, 2004). Los individuos de la subfamilia Cebinae son omnívoros. Los depredadores más comunes de C. albifrons son los felinos, la boa, el águila arpía y el ser humano (Defler, 2010; Fragaszy, 2004). C. albifrons posee cerca de 11 subespecies distribuidas en Colombia, Venezuela, la región noroccidental de Perú, la cuenca amazónica superior de Brasil, el norte de Bolivia, el oeste de Ecuador y en la isla de Trinidad (Fragaszy, 2004; Rylands et al., 1995).
C. albifrons está categorizada en el Apéndice II de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES), en el cual se encuentran las especies que no necesariamente están en peligro de extinción, pero su comercio debe controlarse (CITES, 2017). Asimismo, C. albifrons se encuentra dentro de la categoría global de Preocupación Menor (LC) de la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (UICN), debido a su amplio hábitat, el cual permite presumir una alta población; sin embargo, actualmente la población global de esta especie presenta una tendencia a disminuir (Link et al., 2020). Sin embargo en Perú, C. albifrons no es considerada como una especie amenazada de acuerdo con el Decreto Supremo DS N.º 0042014-MINAGRI, que aprueba la actualización de la lista de clasificación y categorización de las especies amenazadas de fauna silvestre legalmente protegidas (MINAGRI, 2014)
El diagnóstico por imágenes es un método no invasivo, utilizado para observar estructuras biológicas y su función en vivo (Kiessling y Pichler, 2011). La ecografía o ultrasonografía es una herramienta diagnóstica que no solo permite observar la silueta de los órganos, sino que además permite observar su arquitectura interna (Barr, 2006). La ultrasonografía en animales se utiliza para determinar el sexo, evaluar el estado reproductivo y de diversas estructuras y órganos anormales, así como para realizar biopsias eco-guiada en órganos o lesiones específicas (Mannion, 2006); sin embargo, su uso es aún limitado en animales silvestres debido al escaso conocimiento de la topografía y anatomía ecográfica de los órganos de estas especies (Ribeiro et al, 2013).
Existen escasos estudios de la anatomía abdominal en primates neotropicales. Hill (1957) describió la anatomía gastrointestinal, puntualizando que el estómago posee una conformación periforme o en forma de J, el intestino es de gran longitud, y que el hígado está dividido en lóbulos central, izquierdo, derecho y caudal, siendo el lóbulo caudal de gran tamaño que cubre al riñón derecho. Mayor y López (2019) reportan que el hígado de C. albifrons está formado por los lóbulos lateral izquierdo, medial izquierdo, cuadrado, caudado (en el que se diferencian el proceso papilar y el proceso caudado), medial derecho y medial izquierdo. Asimismo, describen la posición del bazo, estomago, riñones, vejiga urinaria y testículos.
Existe una amplia documentación referida a la ultrasonografía en especies domésticas, como el perro y el gato, que permite comparar los hallazgos observados en otras especies (Mattoon y Nyland, 2004), pero hay una cantidad limitada de reportes de estudios ecográficos en animales silvestres (Ribeiro et al., 2013). La anatomía ecográfica de C. albifrons no se encuentra documentada; sin embargo, existen trabajos en especies de la misma familia, como en Cebus apella (Alves et al., 2007) y en Sapajus macrocephalus (Cano, 2016) donde se detallan las características de algunos órganos abdominales. El objetivo del presente estudio fue determinar las características ecográficas y dimensiones los órganos abdominales del machín blanco (Cebus albifrons), en cautiverio.
MATERIALES Y MÉTODOS
El presente estudio se realizó en el Parque Zoológico Huachipa, ubicado en el distrito de Ate Vitarte y en el Zoológico del Patronato del Parque de las Leyendas, ubicado en el distrito de San Miguel, ambos en la ciudad de Lima, Perú. La población objetivo fue la totalidad de monos machín blanco (Cebus albifrons) de ambos parques zoológicos que cumplieran con los siguientes criterios de inclusión: buen estado de salud, no haber recibido tratamientos ni haber cursado enfermedades durante los tres meses previos a las evaluaciones ecográficas, constatado mediante la revisión de la historia clínica y el examen físico.
Para la evaluación ecográfica, los animales fueron sometidos a ayuno por seis horas previas al manejo. Se realizó la contención física con una malla, para luego proceder a la contención química utilizando el protocolo de inmovilización establecido por cada zoológico. Este comprende el uso de clorhidrato de ketamina en dosis de 10 mg/kg IM, clorhidrato de xilazina en dosis de 0.5 mg/kg IM y clorhidrato de midazolam en dosis de 0.5 mg/kg IM.
Como preparación previa al examen ecográfico se colocó al animal sedado en decúbito dorsal, se humedeció las regiones abdominal y escrotal con alcohol al 70%, y se aplicó de manera uniforme el gel ecográfico. Los animales no fueron rasurados al tratarse de animales de la colección de exhibición. Se emplearon los equipos de ultrasonido Pie Medical modelo 100 Falco Vet® con transductor micro-convexo multifrecuencial de 5-7.5 MHz y el ESAOTE, modelo MyLab One Vet®, con el transductor microconvexo multifrecuencial SC3123, 4-9 MHz.
