











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
El Perú es considerado como uno de los cinco países con mayor diversidad de especies de primates no humanos (PNH) (Estrada et al., 2017). El país cuenta con 47 especies en 15 géneros y 3 familias (Pacheco et al., 2020; MINAGRI, 2020); sin embargo, 32% de estos animales presenta algún grado de amenaza debido a actividades antrópicas que modifican sus hábitats (MINAGRI, 2020), entre ellas la cacería indiscriminada (Aquino et al., 2014; Pérez-Peña et al., 2018). En Perú, la caza de subsistencia es una actividad legalmente aceptada mediante el Decreto Supremo Nº019-215-MINAGRI.
La dieta de los PNH está constituida esencialmente por frutos (Hawes y Peres, 2013), lo cual los convierte en diseminadores de semillas luego que pasan por su tracto digestivo; desempeñando de esta manera un papel primordial en la conservación de la vegetación tropical (Murcia, 1995; Domínguez- Domínguez y Morales-Mávil, 2006; Norden, 2014; Calle-Rendón et al., 2016). La alimentación de los PNH se complementa con flores, hojas tiernas y artrópodos (Bowler y Bodmer, 2011).
La investigación de la dieta de los PNH contribuye a mejorar la comprensión de su papel ecológico y de su coexistencia en los hábitats. La constitución de la dieta contribuye a conocer el uso de las fuentes de alimentación, en tanto que el análisis de la amplitud del nicho alimentario permite conocer su interacción con un determinado ambiente (Krebs, 1999). Del mismo modo, identificar la ecología alimentaria de los PNH es crucial para establecer estrategias para la conservación de las especies animales así como de los ecosistemas (Terborgh et al., 2008; Stevenson, 2011; Estrada et al., 2017; Andresen et al., 2018).
A pesar de los diversos estudios sobre los hábitos alimenticios de los PNH, son es- casos los estudios que se han realizado en la Amazonía peruana. Esta falta de información se debe a diversos factores ambientales tales como la accesibilidad a ciertas zonas del bosque y a la altura del dosel, que dificulta la visualización de los alimentos que son consumidos, así como la colecta de las heces (Izawa, 1975; Moreno-Black, 1978). Del mismo modo, la caza furtiva provoca que estas especies alteren su comportamiento natural (Izawa, 1975), impidiendo la observación directa que convencionalmente se utiliza para estudiar la ecología alimentaria (Altmann, 1974), de allí que se requieren métodos alter- nativos para obtener información respecto a su dieta.
El presente estudio planteó utilizar los despojos de los primates no humanos cazados para autoconsumo por los pobladores del norte de la Amazonía peruana como una estrategia para identificar la dieta de estas especies. Ante esto, se determinó y comparó la composición de la dieta de seis especies de primates, además de evaluar la amplitud y diversidad de las dietas. Esta metodología, poco empleada en trabajos de investigación, permitió obtener un tamaño de muestra considerable de estas especies de vida libre.
MATERIALES Y MÉTODOS
Área de Estudio
La investigación fue realizada en tres áreas del departamento de Loreto, Perú. Se trabajó en la comunidad José Olaya (02°33’43" S, 71°57’33’ E) del río Corrientes, en la comunidad Nueva Esperanza (04º19’53" S, 71º57’33" E) del río Yavarí-Mirín y en las comunidades del Porvenir y Los Jardines (2°49’04" S - 76°27’04" E) del río Pastaza. La zona se caracteriza por ser tropical, con temperatura media de 26 °C, precipitación pluvial anual de 2800-3200 mm y con periodo de lluvias entre noviembre y mayo (Egoavil, 1992; OGIE, 2019).
