











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
Aunque la finura de la fibra es la principal característica que define la calidad de la lana y fibra en los diferentes procesos de producción y transformación, la presencia y el porcentaje de medulación también son importantes (Gupta et al., 1981; McGregor, 2006), porque determinan su uso en la industria e influyen en el grado de calidad de la fibra, hilos y tejidos, afectando también su apariencia y rendimiento.
Las fibras meduladas se caracterizan por tener un canal central (médula) conteniendo residuos celulares y bolsas de aire que se di- rigen en forma continua o fragmentada a lo largo de la fibra (Gupta et al., 1981; Hunter, 1993; Lupton y Pfeiffer, 1998; McGregor et al., 2013); no obstante, resulta de particular interés las fibras fuertemente meduladas (fibras objetables o tipo kemp), donde la médula es mayor de 60% del diámetro de la fibra (ASTM, 1986).
Las fibras meduladas, principalmente las fibras objetables, constituyen un defecto desde el punto de vista textil, debido al efecto sobre las propiedades del teñido en los tejidos, pues las bolsas de aire presentes en la médula les dan una apariencia más clara que las fibras normales, por afectar las propiedades ópticas de la luz que pasa a través de la fibra por difracción y no por la diferencia de consumo de tinte por la queratina de la fibra (McGregor, 2002). También son menos resistentes, aun- que más elásticas, y al parecer están asociados a un mayor grosor y pérdida de rizos (Gupta et al., 1981; Moore, 2015), por lo que tienen efectos adversos sobre la suavidad, vellosidad, rigidez y el picor de hilos y telas, siendo así que muchos consideran a las fibras meduladas como contaminantes de los vellones de lana, mohair, cachemira, alpaca y llama (Hunter, 1993; Balasingam, 2005; Frank et al., 2009; McGregor, 2012; Cruz et al., 2019).
A través del tiempo se han realizado enormes esfuerzos para disminuir la MDF de las fibras de lana y alpacas. En el caso de Perú, las alpacas llegan a tener medias de diámetro de fibra entre 20 y 23 µm (Montes et al., 2008; Quispe et al., 2009; Vásquez et al., 2015, Omachea et al., 2015; Pinares et al., 2018; Cruz et al., 2019), que a la clasificación se encuentran dentro de la calidad Alpaca Baby (INDECOPI, 2014). Los programas de mejoramiento genético no consideran el porcentaje de medulación de las fibras; sin embargo, al tener una heredabilidad moderada, posibilita su uso como criterio de selección, lo que permitiría disminuir el efecto del factor de picazón ocasionado principalmente por fibras fuertemente meduladas (Pinares et al., 2018; Cruz et al., 2019), pero también se lograría afinar aún más la fibra. En este sentido, McGregor (2006) y Onal et al. (2007) refieren que la correlación entre medulación y diámetro de la fibra, en alpacas, llamas y conejos entre otros, es positiva con valores entre moderado a alto.
La medulación no siempre resulta ser un defecto, pues la presencia de médula torna las fibras más livianas, razón por la cual, tejidos en base a fibras de conejo y alpaca son más livianas que los de lana (Czaplicki, 2012) y brindan un mejor aislamiento térmico (Moore, 2015), debido a la presencia de bolsas de aire que amortigua el intercambio térmico (Czaplicki, 2012). Por tanto, siendo las fibras objetables la causa del problema del picazón de fibras de alpacas, resulta de importancia reducir o eliminar este tipo de fibras, además de las que tienen medulación continua (Pinares et al., 2018).