La evaluación comprendió los órganos abdominales, efectuándose cortes longitudinales, transversales y sagitales, según corresponda. Se anotaron las dimensiones y se evaluó la topografía, forma, ecogenicidad y ecotextura de cada órgano observado. Con los resultados de las medidas de las dimensiones de los órganos se realizó estadística descriptiva, empleando la media como medida de tendencia central y el desvío estándar y los valores límites como medidas de dispersión.
Para la descripción de la topografía se utilizó la metodología adaptada de Evans et al. (1991), que divide de manera imaginaria la superficie abdominal en tres zonas: abdomen craneal (epigastrio), abdomen medio (mesogastrio) y abdomen caudal (hipogastrio); y divide el abdomen en un total de nueve regiones: hipocondrio derecho, xifoidea, hipocondrio izquierdo, flanco derecho, umbilical, flanco izquierdo, inguinal derecha, púbica e inguinal izquierda (Figura 1).
RESULTADOS
El estudio comprendió a cuatro ejemplares machos adultos enteros del Parque Zoológico Huachipa y tres ejemplares del Zoológico del Patronato del Parque de las Leyendas, siendo un macho adulto castrado, una hembra adulta y un macho juvenil.
El protocolo de inmovilización química utilizado permitió realizar la evaluación ecográfica, logrando una buena inmovilización, minimizando el estrés y optimizando el procedimiento. Asimismo, el uso de ambos transductores permitió obtener imágenes adecuadas de los órganos evaluados. La posición decúbito dorsal facilitó la evaluación ultrasonográfica de los órganos, debido a la forma corporal de esta especie, aplanada ventro-dorsalmente. La aplicación previa de alcohol, para humedecer el pelaje, y de una capa uniforme de gel ultrasonográfico permitió una adecuada cohesión de transductor con la pared abdominal permitiendo una correcta visualización de los órganos evaluados.
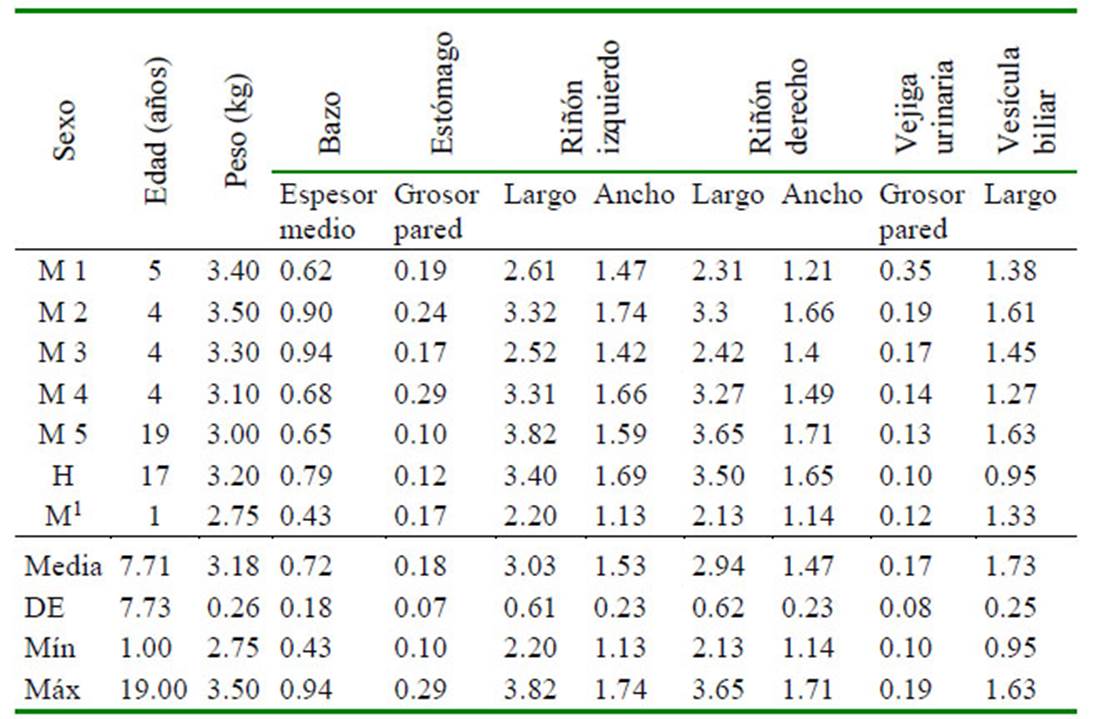
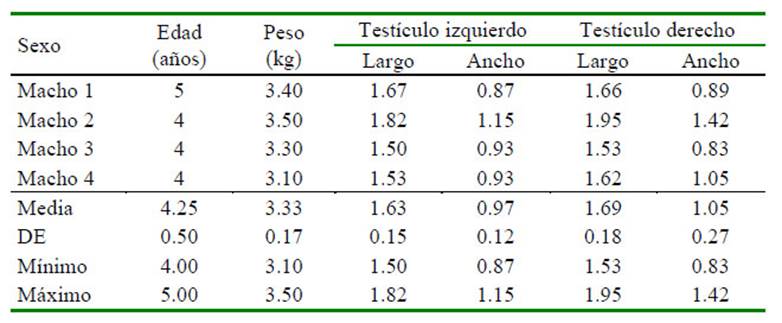
La reseña con las características de cada uno de los ejemplares y las dimensiones de los órganos abdominales se encuentran en el Cuadro 1, y el detalle de la medición de las dimensiones de los testículos de los machos adultos enteros se observa en el Cuadro 2. Las características ecográficas: topografía, márgenes, forma, ecotextura y ecogenicidad de los órganos evaluados se presentan en la Figura 1 y el Cuadro 3. La medición de las dimensiones de los órganos reproductivos del único ejemplar hembra correspondió a: ovario izquierdo de 1.99 x 1.50 cm, mostrando folículo ovárico de 1.08 cm de diámetro; el ovario derecho de 1.00 x 0.99 cm; y el útero de 2.79 cm de largo y 1.50 cm de diámetro.