Colecta de Muestras
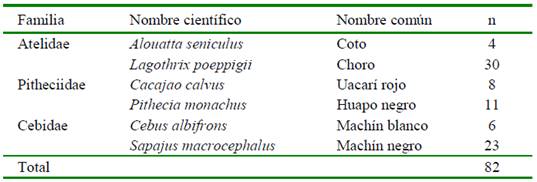
Los estómagos provinieron de 82 primates no humanos (Cuadro 1) cazados con fines de autoconsumo y donados por los cazadores. Las donaciones se realizaron entre años 2012 y 2015, tanto durante las épocas de seca como de lluvias. Los cazadores locales que colaboraron en el estudio (ocho en total), fueron capacitados para realizar la recolección de los órganos digestivos abdominales y torácicos que serían desechados, donando las vísceras conservadas en frascos herméticos con formol al 4% (v/v) proveídos por los investigadores. Los cazadores codificaron los frascos, incluyendo la especie y el sexo de cada individuo, así como el lugar, fecha de la caza.
Las especies de primates cazados pertenecen a tres familias de Platyrrhini (Cuadro 1). El tamaño de la muestra (N) refleja la preferencia de los cazadores, entre estos Lagothrix poeppigii y Sapajus macrocephalus. Las especies cazadas tienen hábitos de alimentación principalmente frugívoros, aunque difieren entre los elementos complementarios de su dieta. Alouatta seniculus tiene un mayor consumo de hojas y L. poeppigii consume hojas tiernas y artrópodos, mientras que los pitécidos son conocidos depredadores de semillas y los cébidos son omnívoros (Norconk et al., 2009).
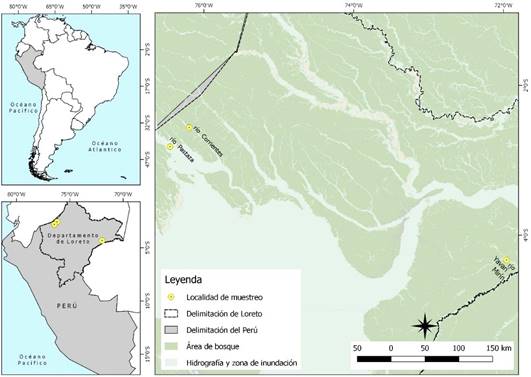
La recolección biológica de todas las muestras provino únicamente de los animales cazados por la población local, con autorización de la Dirección de Gestión Forestal y de Fauna Silvestre según Resolución Directoral 0350-2012-AG-DGFFS-DGEFFS, debido a que en el Perú la caza de subsistencia es considerada una actividad lícita, según Ley 2011 Nº 29763.
Contenido Estomacal
Para el análisis del contenido del estómago, se abrieron los estómagos longitudinalmente y se lavó su contenido con agua potable a presión suave mediante un filtro tamiz de 5 mm, pero se guardó el filtrado ya que contenía semillas de menor tamaño. Los contenidos gástricos se pusieron en recipientes individuales y fueron secados en horno a 60 ºC por 6-8 horas (Dubost y Henry, 2006), y se pesaron. Las semillas removidas en el filtro de 5 mm fueron aisladas, en tanto que el filtrado seco se pasó en un segundo tamiz de 0.2 mm. El aislamiento de las semillas de menor tamaño y de los artrópodos fue realizado utilizando un estereoscopio y la identificación fue realizada por entomólogos y botánicos de la región y por consulta a guías de campo (Loayza y Paine, 2014; Mozombite 2018; Vásquez-Arévalo et al., 2021). En la categorización taxonómica de artrópodos y semillas se determinó la clase, orden y familia. En el caso de las semillas, se determinó hasta el nivel del género y en pocos hasta especie.
Análisis Estadístico
La diversidad de fauna y flora se estimó en términos de la riqueza de artrópodos y de semillas en la dieta mediante el Índice de Diversidad de Simpson (λ), según la ecuación λ =1-(Pi)2. La diversidad es menor si los valores de λ se acercan a cero y es mayor si se acercan a uno (Kumar y Ram 2005, Sahu et al., 2008).