La incidencia de medulación en lanas de ovinos Merino es muy baja, en fibras de mohair y cachemira es de baja a moderada, en fibras del buey almizclero es moderada (McGregor, 2012), en fibras de llamas es generalmente alta (Martínez et al., 1997; Cochi, 1999; Córdova, 2015; Poma, 2018), en fibras de alpacas varía entre regular y alta (McGregor, 2006; Lupton et al., 2006; Contreras, 2010; Ferguson et al., 2012; Pinares et al., 2018, 2019; Radzik et al., 2018; Cruz et al., 2019), mientras que en pelos de conejo Angora es extremadamente alta (McGregor, 2012) a pesar de que la finura varía entre 10 a 15 µm (Chattopadhyay et al., 2005), demostrándose que la incidencia de medulación se incrementa con la edad (Martínez et al., 1997; Lupton et al., 2006, Pinares et al., 2019)
Investigaciones sobre medulación se han realizado con bastante profundidad en lanas y fibras de mohair, utilizando el OFDA 100 y el microscopio de proyección (Balasingam, 2005); sin embargo, en fibras de otras especies existe limitada información al respecto, ya que su determinación mediante el microscopio de proyección resulta bastante laborioso, necesita mucho entrenamiento y los resultados varían de persona a persona (Balasingam, 2005; Shakyawar et al., 2013), mientras que utilizando el OFDA 100 se obtienen resultados dentro de un rango bastante amplio (Pinares et al., 2019) y en algunos casos discordantes con la realidad, pues al parecer los umbrales de opacidad debieran ser redefinidos para su uso en otros tipos de fibra, diferentes al de la lana o mohair. En esta línea, Rafat et al. (2007) y Pinares et al. (2018) muestran resultados discordantes de medulación medidos con el microscopio de proyección y el OFDA 100. Por tanto, los reportes sobre tasa de medulación en alpacas y llamas, realizados por McGregor (2006), Lupton et al. (2006), Ferguson et al. (2012) y Cruz et al. (2019, entre otros, deberían ser discutidos.
Estas razones condujeron a realizar el presente trabajo de investigación con la finalidad de evaluar la incidencia y relación entre los diferentes tipos de medulación (no meduladas, con medulación continua, fragmentada, discontinua y fuertemente meduladas), así como su relación con la finura de las fibras de diferentes especies: ovino, alpaca, llama, paco-vicuña y conejo Angora, utilizando mediciones directas y objetivas.
MATERIALES Y MÉTODOS
Las muestras de lana de ovinos Merino (n=15) y paco vicuña (n=15) fueron obtenidos de dos granjas de Missouri, EEUU, de hembras de variada edad; las muestras de fibras de conejos Angora (n=15) se obtuvieron de granjas de pequeños criadores del distrito de Chota, Perú; y las muestras de llamas (n=15) y alpacas (n=15) de machos y hembras adultas (2-7 años) fueron obtenidos del Centro de Investigación y Producción Lachocc, perteneciente a la Universidad Nacional de Huancavelica. Adicionalmente, 140 muestras de fibras de alpacas tuis machos de color blanco fueron obtenidos de granjas de la región Puno, Perú.
Las muestras de fibras de lana de ovino, conejos, alpacas, llamas y paco vicuñas (75 muestras) fueron analizadas en el Laboratorio de Fibras Textiles de la Universidad Nacional Autónoma de Chota (Perú); mientras que, las 140 muestras de fibras de alpacas tuis machos fueron analizadas en el Laboratorio de Fibras de la Universidad Nacional de Juliaca. Para el análisis se utilizó el equipo denominado Medulómetro, que evalúa la calidad de la fibra animal, teniendo en cuenta el tipo de medulación (tipo kemp, continua, discontinua, fragmentada y no medulada). El Medulómetro es un microscopio de proyección modificado, al que se incorporaron dos motores y una tarjeta electrónica para el escaneo automatizado, conectado a una computadora que tiene instalado un software propietario (Figura 1).

Figura 1 Equipo denominado Medulómetro utilizado para medir el diámetro y el tipo de medulación de muestras de fibras de diversas especies de animales.
La norma IWTO-8-04 fue utilizada para la evaluación del diámetro y medulación, para lo cual las fibras fueron cortadas en fragmentos de 0.8 mm utilizando el micrótomo de Hardy, que luego fueron dispersadas en una lámina portaobjeto utilizando una bagueta y aceite de cedro, y cubierta con una lámina cubreobjetos. Esta preparación fue colocada en el Medulómetro, iniciando las mediciones desde la esquina inferior izquierda para ir avanzando hacia arriba y hacia la derecha. El total de mediciones por muestra fue de 600.75 ± 7.48.
La caracterización de la medulación y diámetro de fibra en las cinco especies estuvo basada en estadística descriptiva (promedio, desviación y error estándar, valores mínimo y máximo). Para las comparaciones entre tasas de medulación se utilizó un modelo lineal general considerando a las especies como variables independientes y como variables dependientes al porcentaje de medulación y la media del diámetro de fibra. Para la evaluación del porcentaje de medulación al no tener una distribución normal se hizo una transformación logarítmica de la proporción de medulación a la que se sumó 1/6. Adicionalmente, para determinar la relación entre medulación y diámetro de fibra se hizo uso de la correlación de Pearson (r) y análisis de regresión simple, realizándose la evaluación de supuestos en base a los residuos (distribución normal, homogeneidad de varianza e independencia de errores) mediante estadísticos y diagramas. El software utilizado fue R v. 3.6.1 (R Core Team, 2019).