El hígado se observó en el abdomen craneal, relacionado a la región xifoidea con predominio en el hipocondrio derecho. El borde craneal estaba asociado a la cúpula diafragmática y caudalmente con el estómago, en el lado derecho estuvo en contacto con el polo craneal del riñón derecho y en el lado izquierdo con el estómago y el bazo (Figura 1). El hígado presentó contornos regulares y lisos, con un parénquima de ecogenicidad y granularidad media, y ecotextura homogénea, siendo levemente hiperecogénica o isoecogénica comparada con la corteza renal; con presencia de tractos anecoicos cortos, correspondientes a vasos sanguíneos, ramas de las venas hepáticas y portales (Figura 2, Cuadro 3).
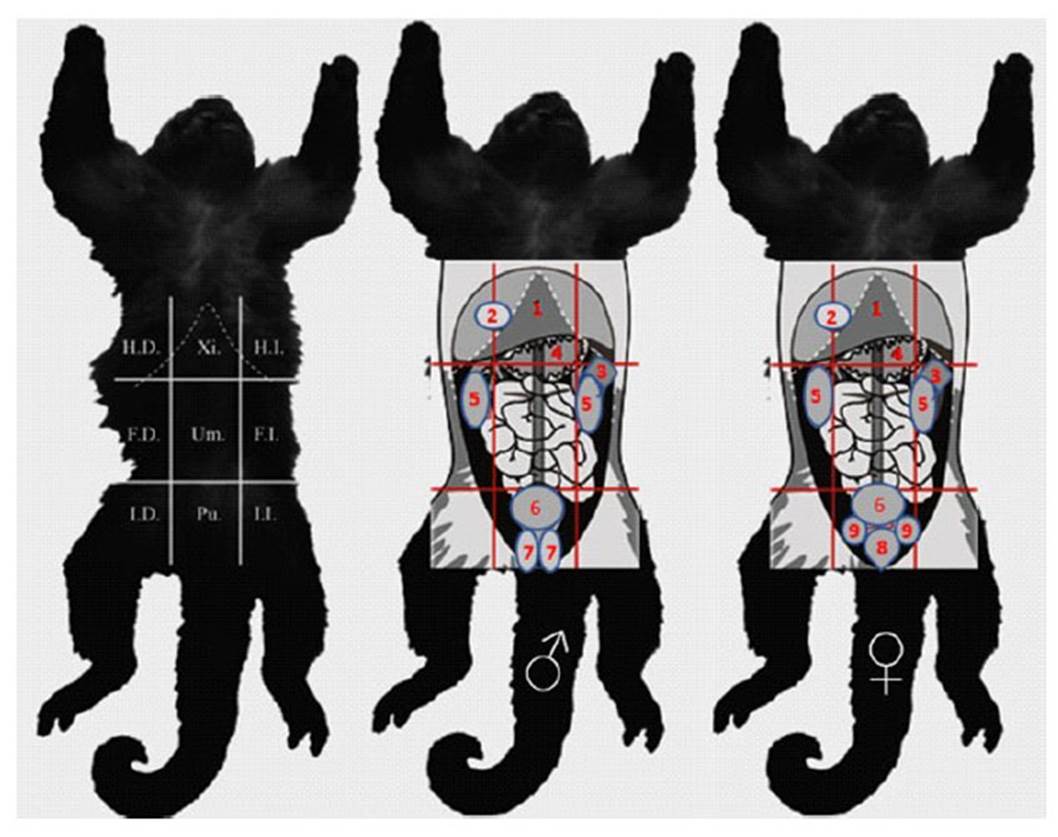
Figura 1 Esquema de las regiones topográficas y órganos del abdomen del machín blanco (Cebus albifrons): hipocondrio derecho (HD), xifoidea (Xi), hipocondrio izquierdo (HI), flanco derecho (FD), umbilical (Um), flanco izquierdo (FI), inguinal derecha, púbica (Pu) e inguinal izquierda (II); y la ubicación de los órganos en ellas, macho (B&) y hembra (@&): hígado (1), vesícula biliar (2), bazo (3), estómago (4), riñones (5), vejiga (6), testículos (7), útero (8) y ovarios (9). Imágenes adaptadas de Cano (2016)
La vesícula biliar fue de tamaño variable, dependiendo del contenido. En el estudio siempre se encontró con abundante contenido y ubicada entre los lóbulos hepáticos en el hipocondrio derecho (Figura 1); recubierta por una mucosa ecogénica regular delgada, de forma ovoide y con contenido anecoico (Figura 2, Cuadro 3).
El bazo se encontró en la región del flanco izquierdo, en posición craneal y medial con respecto al riñón izquierdo (Figura 1). Presentó márgenes lisos y bien definidos, conformando una línea hiperecogénica delgada que corresponde a la cápsula. El bazo se observó de forma alargada con la cabeza ligeramente más ancha, de ubicación craneal; con un parénquima ecogénico, moderadamente hiperecogénico al parénquima hepático y corteza renal, de granularidad fina y densa (Figura 3, Cuadro 3).