Se estimó la amplitud de los nichos tróficos faunívoro y frugívoro de los PNH a través del índice de Levin (B), con la ecuación B=1/(Pi)2. Este índice permitió medir comparativamente los hábitos alimenticios de las especies de primates. El valor de B puede variar de uno hasta n, donde n representa al total de recursos alimenticios que contienen los estómagos de una misma especie de PNH. Si el valor de B es cercano a uno su espectro de alimentación es limitado y se le considera como especialista, el aumento del valor de n indica un mayor espectro de alimentos, y considera que la especie presenta un hábito más generalista (Krebs,1999).
En ambas ecuaciones P representa a la proporción de cada morfotipo respecto al total de cada categoría de alimentos como semillas o artrópodos consumido por cada especie de PNH.
RESULTADOS
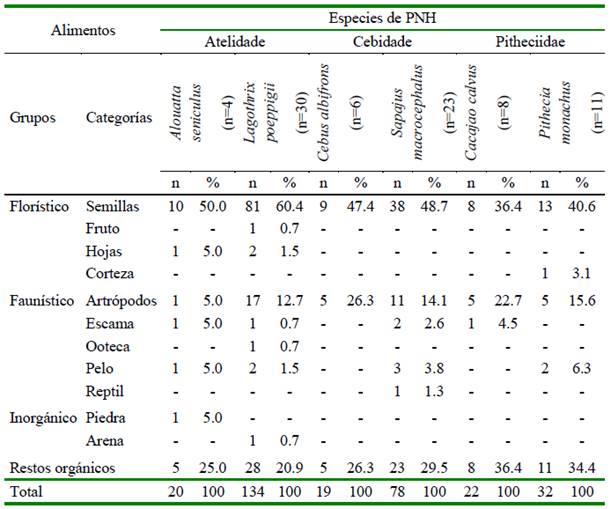
Se hallaron 11 categorías de alimentos en los contenidos gástricos, los que fueron agrupados como tipo florístico (semillas, hojas, corteza y fruto entero pequeño), faunístico (artrópodos, escama, pelo, reptil y ooteca) e inorgánico (arena y piedra) (Cuadro 2). Los componentes con diámetros <5 mm se consideraron como restos orgánicos, representando el 26.1% de los ítems registrados, pero no fue posible identificarlos dado a su avanzado proceso de digestión. La composición alimentaria más frecuente fueron los artrópodos y semillas (Cuadro 2). Las especies de primates con mayor frecuencia de semillas en sus estómagos fueron A. seniculus y L. poeppigii, y aquellos con mayor cantidad de artrópodos en la dieta fueron Cacajao calvus y Cebus albifrons (Cuadro 2).
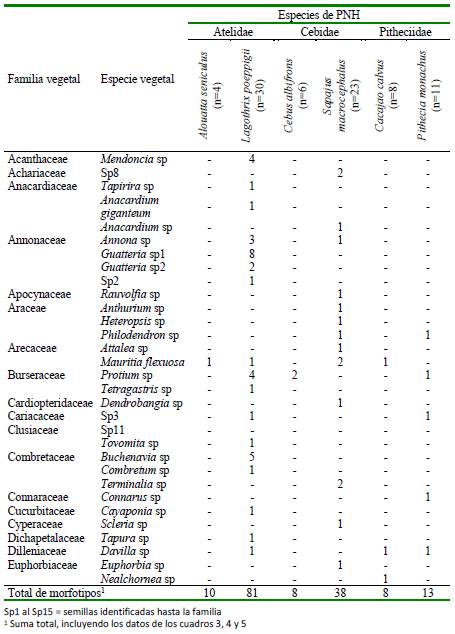
Se encontraron 133 tipos de semillas, pudiéndose identificar el 58% de ellas (n=77). Se halló mayor riqueza de semillas en Sapajus macrocephalus y L. poeppigii (Cuadro 3-5). Las semillas que fueron identificadas pertenecían a 35 familias (Cuadros 3-5). Las familias con mayor cantidad de especies fueron la Sapotaceae (n=7 especies), Moraceae (n=5), Solanacea (n=5), Fabaceae (n=5), Myrtaceae (n=4), Urticacea (n=4) y Annonaceae (n=4). De las especies de semillas, tres fueron consumidas por al menos cuatro especies de primates: Ficus sp (por cinco PNH), Cecropia sp y Mauritia flexuosa (por cuatro PNH) (Cuadros 3-5).