RESULTADOS
Medulación y Diámetro de fibra
No se encontraron muestras conteniendo médulas tipo kemp en lana de ovinos Merino, pero la tercera parte de ellas presentaban algún otro tipo de medulación. Sin embargo, en todas las muestras de fibras de las otras especies (llama, alpaca, conejo, paco-vicuña y conejo Angora) se encontraron fibras con algún tipo de medulación, a excepción de una sola muestra de fibra de llama que no contenía fibras meduladas. Asimismo, en más del 53% de las muestras de camélidos se observaron fibras fuertemente meduladas, tipo «kemp» o fuertemente meduladas. La menor frecuencia (33%) fue en fibras de conejo Angora (Cuadro 1).
Cuadro 1 Cantidad y porcentaje de muestras de lanas y fibras que presentan medulación y específicamente, medulación tipo kemp, en diferentes especies animales
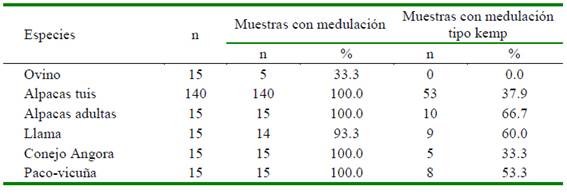
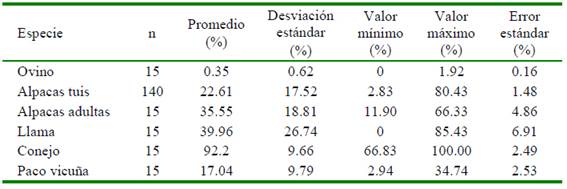
Las fibras de conejo Angora tuvieron la mayor incidencia de medulación, seguido de las fibras de alpacas y llamas, mientras que la menor incidencia se observó en lana de ovino Merino, siendo evidente el efecto de la especie animal sobre el porcentaje de medulación (Cuadro 2). Los menores rangos de variación de la incidencia de medulación se registró en lana de ovino Merino (1.92%), seguido de la fibra de paco-vicuña (31.8%) y de conejo Angora (33.17%); mientras que, los mayores rangos se observaron en fibras de llama (85.43%), seguido muy de cerca en fibras alpacas tuis machos de color blanco (76.4%).
Cuadro 2 Porcentaje de medulación y estadísticos relacionados de fibras de lana de ovino, alpacas, llamas, conejos y paco-vicuña
Los resultados de la incidencia de medulación y media del diámetro de fibra (MDF) por tipo de medulación se presentan en el Cuadro 3. Las fibras de paco-vicuña no solo son las más finas, sino que también tienen una baja tasa de medulación (17.04%). Asimismo, presentan los cuatro tipos de medulación (fragmentada, discontinua, continua y kemp). La fibra de conejo, aunque tuvo una alta tasa de medulación (92.2%) tuvo una media de diámetro bajo (14.72±0.67), indicando que las fibras finas también tienen médula en su estructura.
Cuadro 3 Porcentaje de medulación y media de diámetro de fibra (MDF) ± error estándar según tipo de medulación de lana y fibras de diferentes especies animales
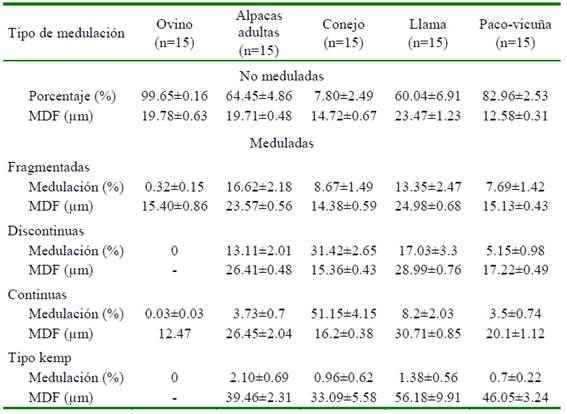
La lana de ovino y las fibras de alpaca presentaron medias de diámetro similares (alrededor de 19.7 µm), aunque la lana está conformada principalmente por fibras sin médula, mientras que las fibras de alpaca tienen una incidencia de medulación por encima del 35%. Por otro lado, las fibras de alpaca y llama tienen similares porcentajes de medulación; sin embargo, las fibras de alpaca son más finas que de la llama, pero ambas son más gruesas que las fibras de paco vicuña y de conejo Angora.