Cuadro 1 Dimensiones (en centímetros) de los órganos abdominales evaluados mediante ultrasonografía de 07 ejemplares machín blanco (Cebus albifrons) en cautiverio en el Parque Zoológico Huachipa y Zoológico del Patronato del Parque de la Leyendas (Lima)
1 Macho juvenil
Cuadro 2 Dimensiones (cm) de los testículos de machos evaluados mediante ultrasonografía de cuatro ejemplares machos adultos enteros de machín blanco (Cebus albifrons) en cautiverio en el Parque Zoológico Huachipa (Lima, Perú)
Cuadro 3 Descripción ecográfica de los órganos abdominales y escrotales del machín blanco (Cebus albifrons) en cautiverio en el Parque Zoológico Huachipa y en el Parque de las Leyendas (Lima, Perú)
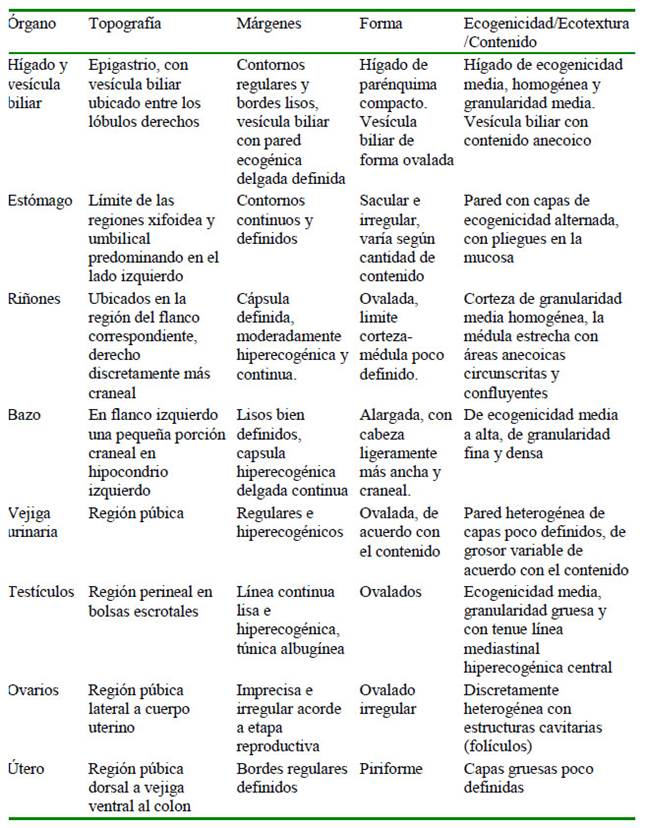
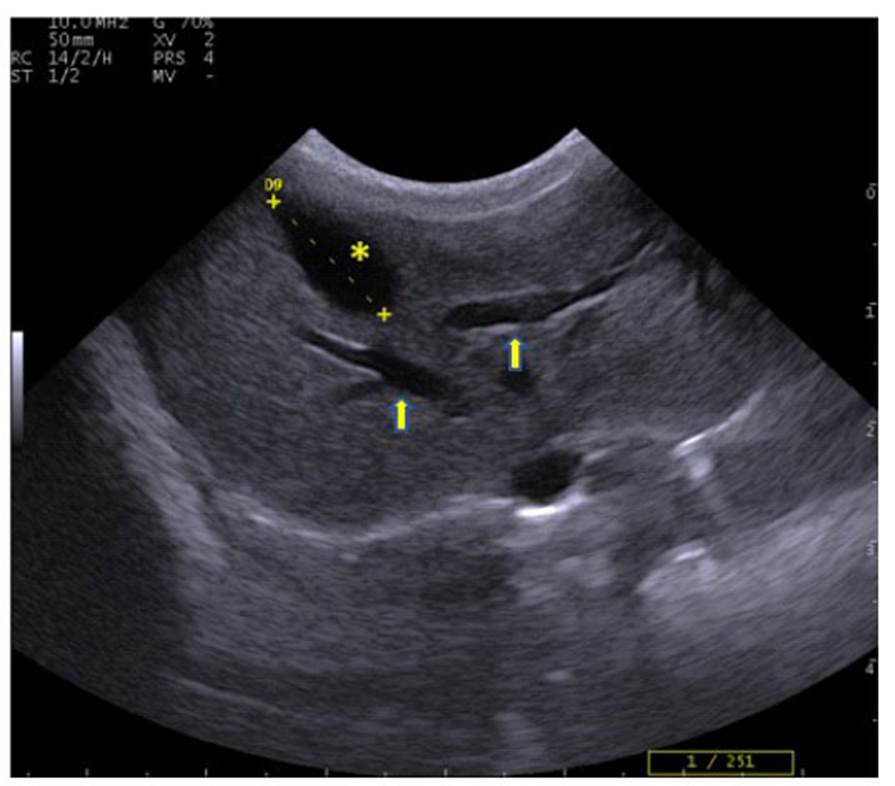
Figura 2 Imagen ecográfica de machín blanco (Cebus albifrons) mostrando la vesícula biliar (*), de 1.33 cm de longitud, y el parénquima hepático mostrando tractos anecoicos, vasos sanguíneos venosos hepáticos (()
El estómago se ubicó en el límite de la región xifoidea y la parte craneal de la región umbilical (Figura 1). Se encontró escaso contenido gástrico y poca cantidad de gases debido al ayuno previo realizado. La pared se mostró como una estructura delgada, curvilínea con bordes bien definidos, distinguiéndose las capas de la pared que se suceden con una ecogenicidad alterna, siendo la capa interna levemente ondulante, correspondientes en a los pliegues gástricos (Figura 3, Cuadro 3).