Cuadro 2 Presencia de categorías alimentarias en número absoluto (n) y frecuencia relativa (%) en los contenidos estomacales de las especies de primates no humanos cazados para consumo en el noreste de la Amazonía peruana (Loreto, 2012-2015)
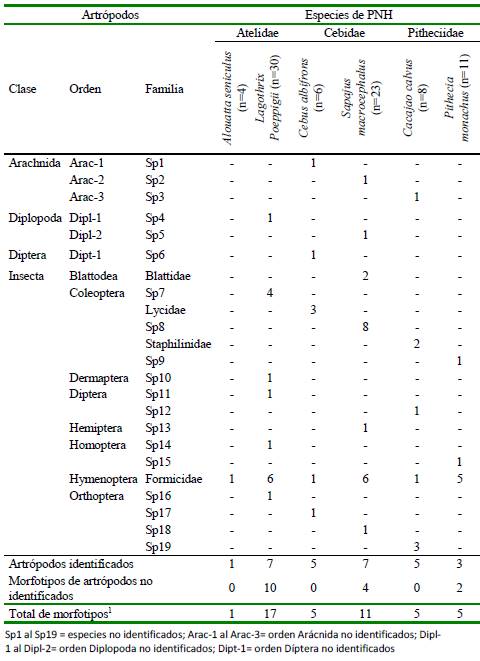
Se hallaron 39 tipos de artrópodos, identificándose 59% hasta al menos la categoría de clase (n=23). Los artrópodos identificados formaron parte de cuatro clases y ocho órdenes (Cuadro 6). La mayoría perteneció a la clase Insecta (74%, n=17 morfotipos), seguidos por Arachinida (13%, n=3), Diplopoda (9%, n=2) y Diptera (4%, n=1).
La mayor riqueza de insectos (Insecta) correspondió a las órdenes Coleóptera (n=5 morfotipos) y Orthoptera (n=4). L. poeppigii y S. macrocephalus fueron las especies de PNH con mayor riqueza de familias de artrópodos (Cuadro 6).
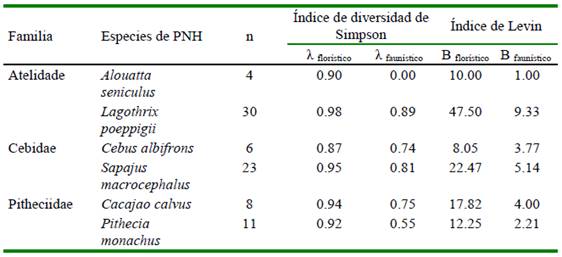
El Índice de Diversidad de Simpson (λ) de los componentes de frutos fue mayor que el correspondiente a fauna (Cuadro 7). La diversidad de frutos fue bastante similar entre especies de PHN aunque ligeramente menor para C. albifrons (0.87), en tanto que la amplitud del nicho frugívoro (B) fue menor en C. albifrons (8.05) y mayor en L. poeppigii (47.50). Por otro lado, λ varió entre 0.0 para A. seniculus y 0.89 para L. poeppigii, en tanto que el Bfaunístico varió entre 1.00 para A. seniculus) y 9.33 para L. poeppigii.