Es de notar un patrón característico, en torno a la media de diámetro y el porcentaje de medulación de las fibras, pues a excepción de la lana de ovino, el MDF incrementa en forma progresiva en casi todas las demás especies, desde las fibras con medulación fragmentada, pasando a la discontinua, continua y terminando con las fuertemente meduladas. Del mismo modo, las fibras no meduladas resultan ser más finas que las fibras con cualquier tipo de medulación (Cuadro 3, Figura 2).
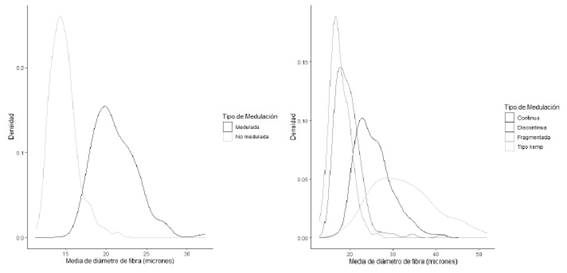
Figura 2 Diagramas de densidad del diámetro de fibras por tipo de medulación. Izquierda: Medias de diámetro de fibras meduladas y no meduladas. Derecha: Medias de diámetro de fibras por tipo de medulación
Los porcentajes de medulación de fibras de alpacas tuis machos y alpacas adultas fue- ron de 22.61±1.48 y 35.65±4.86%, respectivamente (p<0.05). Asimismo, el mayor porcentaje por tipo de medulación se observó en las fibras con medulación fragmentada, seguida de la medulación continua, discontinua y fuertemente medulada (9.17±0.66, 7.85±0.68, 4.86±0.43 y 0.72±0.14%, respectivamente); sin embargo, son más numero- sas las fibras sin medulación en comparación a las meduladas (Cuadro 4).
Relación entre Medulación y Diámetro de Fibra
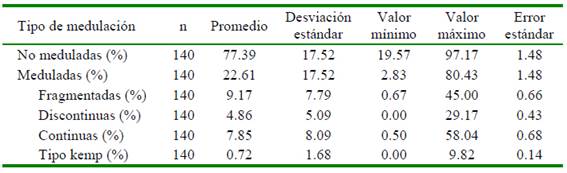
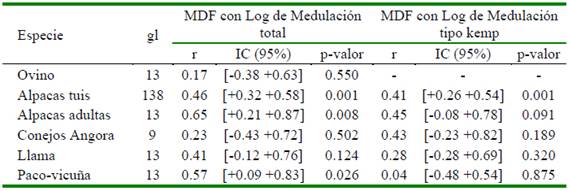
En los tipos de fibras analizados, se observa una aparente relación directa entre la incidencia de medulación con la MDF (evaluada mediante el coeficiente de correlación de Pearson: r). En el análisis estadístico en fibras de alpacas tuis, alpacas adultas y de paco-vicuña se encontró correlaciones positivas entre moderada y alta entre dichas variables (0.65, 0.46 y 0.57, respectivamente). También se encontró que en fibras de alpacas tuis existe una moderada correlación entre la incidencia de medulación tipo kemp con la MDF (Figura 3, Cuadro 5). Por otro lado, en lana de ovinos Merino se encontró correlaciones bajas no significativas, probablemente debido las bajas o nulas incidencias de medulación.