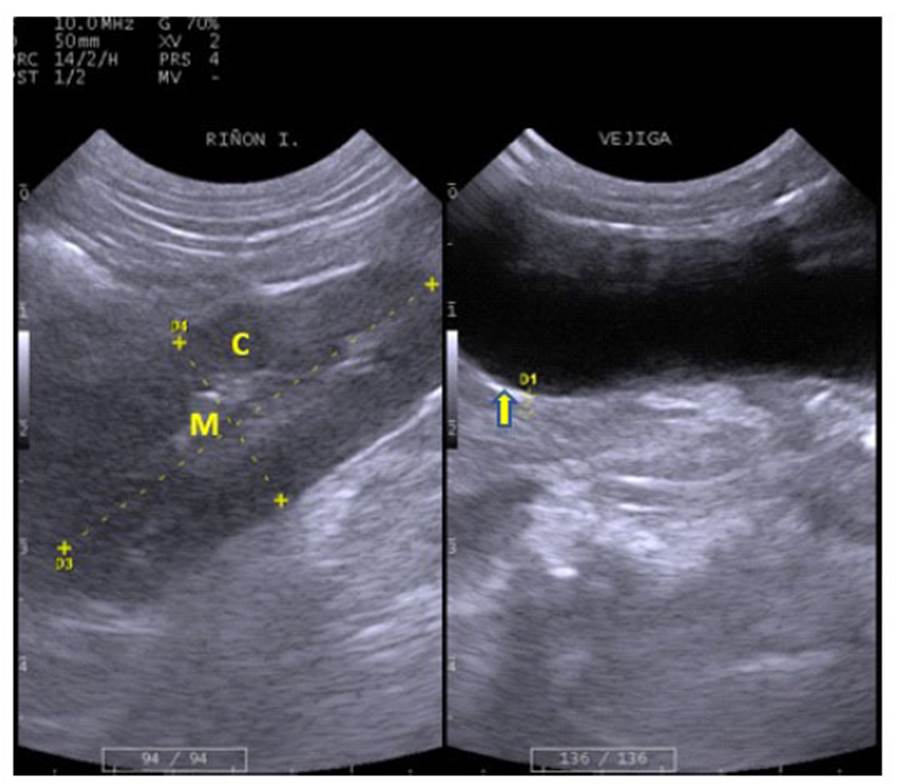
Los riñones, órgano par, presentaron forma ovalada. Se ubicaron lateralmente en los flancos derecho e izquierdo, respectivamente. El riñón derecho se encontró en posición ligeramente más craneal con respecto al izquierdo (Figura 1). La cápsula renal se observó levemente hiperecogénica y bien definida; en tanto que la corteza renal se presentó hipo ecogénica o isoecogénica en relación con el parénquima hepático; con una granularidad media homogénea dispersa y la médula renal estrecha observándose como áreas discretas anecoicas circunscritas confluyentes (Figura 4, Cuadro 3).
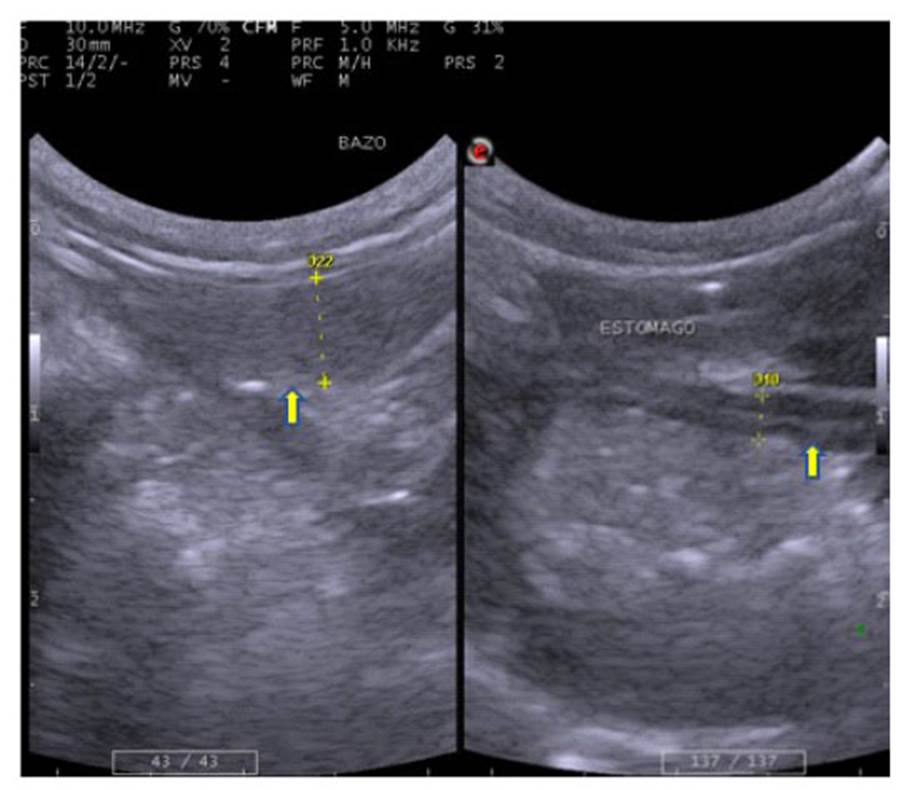
Figura 3 Imágenes ecográficas de machín blanco (Cebus albifrons), a la izquierda mostrando el bazo (() de 0.52 cm de espesor a nivel del cuerpo esplénico; a la derecha muestra el estómago con contenido de ingesta moderado, con pared de 0.24 cm de espesor (()
La vejiga urinaria se encontró en la región púbica. Se encontró con contenido moderado a abundante en todos los especímenes, lo que facilitó el examen de la cavidad abdominal caudal, ya que sirvió como ventana acústica. Presentó forma redondeada, y el tamaño varió de acuerdo con el contenido de orina; sus bordes fueron regulares, hiperecogénicos y bien definidos; pared conformada por capas poco definidas (Figura 4, Cuadro 3).