Cuadro 3 Frecuencia absoluta de semillas registradas en los contenidos estomacales de seis especies de primates no humanos (PNH) cazados para consumo en el noreste de la Amazonía peruana (Loreto, 2012-2015). Parte I
Cuadro 4 Frecuencia absoluta de semillas registradas en los contenidos estomacales de seis especies de primates no humanos (PNH) cazados para consumo en el noreste de la Amazonía peruana (Loreto, 2012-2015). Parte II
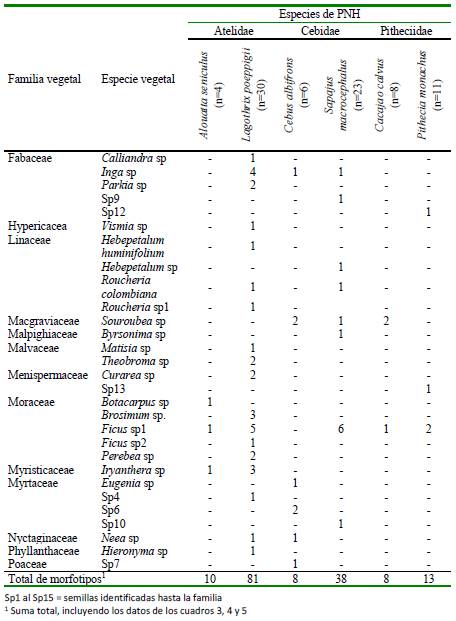
Cuadro 5 Frecuencia absoluta de semillas registradas en los contenidos estomacales de seis especies de primates no humanos (PNH) cazados para consumo en el noreste de la Amazonía peruana (Loreto, 2012-2015). Parte III
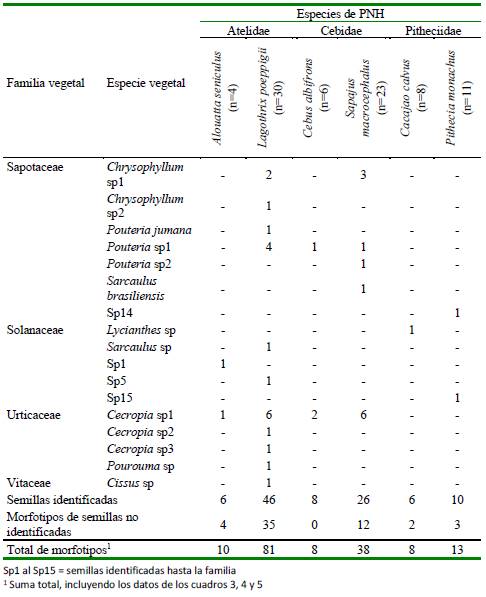
Cuadro 6 Frecuencia absoluta de artrópodos registrada en los contenidos estomacales de seis especies de primates no humanos (PNH) cazados para consumo en el noreste de la Amazonía peruana (Loreto, 2012-2015)
Los resultados indican que Lagothrix poeppigii es la especie con mayor diversidad y amplitud de uso de frutos (λ =0.98; B=47.50) y de artrópodos (λ=0.89; B=9.33) en su dieta, seguido por S. macrocephalus en la dieta a base de frutos y por C. calvus con el consumo de artrópodos (Cuadro 7).
DISCUSIÓN
Los estudios sobre la ecología alimentaria tienen gran relevancia debido a que contribuyen a conocer como los primates no humanos utilizan los recursos alimenticios del ecosistema. La investigación demuestra que las semillas constituyen el principal componente en las muestras de contenido gástrico de los PNH. La cantidad de artrópodos hallados en la dieta también confirma la importancia de la fauna como proveedor de proteínas para las especies en estudio. Los resultados coinciden con Aldana (2014), así como con Hawes y Peres (2013). La información sobre los frutos y artrópodos que constituyen la dieta de los PNH amazónicos es importante para establecer estrategias para la conservación de su hábitat y subsistencia en cautiverio (Williams et al., 2015).