Cuadro 5 Coeficiente de correlación de Pearson (r), intervalo de confianza (IC) de r y p- valor, de la media del diámetro de fibra (MDF) con logaritmo de medulación total y medulación tipo kemp de fibras de cinco especies animales
gl: Grados de libertad
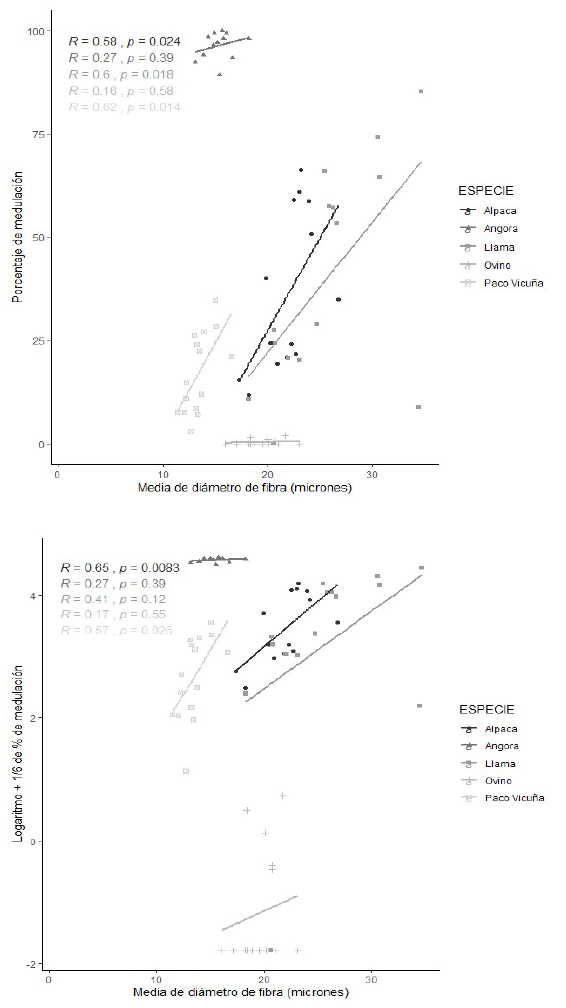
Figura 3 Diagramas de dispersión con ajuste de línea de regresión de la media del diámetro de fibra (MDF) y el porcentaje de medulación. Arriba: Sin transformación; Abajo: Con transformación logarítmica más 1/6) de fibras de alpaca, conejo Angora, llama, de lana de ovino y paco vicuña. Se incluye los coeficientes de correlación de Pearson
Los coeficientes de correlación en fibras de alpacas y paco-vicuñas indican que a mayor porcentaje de medulación corresponde mayor diámetro de las fibras. En el análisis de regresión, el coeficiente de regresión de la MDF sobre el logaritmo del porcentaje de medulación resulta significativo a nivel del 95% (bMDF,Log Med)= 1.11±0.2) solo en alpacas tuis, en tanto que en alpacas adultas resulta significativo a un nivel del 90%. Los coeficientes de regresión no fueron significativos para las fibras de las demás especies (Figura 3, Cuadro 6). En fibras de alpacas tuis también se encontró una relación directa entre los porcentajes transformados de los diferentes tipos de medulación (porcentaje de medulación total, fibras fragmentadas, discontinuas, continuas y tipo kemp) con la MDF (Cuadro 7), con coeficientes de regresión que varían entre 0.41 y 1.1 y con coeficientes de determinación moderados (r2 entre 0.10 y 0.19), que permite afirmar que a mayor incidencia de cualquier tipo de medulación corresponde una mayor MDF (Figura 3). Bajo estas consideraciones, con fines predictivos, se puede indicar que por cada unidad de incremento del porcentaje de medulación total transformado, la MDF incrementa en 1.11 ± 0.20 µm, siendo menor el incremento cuando se aumenta en una unidad los otros porcentajes de tipo de medulación transformados (Cuadro 7).
Cuadro 6 Estadísticos de regresión lineal simple de la media de diámetro de fibra (MDF, variable dependiente) sobre el porcentaje de medulación total transformado (variable independiente) en fibras de cinco especies animales
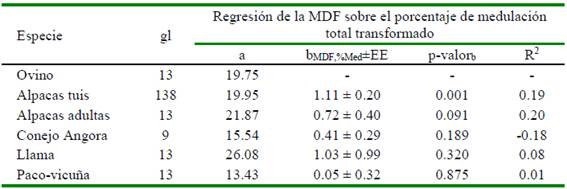
gl: Grados libertad; a: Intercepto; bMDF,%Med: Coeficiente de regresión; R2: Coeficiente de determinación
Cuadro 7 Estadísticos de regresión lineal simple de la media de diámetro de fibra (MDF) con el porcentaje transformado de medulación en fibras de alpacas Huacaya tuis de color blanco, según tipo de medulación
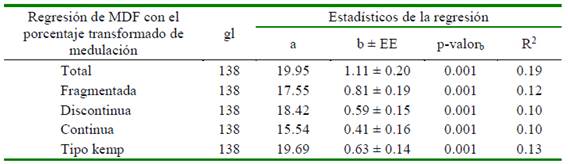
gl: Grados libertad; a: Intercepto; bMDF,%Med: Coeficiente de regresión; R2: Coeficiente de determinación
En una vista transversal, las fibras de lana son generalmente redondeadas, observándose ocasionalmente alguna medulación; sin embargo, las fibras de alpaca, llama y paco-vicuña varían de forma redonda a ovalada, observándose que las fibras más gruesas y con mayor diámetro de médula tienden a ser mayormente ovaladas. De otro lado, las fibras de conejo Angora son generalmente todas meduladas, pocas de forma redondeada, predominando médulas de forma lobulada, elíptica, bastante ovaladas y en algunas casi aplanadas. Estas se logran ver en fibras más gruesas, encontrándose entre dos y tres canales medulares, mientras que la presencia de medulas casi tubulares o levemente ovales se visualizan en las fibras finas y redondeadas. Las fibras de conejo vistas longitudinalmente tienen forma reticular o escalera, siendo esta de tipo uni o multiserial; es decir, apareciendo como si hubiera varias escaleras una al lado de otra (Figuras 4 y 5).