El útero se ubicó en región púbica, dorsal a vejiga y ventral al colon (Figura 1). Presentó aspecto piriforme con capas gruesas poco diferenciadas. Los ovarios se ubicaron laterales al útero (Figura 1), presentando una forma ovalada irregular discretamente heterogénea, pudiendo observarse estructuras cavitarias compatible con folículos ovárico (Figura 5, Cuadro 3).
Los testículos, órgano par, se ubicaron en la región perineal, en las bolsas escrotales (Figura 1). Presentaron forma redondeada y limitada por una línea continua lisa e hiperecogénica, correspondiente a la túnica albugínea. El parénquima presentó una ecogenicidad media y una granularidad gruesa y con una línea mediastinal hiperecogénica central tenue, mediatinun testis (Figura 6, Cuadro 3).
DISCUSIÓN
La preparación previa al examen ecográfico es determinante para una eficiente visualización de los órganos abdominales. Se recomienda que el paciente esté en ayunas al menos 12 horas antes de una evaluación ecográfica, debido a que la comida y gas presentes en el estómago no permiten la correcta evaluación de su luz y pueden ocultar estructuras cercanas (Mannion, 2006). Asimismo, se recomienda que el paciente sea rasurado antes del examen ecográfico para la obtención eficiente de imágenes ecográficas, lo cual puede constituir un problema en animales silvestres. En este caso se obvió este paso, ya que la eliminación de pelo en zonas amplias puede predisponer al paciente a hipotermia (Redrobe, 2006), de modo que el estudio ecográfico en este estudio se realizó directamente sobre el pelo humedecido con alcohol y utilizando una capa uniforme de gel para eliminar el aire que pueda quedar atrapado entre la cubierta pilosa y el transductor.
La contención química con sedación es de esencial importancia para realizar exámenes ecográficos en animales silvestres (Cortopassi, 2004), para evitar posibles complicaciones por el estrés del manejo. El protocolo de anestesia convencionales en primates incluye fármacos anestésicos disociativos y agonistas alfa-2 adrenérgicos (Carpenter, 2012); sin embargo, el protocolo empleado por ambas instituciones adicionó clorhidrato de midazolam (benzodiacepina), permitiendo realizar la evaluación ecográfica de manera exitosa, obteniéndose imágenes de manera óptima y sin causar estrés innecesario a los especímenes.
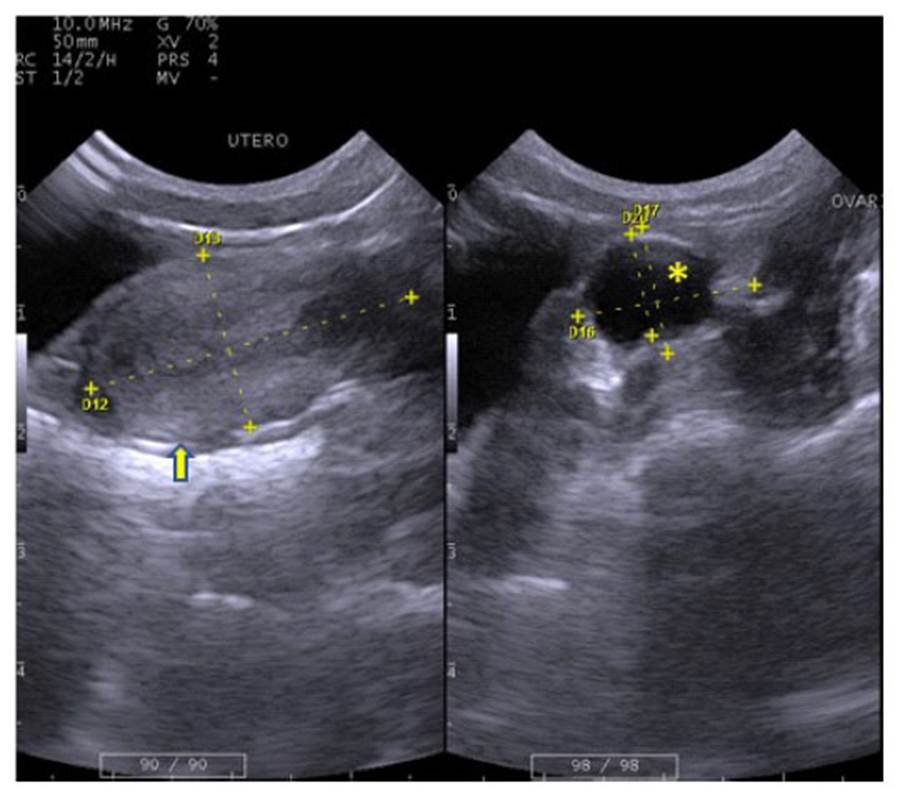
Figura 5 Imágenes ecográficas de machín blanco (Cebus albifrons) mostrando a la izquierda el útero ((), de 2.79 de longitud y 1.50 cm de diámetro, y a la derecha el ovario izquierdo, de 1.99 x 1.50 cm, mostrando folículo ovárico de 1.08 cm de diámetro (*)
En un estudio realizado en Titi común (Callithrix jacchus) se describió la evaluación ecográfica con el uso de una frecuencia mínima de 7.5 MHz para obtener imágenes con una resolución adecuada (Wagner y Kirberger, 2005). En el presente estudió el rango de frecuencia de los transductores empleados (4 a 9 MHz) fueron adecuados para la evaluación de los órganos abdominales del machín blanco, que supera en tamaño y peso (4.0 kg) al de los monos Titi (250 g).