La comparación de la diversidad y la amplitud trófica entre PNH simpátricos contribuye a describir diversas estrategias de supervivencia en un mismo hábitat. Las dietas de S. macrocephalus y L. poeppigii sugieren que son especies generalistas, en tanto que las dietas de Cacajao calvus, Cebus albifrons y Pithecia monachus sugieren que son especies con dietas especializadas. Las especies generalistas en periodos de escasez podrían sobrevivir sin mayores dificultades consumiendo diversos recursos de su entorno, en tanto que en caso de agotarse el recurso preferido de las especies especialistas, estarían obligados a un mayor gasto de energía en la búsqueda de dicho alimento, lo cual podría comprometer su supervivencia (Marshall et al., 2009). Sin embargo, se debe tener cierta cautela con la interpretación de los resultados, debido a la elevada proporción de alimentos que no han podido ser identificados debido a su avanzada digestión (Norconk, 2020).
La mayoría de los primates estudiados consumieron los frutos de Cecropia sp1, Ficus sp1 y Mauritia flexuosa. Al parecer, Cecropia sp y Ficus sp son recursos indispensables en la dieta de los PNH neotropicales, los cuales favorecen la supervivencia de estos animales en periodos críticos de oferta alimentaria (Felton et al., 2008; Luna, 2014). Las especies de estos géneros presentan una fenología asincrónica (Guariguata y Kattan, 2002; Tweheyo y Lye, 2003), debido a que están disponibles durante todo el año, de modo que pueden ser consumidos cuando se reduce la disponibilidad de otras especies de frutos.
La variedad de la ingesta de frutos está directamente relacionada con la multiplicidad de frutos disponibles (Stevenson, 2011). Por lo tanto, su consumo está asociado con el mantenimiento de la heterogeneidad de los bosques tropicales (Terborgh et al., 2008; Stevenson, 2011), pues estos PNH, especialmente Lagothrix sp, son dispersores de semillas, contribuyendo al mantenimiento y reforestación de los bosques (Andresen et al., 2018).
A pesar de que los artrópodos son un componente importante y reconocido de la dieta de algunas especies de primates neotropicales (Stevenson, 2011; Aldana, 2014), se dispone de pocos estudios que demuestren la composición de la fauna ingerida. La diversidad de artrópodos y la amplitud del nicho encontradas en esta investigación confirman la importancia de los artrópodos como fuente de proteínas para los PNH, especialmente para S. macrocephalus, L. poeppigii y C. calvus.
Cuadro 7 Índice de Diversidad de Simpson e Índice de Levin (amplitud del nicho) de los morfotipos de frutos y artrópodos hallados en los contenidos estomacales de seis especies de primates no humanos (PNH) cazados para consumo en el noreste de la Amazonía peruana (Loreto, 2012-2015)
Se reporta que el consumo de artrópodos suele ser mayor en primates de pequeño tamaño (Hawes y Peres, 2013); sin embargo, la importancia de la ingesta de artrópodos parece general para la mayoría de las especias estudiadas, independientemente de su tamaño, incluyendo a C. albifrons y P. monachus, que son las especies más especialistas en lo referente a la dieta faunívora. A nivel nutricional, el aporte proteico y de lípidos de los insectos es mayor que aquel de las plantas y frutos en general (Rothman et al., 2014), siendo las proteínas de alta calidad (Gutiérrez, 2005).
CONCLUSIONES
La dieta de los PNH se caracteriza por una elevada diversidad de frutos y artrópodos.
Lagothrix poeppigii es la especie más generalista en el uso de frutos y artrópodos.
Cebus albifrons es la especie más especialista en el consumo de frutos.
Pithecia monachus es la especie más especialista en el consumo de artrópodos.
Frutos de los géneros Cecropia sp, Guateria sp, Buchenavia sp y Ficus sp son los más ingeridos por las poblaciones de PNH.
Los artrópodos de los órdenes Coleoptera y Hymenoptera son los recursos faunísticos más ingeridos por los PNH.
Agradecimientos
Nuestro agradecimiento a los pobladores locales de las cuatro comunidades que participaron voluntariamente en la recolección de las muestras biológicas. La autora Anamélia de Souza Jesus fue acreedora de una beca doctoral otorgada por la Fundación Amazon de Apoyo al Estudio e Investigación de Pará (FAPESPA) con registro N.° 007/2017-FAPESPA/UFRA.