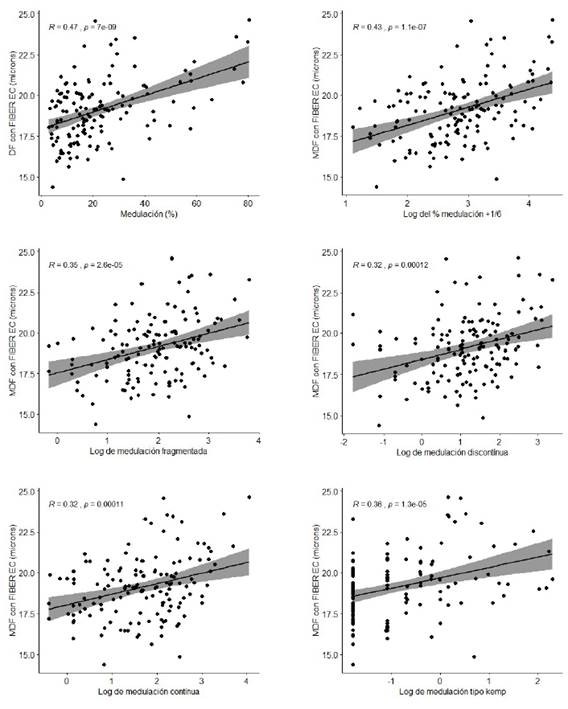
Figura 4 Diagramas de dispersión con ajuste de línea de regresión de la media de diámetro de fibra (MDF) y el logaritmo del porcentaje de medulación según tipos de medulación en fibras de alpacas tuis Huacaya de color blanco

DISCUSIÓN
La incidencia, porcentaje y forma de medulación resultan ser características propias de fibras de cada especie animal (McGregor, 2012). En estado silvestre, los animales cubiertos por fibras, tienen doble capa de fibras (Vásquez et al., 2000), unas son fibras finas («down») y otras muy gruesas («pelos o cerdas»), las que tienen alta incidencia de medulación (Antonini et al., 2004; Allain y Renieri, 2010); sin embargo, al pasar por el proceso de domesticación y debido a la aplicación de técnicas de mejoramiento animal en animales que producen lana y fibras, se optó por disminuir el diámetro de fibra, lo cual conllevó también a disminuir la incidencia de medulación. Estas explicaciones sustentarían que, en ovinos Merino, son pocos los animales cuyo vellón contiene vestigios de fibras meduladas. Los camélidos domésticos también fueron sometidos a procesos de mejoramiento genético mediante selección, habiéndose logrado disminuir el grosor en alpacas más no la incidencia de fibras meduladas, probablemente debido que las técnicas de selección empleadas no han sido del todo bien encaminadas y eficientes (Pinares et al., 2018). Contrariamente, las fibras de conejo Angora son generalmente meduladas, pero a la vez finas (Chattopadhyay et al., 2005), razón por la cual tienen una baja densidad y al confeccionar prendas en base a ella, resultan ser muy livianas (Süpüren et al., 2014).