Debido a la escasas información sobre la ecografía en esta especie los resultados obtenidos son comparados con información obtenida de otras especies de la misma familia: mono Machín negro o Capuchino cabeza dura, Cebus apella (Alves et al., 2007) y del Machín o Capuchino cabeza grande, Sapajus macrocephalus (Cano, 2016); así como con especímenes de otra familia como el Titi común Callithrix jacchus (Wagner y Kirberger, 2005) y de mamíferos de especies domésticas, donde existe descripciones ultrasonográficas más detalladas, como en perros de talla pequeña (<4 kg) Canis lupus familiaris y el gato Felis silvestriss catus (Mannion, 2006; Mattoon y Nyland, 2004).
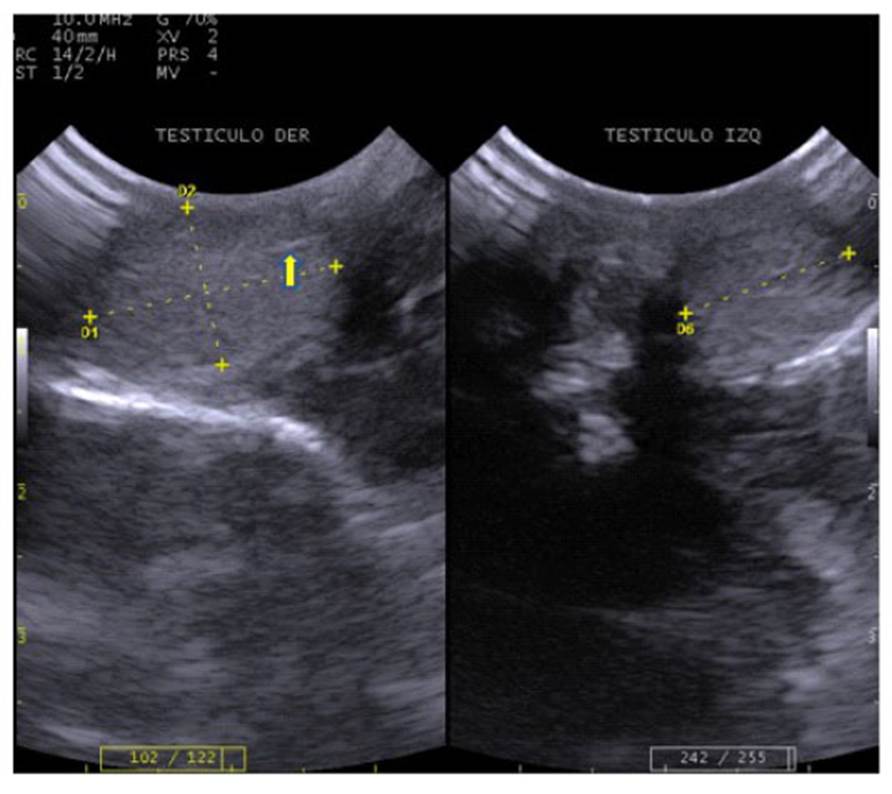
Figura 6 Imágenes ecográficas de machín blanco (Cebus albifrons) mostrando a la izquierda corte longitudinal de testículo derecho de 1.73 x 1.11 cm, evidenciando discreta línea mediastinal ((); y a la derecha corte transversal de testículo izquierdo de 1.20 cm
Al igual que en otros estudios en primates neotropicales, C. albifrons presenta el hígado en la región xifoidea con predominio en el hipocondrio derecho, en íntimo contacto con el diafragma; con características ecográficas similares a las descritas para C. jacchus por Wagner y Kirberger (2005), C. apella por Alves et al. (2007), S. macrocephalus por Cano (2016) y mamíferos domésticos, perros y gatos, por Mattoon y Nyland (2004). Asimismo, al igual que en estas especies, no se pudo distinguir la separación de los lóbulos hepáticos.
La vesícula biliar se observó como una estructura piriforme anecoica, evidenciando considerable contenido líquido, debido al periodo de ayuno Su ubicación, hacia la derecha del abdomen craneal, se observó similar a lo descrito en C. apella por Alves et al. (2007), S. macrocephalus por Cano (2016) y caninos (Mattoon y Nyland, 2004). La vesícula biliar de C. albifrons difiere de la de C. jacchus (Wagner y Kirberger, 2005) y felinos (Mattoon y Nyland, 2004) en que la vesícula presentó una apariencia interna bilobulada, e incluso multilobulada, debido a la disposición plegada del órgano, dando este aspecto.