La existencia de medulación en todas las muestras de camélidos (caso excepcional de una muestra de llama donde no se encontró medulación alguna), así como de medulación tipo «kemp» (en más del 33%), se debería a factores intrínsecos (genético, especie y variedad animal). Sin embargo, algunos factores extrínsecos pueden tener efecto sobre la medulación, pues se sabe que las alpacas, llamas y paco-vicuñas son criadas generalmente en condiciones extensivas y bajo sistemas de bajos recursos logísticos y de infraestructura (Quispe et al., 2009) y, por tanto, propensos a sufrir el efecto de la alimentación, manejo y sanidad, efectos que han sido discutidos y explicados por McGregor (2012) y Radzik et al. (2018); en tanto que efectos de edad, color y ubicación dentro de la zona corporal han sido comentados por Martínez et al. (1997).
El bajo porcentaje de medulación en las fibras de lana de ovinos Merino concuerda con los reportes de Ferguson et al. (2012) y McGregor (2012), quienes afirman que la medulación se presenta en forma esporádica. De otro lado, los porcentajes de medulación encontrados en fibras de alpacas adultas y alpacas tuis de color blanco son ligeramente mayores o similares, respectivamente, a los reportados por Lupton et al. (2006) y Cruz et al. (2019) y dentro del rango presentado por McGregor (2006). No obstante, otros autores reportan valores inferiores (Contreras, 2010; Ferguson et al., 2012; Córdova, 2015; Radzik et al., 2018; Pinares et al., 2019). Estas diferencias podrían deberse al método de medición empleado (Ferguson et al. [2012] utilizaron el OFDA 100), el lugar de crianza (Radzik et al., 2018; trabajaron con alpacas en Australia, África y Europa) y al rango de diámetro de fibras (Córdova, 2015 trabajó con fibras de 25-29 µm y en este estudio fue de 19.11±0.16 µm y 21.81±0.63 µm en alpacas tuis y adultas, respectivamente).
La mayor tasa de medulación de las fibras de camélidos, comparados con fibras de lana, generaría una menor densidad a las fibras de alpacas y llamas (Czaplicki, 2012). Asimismo, podría contribuir a explicar el efecto aislante, dado que la fibra de alpaca tiene mayor capacidad aislante que la lana, lo cual se debería a los espacios vacíos existentes dentro de la fibra (médula) que atraparían el aire del medioambiente (Wang et al., 2005). Por tanto, la medulación en fibras de camélidos más que un defecto podría ser una bondad (McGregor, 2012). Las fibras de medulación fragmentada, discontinua y continua no tienen un grosor exagerado (MDF menor a 26.5µm); sin embargo, las fibras que tienen medulación tipo «kemp», que son las llamadas fibras objetables tienen un serio defecto (ASTM, 1986; Lupton y Pfeiffer, 1998), pues producen el efecto picazón en alpacas (McGregor, 2006) y en llamas (Cochi, 1999; Frank et al., 2009).
Las fibras tipo kemp encontradas en alpacas adultas y llamas (MDF 39.5±2.3 y 56.2±9.9 µm, respectivamente) serían las que producen la sensación incómoda de picazón. Sin embargo, gran parte de este tipo de fibras pueden ser eliminados mediante el descerdado (Cochi, 1999; Wang et al., 2003; Frank et al., 2009; Poma, 2018). La selección animal sería otro proceso que permitiría disminuir la tasa de medulación tipo «kemp», pues esta característica tiene una moderada heredabilidad (Pinares et al., 2018), que permitiría tener un rápido avance genético.
El 92.2±2.5% de medulación encontrado en fibras de conejo Angora se ubica dentro del rango reportado (Herrmann et al., 1996; Blacutt, 2002; Onal et al., 2007; Rafat et al., 2007), aunque inferior al hallazgo de Chattopadhyay et al. (2005) en conejos Angora de la India, habiéndose demostrado que el origen y el lugar de crianza tienen efecto sobre la tasa de medulación (Herrmann et al., 1996).
En base a las correlaciones encontradas, la deducción de la relación directa entre porcentaje de medulación y la MDF concuerda con reportes en alpacas (Moore, 2015; McGregor, 2006). Asimismo, Radzik et al. (2018) encontraron que a medida que aumenta el grosor de las fibras disminuye el porcentaje de fibras no meduladas y de medulación fragmentada, pero incrementa la medulación continua. En llamas, Martínez et al. (1997) concluyeron que el porcentaje de medulación se incrementa con la edad y el mayor diámetro de fibra de llama, en tanto que Onal et al. (2007) encontraron correlaciones positivas mayores a 0.88 con la MDF.
La existencia de medulación en fibras finas es respaldada por Radzik et al. (2018), quienes, trabajando en alpacas, encontraron médulas en fibras con diámetros entre 14 y 20 µm. Similarmente, Radzik et al. (1995) encontraron fibras meduladas en lanas finas de ovinos nativos de la India, aunque McGregor (2006) indica que en fibras de alpacas con MDF de 18 µm a menos no se encontraría fibras meduladas.
Para fines de mejoramiento de la calidad de las fibras de alpacas y llamas (a través de la selección animal) resulta conveniente considerar la relación directa entre el diámetro (MDF) y el porcentaje de medulación (%M), pues al seleccionar alpacas de menor MDF indirectamente se obtendrían animales con menores porcentajes de medulación, o viceversa (Gupta et al., 1981, Cruz et al., 2019). A la fecha, el porcentaje de medulación no es considerado como criterio de selección, lo cual se debería a tres razones. Primero, a la fecha no existe un equipo que mida con buena precisión, exactitud y en forma práctica la medulación de la fibra de alpacas y llamas; segundo, porque no todas las fibras meduladas tienen un efecto negativo sobre la calidad de las fibras y por tanto no sería recomendable disminuir el porcentaje global de medulación; y tercero, porque solamente las fibras con medulación continua y especialmente las fibras tipo kemp o fibras objetables deberían ser eliminadas o drásticamente disminuidas a fin de mejorar la calidad y, por tanto, el precio de las fibras (Frank et al., 2009).
Así, aunque las alpacas producen vellones con fibras finas, (Quispe et al., 2009; Omachea et al., 2015; Vásquez et al., 2015, Pinares et al., 2018) -los cuales tendrían bajos porcentajes de medulación-, aún se tiene el problema del picazón, razón por la cual, algunos investigadores como Pinares et al. (2018) y Cruz et al. (2019), entre otros, vienen proponiendo considerar al porcentaje de fibras con medulación continua y tipo kemp como criterio de selección para mejorar la calidad de la fibra, pues a la fecha la disminución de la MDF pasaría ser relegada (Pinares et al., 2018).
Las vistas transversales de las fibras de lana Merino redondeadas estarían relacionadas con una MDF baja. Así, Frank et al. (2009) afirman que las fibras finas, al corte transversal, normalmente tienen una forma casi circular o apenas elíptica. McGregor (2012), asimismo, afirma que este patrón también se observa en fibras especiales. Los hallazgos del presente estudio en torno a las diferentes formas y tamaños de los canales medulares de las fibras de llamas y alpacas, concuerda con otros reportes que indican que la forma transversal de las médulas varía de acuerdo al rango de la MDF, de modo que las fibras que tienen médulas son redondeadas, mientras que en las fibras medianas y gruesas aparecen formas más irregulares (Frank et al., 2009; Allain y Renieri, 2010; Czaplicki, 2012).
Las vistas transversales de fibras de conejo, registradas en el presente trabajo, concuerdan con las reportadas por Chattopadhyay et al. (2005) y Allain y Renieri (2010), quienes encontraron fibras con uno a tres canales medulares. Asimismo, las vistas longitudinales observadas en este trabajo son similares a las medulaciones de tipo reticular encontrados por Langley y Kennedy (1981) y Onal et al. (2007), mientras que Süpüren et al. (2014) reportaron medulaciones tipo escalera uni y multiserial, que van de formas ovales a rectangulares.
Bajo estas consideraciones se puede indicar que en fibras de alpacas con mayor diámetro al corte transversal se encuentran fibras de forma de elíptica, con similares características en sus médulas. Esto no puede ampliarse a las fibras de conejos que, a pesar de ser bastante finas, las fibras en casi su totalidad son muy elípticas y con médulas lobuladas, principalmente en fibras de forma ovoide alargada. McGregor y Quispe (2018) encontraron que la elipticidad se incrementa en 0.006 por cada micrómetro en la MDF.
CONCLUSIONES
La tasa de medulación en ovinos Merinos es cercana a cero, en conejos Angora es cercana al 100% y en fibras de camélidos varía entre 17 y 40%, observándose menores tasas de medulación en paco-vicuña y alpacas tuis machos comparados con alpacas adultas.
Existe una relación directa entre la medulación con el diámetro de las fibras; sin embargo, la presencia de fibras meduladas se observa tanto en fibras gruesas como en fibras finas.
La forma de las fibras y las médulas varían según las especies; sin embargo, las fibras finas, tanto las fibras como las médulas son redondeadas, en tanto que en fibras gruesas ambas son bastante elípticas.