La ubicación del bazo en el flanco izquierdo, en posición craneal y medial con respecto al riñón izquierdo es similar a lo reportado en S. macrocephalus (Cano, 2016), y perros y gatos (Mannion, 2006; Mattoon y Nyland, 2004). En este estudio ecográfico se observó al bazo de una forma alargada, con la cabeza ligeramente más ancha; forma de «lengua» también descrita por Mayor y López (2019) en su atlas anatómico, y coincide con lo reportado en caninos y felinos (Mannion, 2006; Mattoon y Nyland, 2004). En cambio, en estudios en S. macrocephalus se observa que el bazo de esa especie tendría una aparente forma triangular (Cano, 2016). El parénquima del bazo de C. albifrons muestra características ecográficas muy similares a las reportadas en S. macrocephalus, canino y felinos, con una textura granular fina, densa y homogénea, con un nivel de ecogenicidad mayor en comparación al hígado y la corteza renal (Cano, 2016; Mannion, 2006; Mattoon y Nyland, 2004).
Los estómagos evaluados en este estudio presentaron escaso contenido gástrico, cuyas características fueron muy similares a las descritas para S. macrocephalus (Cano, 2016), C. jacchus (Wagner y Kirberger, 2005) y para el perro y gato doméstico (Mattoon y Nyland, 2004; Mannion, 2006). La pared del estómago de C. albifrons cuenta con cuatro capas distinguibles, las cuales son alternadamente hiperecogénicas e hipogénicas. En el estudio en S. macrocephalus también se pudo identificar con precisión las cuatro capas de la pared gástrica, siendo el grosor de la pared de dimensiones similares a las descritas en este estudio.
Si bien Mayor y López (2019) describen el riñón de C. albifrons como liso y multilobulado, con una médula renal subdividida en segmentos de forma piramidal o pirámides renales bien diferenciadas, estos detalles no son distinguibles ecográficamente. A la evaluación ecográfica se pudo observar que las pirámides renales no fusionadas se distinguen como áreas anecoicas circunscritas confluyentes en una médula renal estrecha. Asimismo, las imágenes ecográficas permitieron observar que los riñones presentaron forma ovalada y con una diferenciación poco precisa entre corteza y médula y una médula estrecha, también similar a lo reportado para las otras especies de primates neotropicales como C. apella y S. macrocephalus (Alves et al., 2007; Cano, 2016); excepto para C. jacchus, donde la diferenciación cortico-medular fue limitada (Wagner y Kirberger, 2005). Sin embargo, esta descripción de la forma anatómica del riñón de C. albifrons difiere con lo reportado por Mayor y López (2019), que los describen en forma de frejol similar a lo descrito en el riñón del perro y gato (Mannion, 2006; Mattoon y Nyland, 2004). Esto pudo deberse al corte ecográfico obtenido, que difiere a los observado entre las especies aplanadas ventro-dorsalmente (primates) de los aplanados latero-lateralmente (caninos y felinos domésticos).
La cápsula renal se observó levemente hiperecogénica y bien definida como se reporta en otras especies de primates neotropicales (Alves et al., 2007; Cano, 2016; Wagner y Kirberger, 2005). La corteza renal en C. albifrons, así como en C. apella, S. macrocephalus, perros y gatos, se presentó hipoecogénica en relación con el hígado y el bazo (Alves et al., 2007; Cano, 2016); a diferencia de lo reportado en C. jacchus, donde la corteza renal fue hiperecogénica respecto a estos órganos (Wagner y Kirberger, 2005).
La forma y el espesor de la pared de la vejiga urinaria del C. albifrons fue similar al descrito en C. apella (Alves et al., 2007); siendo mayor al espesor en S. macrocephalus (Cano, 2016). El tamaño de la vejiga urinaria varió de acuerdo con el contenido de orina, no diferenciándose las capas en contraste con lo reportado en el coatí (Nasua nasua) donde se pudieron diferenciar claramente su estratificación (Ribeiro et al., 2013).
La ubicación del cuerpo uterino, en la región púbica, la morfología mostrando un cuerpo del útero sin presencia de cuernos, y las dimensiones son similares a los descritos en C. apella por Alves et al. (2007) y S. macrocephalus por Cano (2016). Los testículos y ovarios no fueron descritos en trabajos previos de Alves et al. (2007) o Cano (2016), debido a que en el primer estudio solo se trabajó con hembras sin ubicar los ovarios, y en el segundo solo se trabajó con machos castrados; por lo que este estudio constituye la primera descripción de las gónadas en primates neotropicales de la Subfamilia Cebinae, observándose que comparten características generales de forma y características ecográficas descritas en caninos y felinos (Mattoon y Nyland, 2004; Mannion, 2006).
Se puede concluir que las características ecográficas y las dimensiones de los órganos abdominales de C. albifrons son similares a las descritas para otras especies de la Subfamilia Cebinae, caninos de tamaño pequeño y felinos domésticos. Los ovarios y testículos fueron descritos por primera vez en especímenes de la Subfamilia Cebinae y muestran similitud con lo descrito en caninos de tamaño pequeño y felinos domésticos.

